
羊栖菜硫酸多糖诱导下的人红白血病HEL细胞差异表达基因分析
2021-02-22 08:48丁浩淼杜昊霏付瑞洁金旭东陈彤妮汪财生钱国英
核农学报 2021年3期
丁浩淼 杜昊霏 付瑞洁 金旭东 谢 策 陈彤妮 汪财生 钱国英
(浙江万里学院生物与环境学院,浙江 宁波 315100)
白血病是一类造血干细胞恶性克隆性疾病。克隆性白血病因为增殖失控、分化障碍、凋亡受阻等机制在骨髓和其他造血组织中大量累积,并浸润其他非造血组织和器官,同时抑制正常造血功能[1-2]。据报道,我国各地区白血病的发病率在各种肿瘤中位居第六位。白血病主要包括急性淋巴细胞白血病、急性髓系白血病、慢性淋巴细胞白血病和慢性髓系白血病4 种类型[3-4]。当前白血病常规疗法包括化学疗法、放射疗法、靶向疗法和骨髓移植等。尽管过去几十年发达国家在治疗方面取得重大进展,但白血病患者的治疗效果仍存在显著差异[5-7]。化学疗法是一种常见的治疗方式,但其会带来严重的不良反应且效果较差[8]。且这些不良反应与致残性副作用相关,如贫血、免疫抑制和脱发等。因此,对患者来说需要低毒性的新型抗肿瘤药物。当前,已经研究了诸多来源于植物、真菌、海洋生物和微生物的天然活性成分,并发现它们具有抗癌特性。因此可从植物、动物和微生物中发掘对癌细胞具有高功效且对正常细胞无毒性的新天然产物。在生物活性天然产物领域,当前的趋势是将研究对象转向海洋生物[9]。最近,已发现源自海藻的化合物与多种健康促进作用有关,特别是抗氧化、抗炎、抗病毒、抗微生物和抗肿瘤作用[10-11]。多糖作为一种天然的抗肿瘤物质,受到越来越多的关注,多糖的研究工作正逐渐转向肿瘤疾病的预防和临床干预[12]。
羊栖菜(Sargassum fusiforme) 隶属褐藻门(Phaeophuta) 墨角藻目( Fucales) 马尾藻科(Sargassaceae),又名玉草、六角菜、鹿角尖,在我国辽东半岛、山东、浙江、福建、广东浅海域均有分布,日本和韩国也有生长。《神农本草》记载羊栖菜可用于治疗甲状腺疾病,其作为食品和药物已有数千年的历史[13-15]。羊栖菜中含有多种对身体有益的生物活性物质,如硫酸酯多糖、海藻酸钠和岩藻黄质等[16]。近年来发现天然来源的多糖具有多种生物活性且毒副作用小,已在生物和医学应用中引起广泛关注[17]。羊栖菜硫酸酯多糖是一种含有硫酸基团的天然多糖衍生物,具有多种生物功能,如抗高血脂[18]、抗氧化[19]、免疫增强[20]、降血糖[21]和抗肿瘤[22]。先前研究已经报道了羊栖菜的多糖在诱导白血病、胃癌、膀胱癌和乳腺癌细胞凋亡方面的潜力。然而,尚未阐明从羊栖菜中纯化的多糖抗肿瘤特性及其潜在细胞和分子机理。
转录组分析是对基因表达谱的全面认识,为基因组研究、大规模功能基因和分子标记识别提供了一种有效的手段。近年来,转录组分析已广泛应用于癌细胞研究中,如人肺癌细胞(NCI-H292)[23]、人结肠腺癌细胞(Caco-2)[24]、小鼠单核巨噬细胞白血病细胞(RAW 264.7)[25]、人肝癌细胞(HepG2)[26]、人非小细胞肺癌细胞(A549)[27]、 人肾透明细胞癌细胞(Caki-1)[28]和人乳腺癌细胞(MDA-MB-231)[29]。本研究对羊栖菜硫酸多糖( sargassum fusiforme polysaccharide Ⅱ, SFPS Ⅱ) 诱导的人红白血病(human erytro leukemia cells, HEL)细胞进行了转录组分析,以期筛选相关的应答基因并分析凋亡相关信号通路,有利于了解SFPS Ⅱ诱导HEL 细胞凋亡机制,并为利用羊栖菜硫酸多糖预防和治疗白血病提供一定的理论基础。
1 材料与方法
1.1 主要试剂
RPMI-1640 培养基、高糖DMEM 培养基、胎牛血清(fetal bovine serum,FBS)、抗生素P/S(青霉素10 000 U.mL-1,链霉素10 000 μg.mL-1)和胰酶,美国Gibco 公司;二甲基亚砜(dimethyl sulfoxide,DMSO)、四甲基偶氮唑蓝(methyl thiazolyl tetrazolium,MTT)和CCK-8,北京索莱宝公司;25 cm2培养瓶、96 孔板和6孔 板, 美国Corning 公 司; TRIzol、 PrimeScriptTMRT reagent Kit with gDNA Eraser 和SYBR Premix Ex TaqTMⅡ,日本TaKaRa 公司。
1.2 主要仪器与设备
Forma series Ⅱwater jacket 型恒温CO2培养箱和5417R 高速冷冻离心机,美国Thermo 公司;Heal Force BIOsafe 12 超净工作台,上海力申科学仪器有限公司;Eclipse Ti-S 荧光倒置显微镜,日本Nikon 公司;CFX96Touch 荧光定量PCR 系统,美国Bio-Rad 公司;SpectraMax190 连续波长多功能酶标仪,美国MD 公司;Agilent 2100 生物分析仪,美国Agilent 公司。
1.3 试验细胞
人红白血病细胞(HEL)和人胚肺成纤维细胞(human embryonic lung cells, MRC-5)购自中国科学院上海细胞库,HEL 细胞在含10% FBS 的RPMI-1640 培养基中,MRC-5 细胞在含10% FBS 的高糖DMEM 培养基中。
1.4 试验材料
前期试验[30-31]分离得到一种羊栖菜硫酸多糖(SFPS Ⅱ),多糖检测纯度达96%以上且不含蛋白质,SFPS Ⅱ红外光谱表征具有糖类特征结构,且含有硫酸基功能基团,在1 250 cm-1处的峰增强,具有硫酸根S=O 的对称伸缩振动,754.6 cm-1和842 cm-1处α-吡喃环的特征吸收。SFPS Ⅱ采用硫酸钡比浊法检测硫酸基含量为26.47%。
1.5 细胞培养
HEL 细胞株复苏后,在37℃、5%CO2培养箱,用含有10%胎牛血清、1%青霉素-链霉素的RPMI-1640培养基对HEL 细胞进行常规培养。MRC-5 细胞株复苏后,在37℃、5%CO2条件下,用含有10%胎牛血清、1%青霉素-链霉素的高糖DMEM 培养基对MRC-5 细胞进行常规培养。
1.6 CCK-8 检测HEL 细胞增殖抑制作用
取对数生长期的HEL 细胞,以5×104个.mL-1接种于96 孔板中,每孔加入100 μL 细胞悬液,于细胞培养箱中继续培养24 h 后给药处理,每孔加药10 μL,每个浓度平行5 个复孔。阴性对照组加入等量无血清无抗生素的RPMI-1640 培养基,试验组分别加入SFPSⅡ终浓度为10、20、40、60、80、100 μg.mL-1,药物作用24 h 后,每孔中加入10 μL CCK-8 溶液,继续于培养箱中孵育4 h,用酶标仪测定450 nm 波长处OD值[32],按照公式计算细胞存活率:
As we are interested to the CRLB of the 2-D central DOAs for the ID sources,we can use the inversion of block matrices to obtain the following expression of CRLB(l):
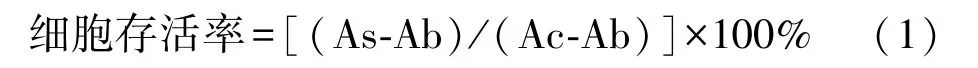
式中,As: 试验孔(含有细胞的培养基、CCK-8、SFPS Ⅱ);Ac:对照孔(含有细胞的培养基、CCK-8、不含SFPS Ⅱ);Ab:空白孔(不含有细胞和SFPS Ⅱ、含有CCK-8)。
1.7 MTT 检测MRC-5 细胞增殖抑制作用
取对数生长期的MRC-5 细胞,以5×104个.mL-1接种于96 孔板中,每孔加入细胞悬液100 μL,于细胞培养箱中继续培养24 h 后给药处理,每孔加药10 μL,每个浓度平行5 个复孔。阴性对照组加入等量无血清无抗生素的高糖DMEM 培养基,试验组分别加入SFPS Ⅱ终浓度为10、20、40、60、80、100 μg.mL-1,药物作用24 h 后,每孔中加入10 μL MTT(5 mg.mL-1)溶液,继续培养箱中孵育4 h,吸取上清液后加入100 μL无水DMSO,置于摇床上低速避光震荡15 min,使晶体充分溶解,用酶标仪测定570 nm 波长处OD 值,计算细胞存活率:

式中,As: 试验孔(含有细胞的培养基、MTT、SFPSⅡ);Ac:对照孔(含有细胞的培养基、MTT、不含SFPSⅡ);Ab:空白孔(不含有细胞和SFPS Ⅱ、含有MTT)。
1.8 RNA 提取及质量检测
收集SFPS Ⅱ处理和未处理的HEL 细胞用TRIzol试剂按照说明书提取细胞总RNA[33]。1%琼脂糖凝胶电泳和酶标仪鉴定RNA 的纯度、浓度及完整性,合格的总RNA 保存于-80℃冰箱待用。
1.9 RNA 文库构建及测序
RNA-Seq 试验由北京诺禾致源生物信息科技有限公司进行。每个样本中收集1 μg 总RNA(total RNA)用于RNA-Seq 文库的制备和测序。建库中使用的建库试剂盒为Illumina 的NEBNext® UltraTM RNA Library Prep Kit,对mRNA 进行纯化和片段化,制备cDNA 文库。文库库检合格后,将不同文库按照有效浓度及目标下机数据量的需求集中后进行Illumina 测序。
1.10 RNA-Seq 数据分析
测序片段经CASAVA 碱基识别转化为序列数据(reads),文件为fastq 格式。对原始数据进行过滤。主要包括去除带接头(adapter)的reads、去除含N(N表示无法确定碱基信息)的reads、去除低质reads(Qphred ≤20 的碱基数占整个read 长度的50%以上的reads)。同时,对干净数据(clean data)进行Q20、Q30 和GC 含量计算。后续所有分析均是基于clean data 进行的高质量分析。
使用HISAT2 v2.0.5 构建参考基因组的索引,并使用HISAT2 v2.0.5 将配对末端clean reads 与参照基因组比对。根据基因的长度计算每个基因的FPKM,并计算映射到该基因的读数(FPKM 指每百万碱基对测序的转录本序列片段的每千碱基片段的预期数量)。
1.11 差异表达基因及功能富集分析
使用DESeq2 R 软件(1.16.1)进行两个比较组合间的差异表达分析(每个组3 个生物学重复)。使用Benjamini 和Hochberg 的方法来调整所得P值以控制错误发现率。通过DESeq2 发现调整的P值<0.05 以及|log2fold change |≥2 作为显著差异表达的基因。通过ClusterProfiler R 软件实现差异表达基因的GO 富集分析和KEGG 富集通路分析。
1.12 RT-qPCR 验证分析

表1 Real-time PCR 引物序列Table 1 Primer sequences used for real-time PCR
1.13 数据统计与分析
采用SPSS 19.0 软件对试验数据进行统计分析,数据以±s表示,多组间比较数据符合正态分布且方差齐时用ANOVA 分析。
2 结果与分析
2.1 SFPS Ⅱ对HEL 和MRC-5 细胞增殖的影响
肿瘤细胞增殖是肿瘤发生和发展的最重要机制之一。由图1 可知,与对照组相比,SFPS Ⅱ处理HEL 细胞24 h,SFPS Ⅱ浓度为10 μg.mL-1时,HEL 细胞增殖活性虽有降低,但无统计学差异(P>0.05);SFPS Ⅱ浓度为20 ~100 μg.mL-1时,可不同程度抑制HEL 细胞增殖活性,且随着SFPS Ⅱ浓度的增加,其抑制细胞活性的作用逐渐增强,呈剂量依赖性。此外,SFPS Ⅱ处理HEL 细胞24 h 所对应的半数最大抑制浓度(IC50)为72.78±2.01 μg.mL-1。MTT 法检测发现,不同剂量的SFPS Ⅱ处理后,MRC-5 细胞活力不变(P>0.05),表明SFPS Ⅱ对正常细胞MRC-5 几乎无细胞毒性。因此,取24 h 的IC50为最大用药终浓度,即72 μg.mL-1SFPS Ⅱ作为后续机制研究最大用药终浓度。
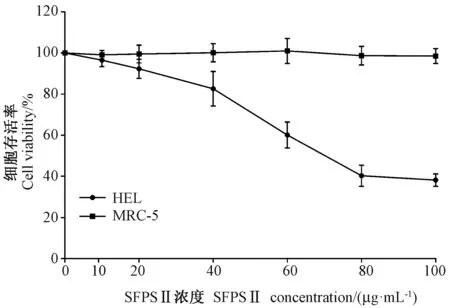
图1 SFPS Ⅱ对HEL 和MRC-5 细胞存活率的影响(±s, n=5)Fig. 1 Inhibitory effects of Sargassum fusiforme polysaccharide Ⅱ(SFPS Ⅱ) on the growth of human erythroleukemia (HEL) cells and human embryonic lung (MRC-5) cells(±s, n=5)
2.2 转录组测序数据评估及筛选差异表达基因
HEL 细胞经SFPS Ⅱ处理后,经Agilent 2100 精确检测RNA 完整后进行文库构建。细胞对照组和SFPSⅡ处理组分别得到57 383 361.33 和55 107 124.67 个原始数据(raw data),其中干净序列(clean data)分别为56 065 294.67 和53 623 604.67 个,分别占原始数据的97.70%和97.31%(表2)。测序数据的质量评估表明,每个样品中去除低质量数据(phred)数值大于30 的碱基占总碱基的百分比(Q30)均高于92.80%,且每个测序周期中的GC 和AT 含量几乎相等(表2)。HISAT2 比对软件由约翰霍普金斯大学开发,该软件采用全局和局部搜索的方法,利用大量FM 索引,覆盖整个基因组,能够将RNA-Seq 测得的reads 与基因组进行快速精确的比对,同时支持基因组特别大的物种。过滤后的数据使用HISAT2 软件成功地将96.37%~96.73%平均长度为150 bp 的干净序列与人类基因组成功匹配(Mapped reads)。SFPS Ⅱ处理组和对照组中有93.74%~93.92%的数据是比对到人类基因组唯一位置的(Unique mapped reads)。以上结果证明测序质量相当好,因此,随后的转录组分析是可靠的。本试验使用Cuffdiff 定量分析RPKM 的基因表达水平,每个基因的表达信号都被归一化,符合标准的正态分布,并在图2 中显示SFPS Ⅱ处理组和对照组基因表达水平之间的差异,86.4%SFPS Ⅱ处理组和87.5%对照组的唯一表达基因在1 ~32 个拷贝数之间且没有高于210个拷贝数的基因表达。经过严格筛选(P<0.05 且|log2fold change |≥2),共鉴定了1 248 个差异表达基因,红色代表显著上调基因,绿色代表显著下调基因,蓝色代表无显著变化基因,其中17.55%(219)基因上调,82.45%(1 029)基因下调。这1 248 个表达差异基因由火山图显示(图3)。

图2 SFPS Ⅱ处理组和对照组中log2 基因表达分布图Fig.2 Graph distribution of log 2-transformed gene expression in the SFPS Ⅱchallenge groups (SFPSⅡ)and the control (control)
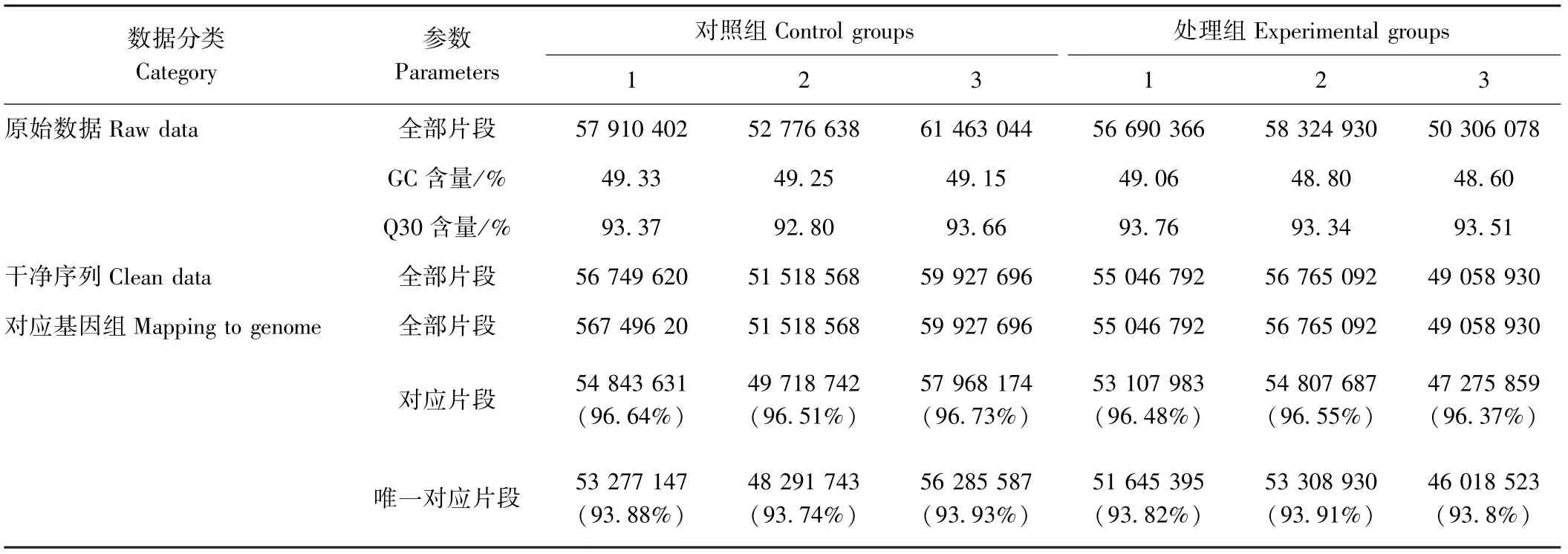
表2 转录组测序数据统计分析Table 2 Statistical analysis of transcriptome sequencing data
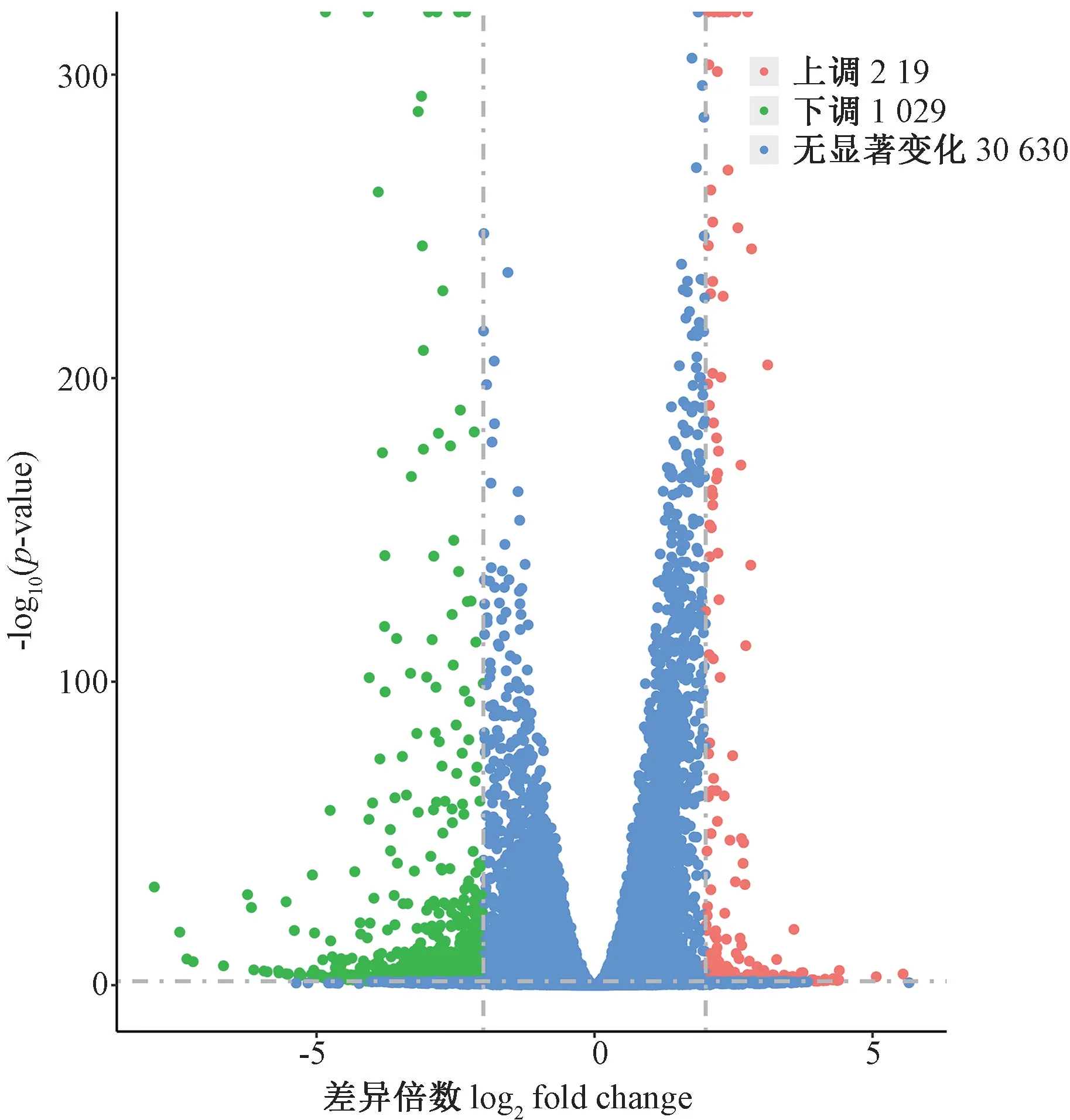
图3 SFPS Ⅱ处理后差异表达基因的火山图Fig.3 DEGs identified after SFPS Ⅱexposure
2.3 GO 功能显著性富集分析
对差异表达基因进行GO 功能分析,共得到464条GO 注释,其中75%(348 条)GO 注释为生物学过程,6.68%(31 条)GO 注释为细胞组分,18.32%(85条)GO 注释为分子功能(S2)(图4)。差异表达基因的生物学功能变化最显著的分子功能集中在DNA 构象变化和对未折叠蛋白的反应;细胞组分中,变化最显著的集中在蛋白-DNA 复合物、核小体和DNA 包装复合体;分子功能中,变化最显著的集中在养载体活性和二羧酸跨膜转运蛋白活性。生物学过程有个别显著丰富的GO 注释与凋亡和硫酸盐转运有关,如:膜电位调节(GO:0042391)、药物运输(GO:0015893)、黏膜固有免疫反应(GO:0002227)、硫酸盐跨膜转运(GO:1902358)、 细胞对肿瘤坏死因子的反应(GO:0071356), 以及外在凋亡信号通路调控(GO:2001236)。这些结果表明SFPS Ⅱ可能通过改变核酸合成、转录及翻译来调节HEL 细胞的生物学过程,激活细胞凋亡以及与细胞存活生长密切相关的免疫系统和代谢过程。
2.4 Pathway 显著性富集通路分析
为了进一步研究差异表达基因的功能,进行了KEGG 富集分析。结果显示(图5),人红白血病HEL细胞响应SFPS Ⅱ处理的1 248 个差异基因可以映射到256 条不同的生物途径,显著富集的途径(P<0.05)有16 条。差异表达基因的信号通路富集主要集中在系统性红斑狼疮、神经活性配体-受体相互作用,病毒致癌、cAMP 信号通路以及Rap 1 信号通路。
2.5 RT-qPCR 验证
为了证实RNA-Seq 数据的可靠性,随机选择了涉及不同功能的16 个差异基因进行RT-qPCR 分析。结果显示,RNA-Seq 和RT-qPCR 的数据之间具有高度相关性(R2=0.81),并且通过RT-qPCR 确定的所选差异基因的表达与RNA-Seq 的表达基本相同(图6~8),说明通过RNA-Seq 确定的DEGs 的表达谱是可靠和准确的。
3 讨论
癌症是一个重要的公共健康难题,肿瘤患者使用化疗药物后显著的复发比例说明肿瘤有耐药性,甚至患者在接触具有不同结构和作用机制的多种抗肿瘤药物后导致肿瘤多药耐药性[34-35],因此,化疗出现的耐药性是治疗各种癌症的主要障碍,治疗癌症亟需新一代药物。
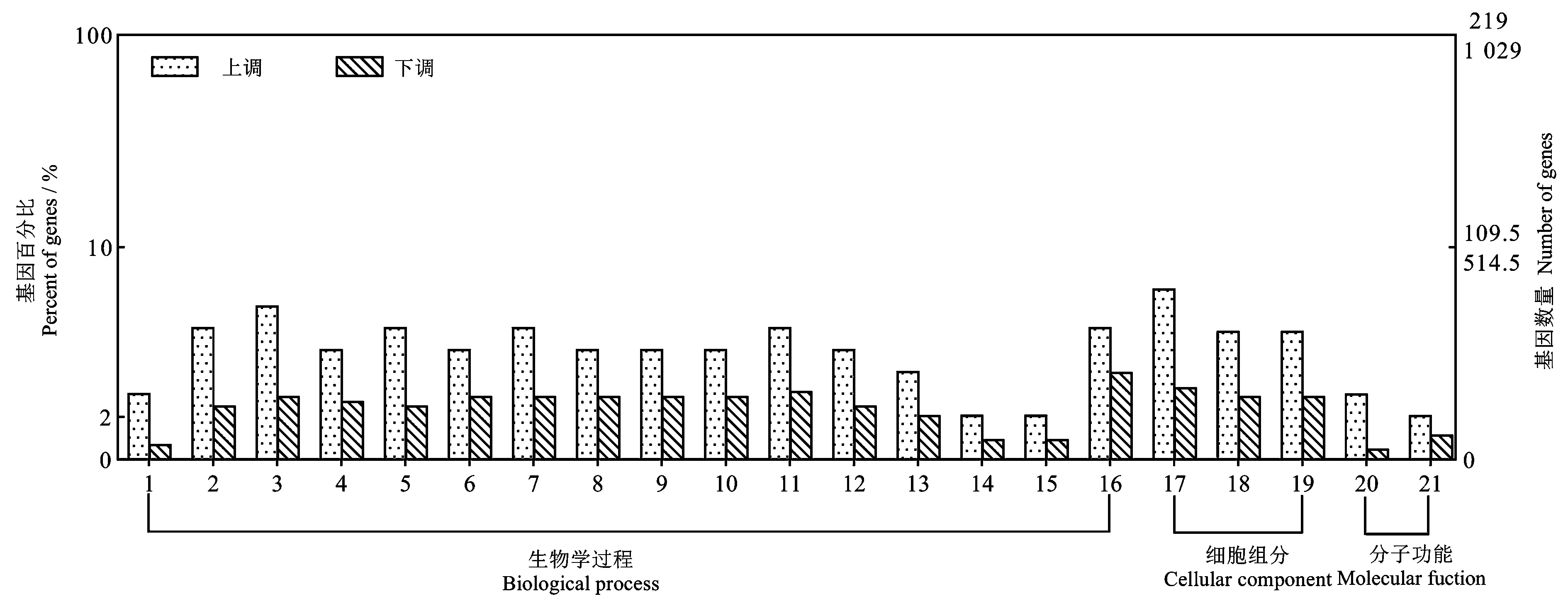
图4 SFPS Ⅱ处理组和对照组差异表达基因的GO 功能显著富集分析Fig.4 GO analysis of DEGs in the SFPS Ⅱversus the control
本研究从转录组概况分析SFPS Ⅱ抗肿瘤机制,研究了在红白血病细胞株HEL 中加入SFPS Ⅱ后的细胞增殖能力。结果表明,SFPS Ⅱ能降低HEL 红细胞对白血病细胞的细胞活力,说明SFPS Ⅱ可以抑制细胞增殖能力,降低细胞活力,且这些作用呈浓度依赖性,但对正常细胞株MRC-5 细胞无毒性作用。目前系统研究天然产物促进人红细胞白血病细胞凋亡作用的报道较少。Cheng 等[36]利用流式细胞术检测了黄芪提取物诱导的HEL 细胞凋亡,发现4.5 mg.mL-1黄芪提取物作用HEL 细胞3 ~5 d 后,能诱导HEL 细胞凋亡,且凋亡细胞的最大百分比(54.79%)出现在浓度为4.5 mg.mL-1,作用时间为5 d时,这表明黄芪提取物以浓度和暴露时间依赖性的方式诱导HEL 细胞凋亡,这与本研究结果一致。Fan 等[22]在不同的时间段(24、48 和72 h)研究不同浓度羊栖菜硫酸多糖(0、125、250、500、1 000 和2 000 μg.mL-1)对HepG2 细胞的细胞毒性作用,发现羊栖菜硫酸多糖对HepG2 细胞的生长有明显的剂量和时间依赖性。羊栖菜硫酸多糖作用HepG2 细胞48 h的IC50为1 158.6 μg.mL-1,是本研究IC50的近15 倍,这意味着纯化多糖SFPS Ⅱ具有更有效的抗肿瘤作用。上述研究只报道了天然产物对肿瘤细胞的抑制作用,对正常细胞是否有毒性尚未研究。本研究不仅设计了羊栖菜多糖对肿瘤细胞的试验,而且分析了羊栖菜多糖对正常细胞的影响,结果表明羊栖菜多糖对肿瘤细胞具有选择性,为了解SFPSⅡ诱导的抗肿瘤的分子机制,本研究首先分析了转录组差异表达基因,以鉴定表达受SFPS Ⅱ调控的基因。结果表明在SFPS Ⅱ作用下,存在大量差异表达基因。其中,219 个基因上调,1 029 个基因下调。此外,使用GO 和KEGG 数据库对相关途径的分析揭示了各种生物学过程和途径。
众所周知,细胞凋亡是细胞程序性死亡的过程,是引起癌细胞生长抑制的主要原因之一。Rap 1 是一种小GTP 酶,可以控制多种生物功能,如细胞粘附、细胞间连接形成和细胞急性。Rap 1 通过调节各种类型细胞中的整合素和其他粘附分子的功能,在细胞-细胞和细胞-基质中起主要作用。Rap 1 还以高度依赖于细胞类型的方式调节MAP 激酶(mitogen-activated protein kinase, MAPK)的活性。刘云昊等[37]收集100例术前未进行放化疗,且经病理证实为癌组织的临床标本,通过免疫组化技术检测RAP 1B 在胃癌组织中的表达情况,结果发现胃癌组织中Rap 1B 表达率高达72%。邹寒冰等[38]用白花蛇舌草处理人肾细胞腺癌ACHN 和人肾近曲小管HK-2 细胞24 h 后发现,ACHN 细胞增殖受到抑制,Rap 1GAP、Rap GEF3 和p-JNK 蛋白表达水平均显著下调,表明白花蛇舌草可能通过抑制Rap 1-JNK 信号通路来选择性抑制肾癌细胞ACHN 的增殖。SFPS Ⅱ作用HEL 细胞后信号通路显著改变的包括Rap 1 信号通路,说明羊栖菜多糖可能也是通过Rap 1 信号通路来选择性抑制HEL 细胞增殖,这需要通过特异性抑制剂阻断Rap 1 信号通路来进一步验证。本研究通过细胞抗肿瘤活性检测和转录组差异基因表达分析,为SFPS Ⅱ对HEL 细胞发挥作用提供了新见解。高度受控的程序性细胞死亡已成为治疗癌症的关键点。尽管多糖是抗肿瘤的一个新研究领域,且其理论和临床应用尚待进一步深入研究,但毫无疑问,多糖是天然药物研发的重要组成部分。因此,筛选可单独使用或与其他化学治疗药物组合使用的海藻多糖以提高治疗效果并减少癌症治疗中的副作用是一项重要的任务。

图5 SFPS Ⅱ处理组和空白组差异表达基因的KEGG 信号通路分析Fig.5 KEGG pathway analysis of DEGs in the SFPS Ⅱversus the control
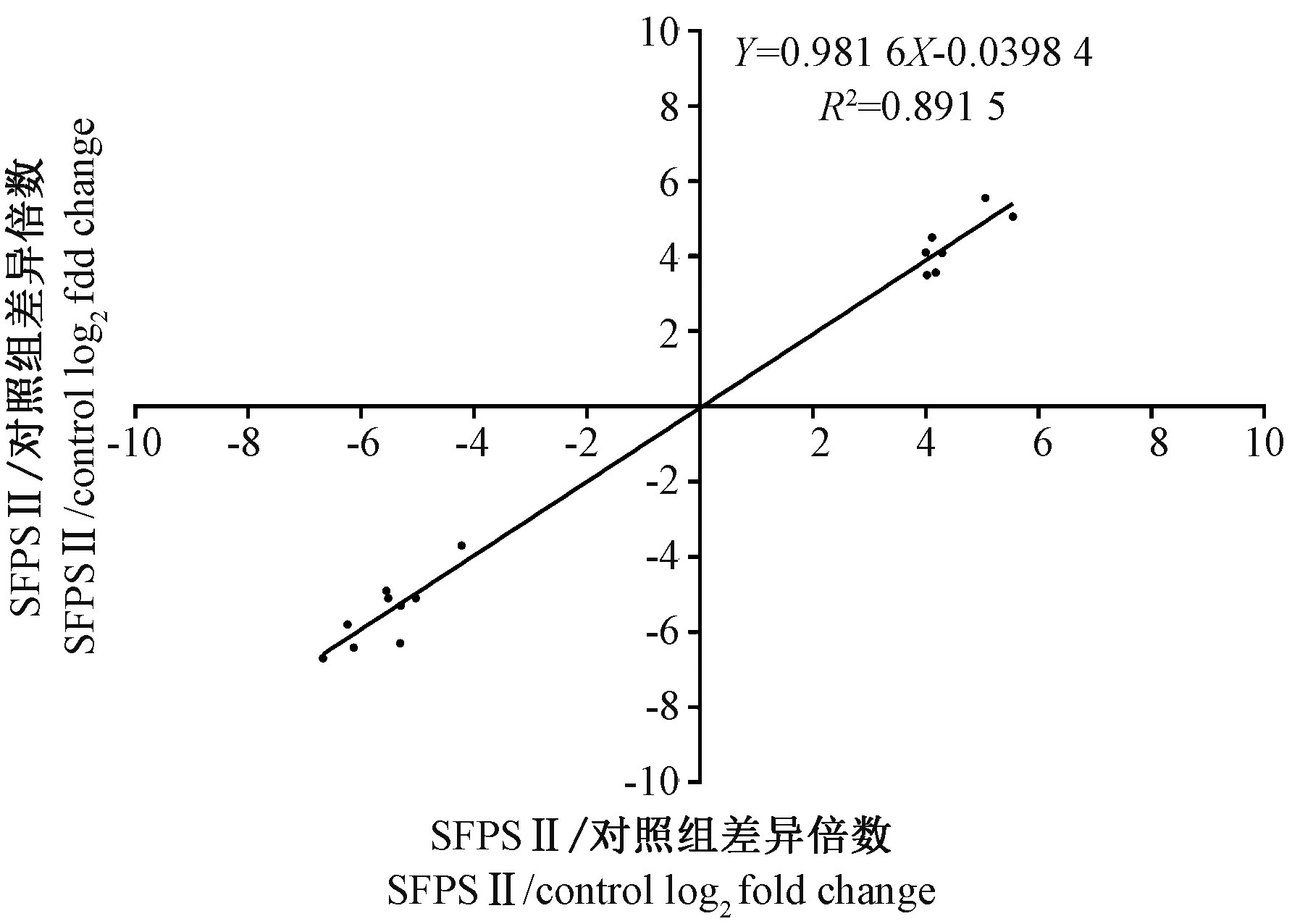
图7 RNA-Seq 和RT-qPCR 检测的基因相关表达率Fig.7 Correlation of the expression ratio of selected genes measured by RNA-Seq and RT-qPCR
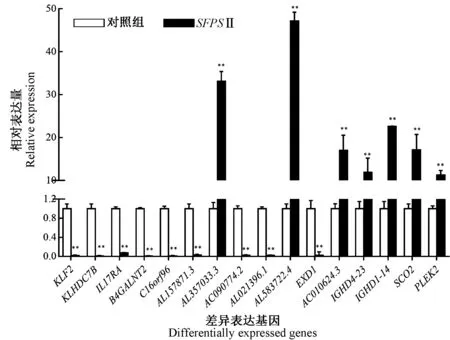
图8 RT-qPCR 检测SFPS Ⅱ处理组和对照组差异基因表达(±s, n=9)Fig.8 The relative expression of the selected DEGs by RT-qPCR between SFPS Ⅱexposure group and control(±s, n=9)
4 结论
本研究发现,羊栖菜硫酸多糖SFPS Ⅱ呈浓度依赖性地抑制HEL 细胞的体外增殖,表明SFPS Ⅱ具有抗肿瘤作用,可作为潜在的抗肿瘤药物;转录组分析为SFPS Ⅱ对HEL 细胞的作用提供了数据资源,这为进一步研究SFPS Ⅱ抗HEL 肿瘤细胞活性机制提供了靶点及理论依据,可指导抗癌药物的研发,从而更有效地预防和治疗癌症。
猜你喜欢
人人健康(2022年2期)2022-11-21
中国生物制品学杂志(2022年10期)2022-10-25
中国人兽共患病学报(2022年9期)2022-10-19
中老年保健(2021年9期)2021-08-24
科学导报(2021年29期)2021-06-03
父母必读(2019年4期)2019-09-09
科海故事博览·下旬刊(2019年6期)2019-04-16
科学中国人(2016年17期)2016-01-28
高中生学习·高三版(2014年3期)2014-04-29
食品工业科技(2014年23期)2014-03-11
