
外源物质对镉胁迫下不同品种冬小麦苗期镉吸收特征的影响
2021-01-27 00:59李乐乐李中阳吴大付班卓昊李宝贵赵志娟
灌溉排水学报 2021年1期
李乐乐,李中阳,吴大付,班卓昊,李宝贵,樊 涛,胡 超,赵志娟,刘 源
外源物质对镉胁迫下不同品种冬小麦苗期镉吸收特征的影响
李乐乐2,李中阳1,吴大付2,班卓昊3,李宝贵1,樊 涛1,胡 超1,赵志娟1,刘 源1*
(1.中国农业科学院 农田灌溉研究所,河南 新乡 453002;2.河南科技学院,河南 新乡 453003;3.长垣职业中等专业学校,河南 新乡 453400)
【】分析不同外源物质对不同品种冬小麦苗期Cd吸收和迁移特征的影响及其差异性。通过向含不同质量浓度Cd(10、30 mg/L)的营养液中添加不同质量浓度的Si(28、56 mg/L)、Ca(50、100 mg/L)、Mg(50、100 mg/L)和腐殖酸(5、15 mg/L),在水培条件下研究了籽粒Cd高积累百农419和低积累百农418小麦苗期植株Cd吸收和转运特征、植株对Ca和Mg的吸收以及根系形态指标的变化。不同品种冬小麦对Cd的吸收存在差异。随着营养液中Cd质量浓度的升高,不同品种冬小麦根系生长受抑制程度更严重。与百农418相比,百农419是喜Ca品种。在低Cd质量浓度时,与CK相比,低Si添加改善百农419根系生长和降低植株Cd量的效果最好,但其他物质添加抑制了其根系生长且高质量浓度腐殖酸处理增加了根系中Cd的累积,同时所有外源物质添加均降低了其根系Ca量;对于百农418来说,加Si和Ca可以促进根系生长且低Si效果最明显,加Mg和腐殖酸对根系生长影响不明显,加Si显著降低了根系和茎叶Cd量,加Ca和Mg只显著降低了根系Cd量,加腐殖酸对植株Cd量无显著影响。在高Cd质量浓度时,添加Si可以促进2种小麦根系生长并降低根系和茎叶Cd量,其中低Si和高Si分别对百农419和418根系生长促进效果更好;而其他外源物质添加对Cd毒害基本无明显缓解效果。与其他处理相比,低质量浓度Cd条件下高Si显著增加了2种小麦的Cd转运系数,高质量浓度Cd条件下高Si显著增加了百农419的Cd转运系数。相比其他外源物质,添加Si对冬小麦Cd毒害缓解效果最明显,且品种、Cd质量浓度和Si质量浓度交互作用明显。
小麦;硅;镉;钙;镁;腐殖酸
0 引言
2014年环保部和国土资源部的联合公报表明全国土壤环境状况总体不容乐观,部分地区土壤污染较重,耕地土壤环境质量堪忧,其中重金属Cd的污染物点位超标率在所有无机污染物中超标率最高[1]。【研究意义】当作物组织中的Cd积累到一定程度时,会使作物出现生长迟缓、产量下降等症状,严重时甚至会造成作物死亡[2]。Cd还会通过食物链进入人体内,对人体健康造成极大影响[3-5]。添加外源物质可以缓解重金属Cd对作物的毒害作用,减少Cd在作物体内的富集[6-8]。
【研究进展】已有研究表明不同品种冬小麦对重金属Cd具有不同的耐受性,通过筛选重金属Cd低积累冬小麦品种,可以有效缓解重金属Cd污染问题[9-12]。研究发现,Cd对苗期小麦具有严重的毒害作用,而添加不同的外源物质会缓解这种毒害现象[13-14]。Ca是植物生长必需的营养元素,能够作为第二信使与CaM(Calmodulin)结合偶联胞外信号与胞内生理生化反应,通过抑制Cd的吸收,促进叶片光合作用及气体交换速率,维持植物体的含水率、植物叶片光合色素量及矿质营养的平衡,稳定细胞壁、细胞膜结构及诱导特异性基因表达等途径来提高植物对重金属Cd毒害等逆境的抗性[15]。同时,Ca可与Cd形成较稳定的与土壤结合的闭蓄态Cd,从而抑制Cd进入生物体;另一方面,Ca可与Cd竞争进入植物细胞,从而降低植物细胞Cd量[16]。Mg也是植物生长必需的营养元素,参与植物根的形成、叶绿素和光合作用的产生以及酶的活化等。Mg还可影响Cd在土壤中的赋存形态以及在植物体内的累积和转运,进而缓解Cd对植物的毒害作用[17]。Si提高植物对重金属胁迫抗性的可能机理有以下几方面:①Si促进根系分泌草酸,降低了Cd在植物体的积累[18];②Si与重金属可形成沉淀,降低了重金属的移动性;③Si促进Ca的吸收和转运;④Si提高了植物抗氧化系统的防御能力[19]。腐殖酸可与Cd结合形成稳定的络合物,从而减少生物可利用态Cd,降低植物对Cd的吸收;另外,腐殖酸的添加还可改变土壤结构,进而影响土壤中一系列生化反应,间接影响Cd的毒害作用[20-21]。Zhu等[22]和李丽君等[23]研究发现,使用腐殖酸可以明显降低土壤中有效态Cd量,并且提高作物的生物量,降低作物中Cd的积累。【切入点】添加不同的外源物质对Cd毒害的缓解作用可能与作物的种类、品种、生长条件等一系列因素有关,前人研究多关注单一外源物质或单一植物品种,而对添加不同外源物质影响富Cd能力不同的冬小麦品种Cd吸收和迁移特征的研究较少。【拟解决的关键问题】本试验选用前期研究发现的籽粒Cd积累差异明显的2个小麦品种(籽粒Cd高积累品种百农419和籽粒Cd低积累品种百农418),通过水培试验,研究不同外源物质对不同品种冬小麦苗期Cd吸收和富集特征的影响,为重金属Cd污染的修复和治理提供理论依据及参考。
1 材料与方法
1.1 试验材料与设计
根据前期Cd积累冬小麦品种筛选的大田试验结果选用了2种小麦(百农419和百农418)进行试验。选择籽粒饱满的小麦种子在75%酒精中浸泡5 min杀菌,再用蒸馏水洗掉残余的酒精,温水浸泡1 h后,将种子均匀摆放在带网格的育苗盘中催芽。待小麦长到二叶一心期时,将其移栽到配制好的改良霍格兰营养液(购自山东拓普生物工程有限公司)中[24],成分见表1。每8株小麦放置在1个盛1 L营养液的灰色PVC小桶(直径10 cm,高20 cm)中。营养液用400 g/L NaOH溶液调节pH值至6.0,小麦在营养液培养2 d后开始试验。鉴于目前我国土壤Cd污染的严峻形势[25-28],本试验营养液中Cd(CdCl2)质量浓度设置了2个水平(10、30 mg/L),缩写为Cd10和Cd30。通过化学平衡模拟软件Visual MINTEQ计算pH值6.0营养液中Cd的形态及质量浓度结果表明2种质量浓度下加入的Cd约99.999%存在于溶液中,基本无沉淀。在不同水平的含Cd营养液中添加不同的外源物质,每种外源物质设置2个水平。外源物质Si(Na2SiO3·9H2O)的质量浓度设置为28 mg/L(S1)、56 mg/L(S2),Ca(CaCl2)的质量浓度设置为50 mg/L(C1)、100 mg/L(C2),Mg(MgCl2)的质量浓度设置为50 mg/L(M1)、100 mg/L(M2),腐殖酸(90%黄腐酸)质量浓度设置为5 mg/L(F1)、15 mg/L(F2)。同时在2种Cd质量浓度下设置只添加Cd不添加外源物质的对照(CK),另外还设置了不加Cd不加外源物质的正常营养液CK(Cd0)。共38个处理,每个处理4次重复,总计152个小桶。水培试验在人工气候室中进行,温度25 ℃,光照时间为08:00—18:00。每3天更换1次营养液,处理30 d后收样。
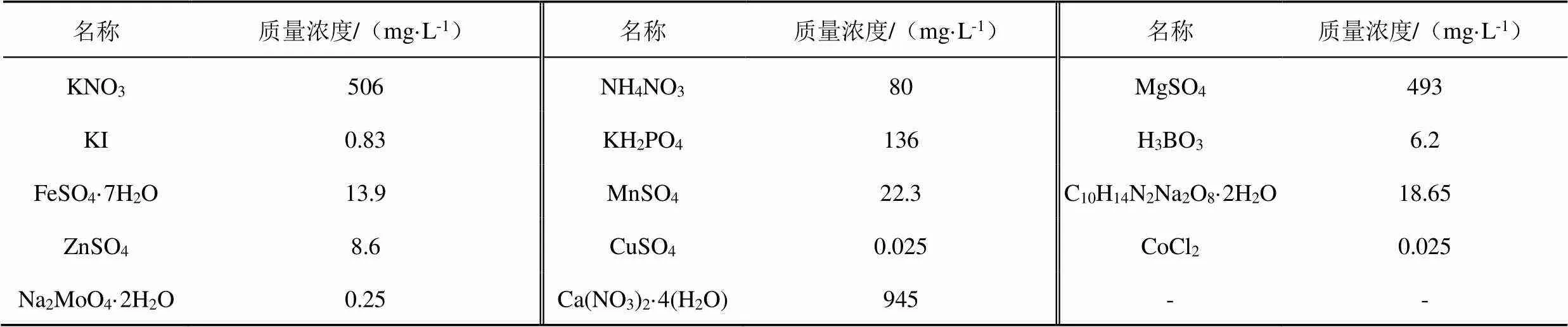
表1 改良霍格兰营养液成分
1.2 测定项目与方法
试验处理30 d后将样品从营养液中取出,用蒸馏水冲洗后分成根和茎叶[29]。茎叶先在烘箱中105 ℃杀青0.5 h,然后在70 ℃下烘干至恒质量。每个处理随机挑选3株小麦根系,用WinRHI-ZO 系列植物根系扫描仪进行扫描,得出3株小麦根系总长度、根系总表面积、根系总体积和根尖数之和。然后所有根系烘干,烘干后的样品用粉碎机(Taisite FW100)磨碎并充分混合均匀装袋。消煮时称取0.200 g植物样加入10 mL浓HNO3,用微波消解仪(Mars CEM 240/50)进行消解,同时做4个空白。消煮液赶酸定容后用原子吸收分光光度计(火焰+石墨炉)(PinAAcle 900,PerkinElmer,美国)测定Cd量。转运系数=茎叶含Cd量/根系含Cd量。
1.3 数据处理
采用Excel 2007对数据进行整理和作图,采用SAS 9.2软件进行单因素方差分析(One-way ANOVA),并对不同处理间的差异进行Duncan多重比较,显著性水平为=0.05。采用SPSS 16.0进行三因素方差分析。
2 结果与分析
2.1 不同外源物质对冬小麦根系形态的影响
小麦根系形态指标结果如表2和表3所示,所有指标均为3株小麦一起扫描的结果。在不加Cd不加外源物质的正常营养液中(表2),2种小麦品种的根系生长无显著性差异。添加不同的外源物质对小麦根系形态指标的影响如表3所示。由表3可知,与正常营养液中根系各形态指标相比,添加2种质量浓度的Cd显著抑制了2种小麦根系生长。在只添加低质量浓度Cd的营养液中,百农419的根系生长显著优于百农418;但Cd高质量浓度时二者无显著差异。
在Cd质量浓度为10 mg/L时,与CK相比,加Si增加了百农419总根长、总根表面积、总根体积和总根尖数,且在低质量浓度Si处理达显著水平,增幅分别为92.17%、102.75%、113.73%、71.08%;在加Ca、Mg、腐殖酸条件下百农419各根系形态指标均显著下降,加高质量浓度腐殖酸时降幅最大,分别为75.81%、68.46%、58.80%、70.16%。添加外源物质后百农418各根系形态指标相比CK均上升,加Mg和腐殖酸处理增加不显著,加Si和Ca处理达到显著水平,Si的效果显著优于Ca,且低质量浓度Si增加效果显著优于高质量浓度,低质量浓度Si时总根长、总根表面积、总根体积和总根尖数分别增加了292.94%、304.92%、320.75%、312.19%。

表2 正常营养液中不同小麦品种根系形态指标
注 表中同列不同小写字母代表处理间差异显著(0.05)。
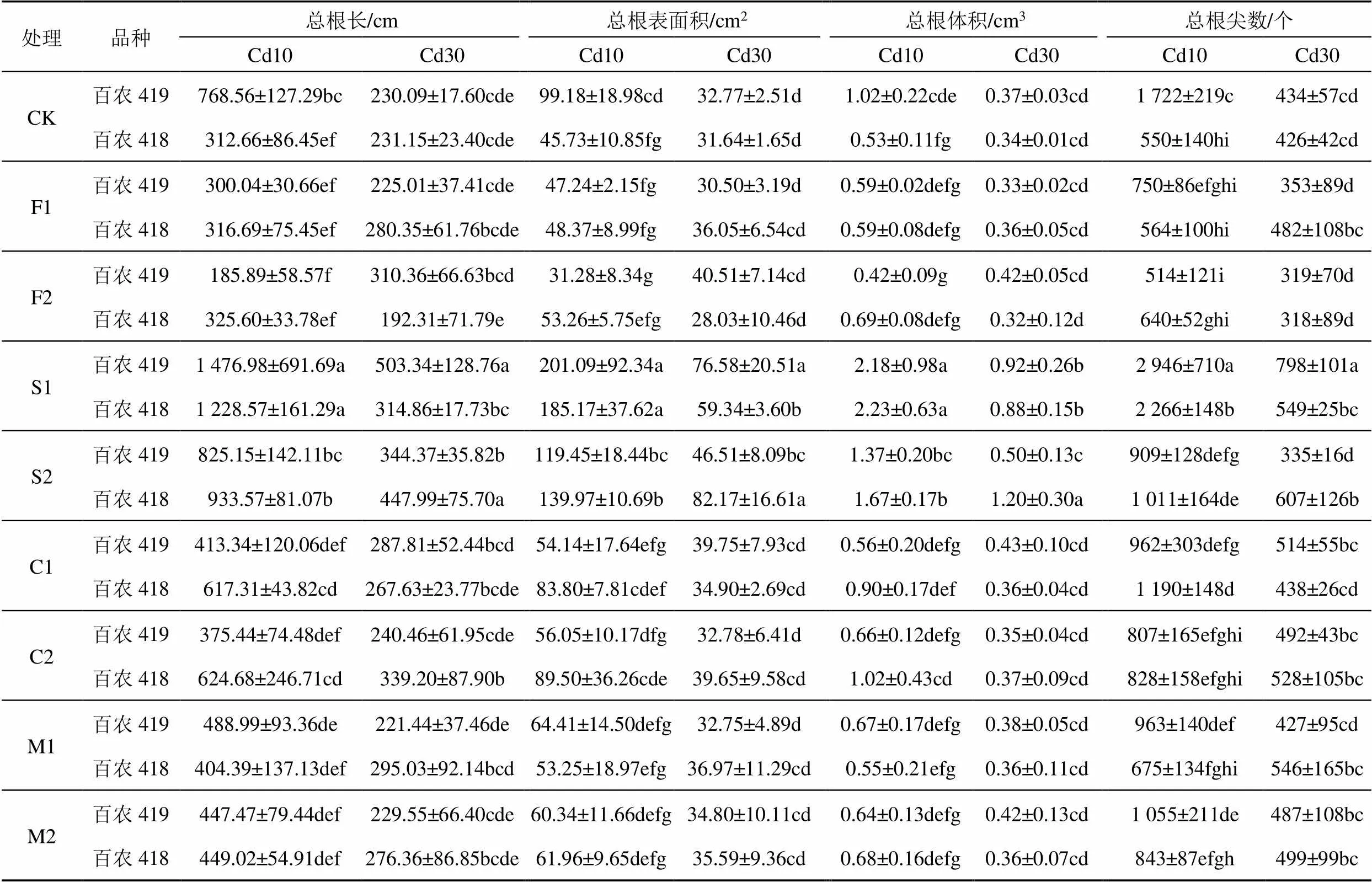
表3 添加不同外源物质或Cd时不同小麦品种根系形态指标
注 表中同列不同小写字母代表处理间差异显著(0.05)。
当Cd质量浓度为30 mg/L时,百农419低Si处理各根系形态指标显著优于不添加外源物质的CK处理,增幅分别为118.75%、133.68%、148.65%、83.76%;高质量浓度Si相比CK只显著增加了总根长;添加腐殖酸、Ca、Mg的影响不显著,低质量浓度腐殖酸对根系生长最不利,总根长、总根表面积、总根体积和总根尖数相比CK下降幅度分别为2.21%、6.93%、10.81%、18.66%。与CK相比,只有高质量浓度Si的添加显著改善了百农418根系各形态指标,高Si时总根长、总根表面积、总根体积和总根尖数增幅分别为93.80%、159.70%、252.94%、42.37%。说明添加外源物质Si可以有效缓解根系Cd毒害现象,低质量浓度Si对百农419两种质量浓度Cd毒害以及百农418低质量浓度Cd毒害的缓解效果较好,高质量浓度Si对百农418高质量浓度Cd毒害的缓解效果较好。
品种、Cd质量浓度和相同外源物质3个质量浓度的三因素方差分析结果如表4所示。从表4可以看出添加Si条件下,不同处理间根系总根长和总根表面积的差异主要是由Cd质量浓度和外源物质量浓度造成的。添加腐殖酸条件下,品种、Cd质量浓度和腐殖酸质量浓度共同造成了根系形态指标的差异。添加Ca条件下,差异主要是由Cd质量浓度造成的。添加Mg条件下,差异主要是由品种和Cd质量浓度造成的。
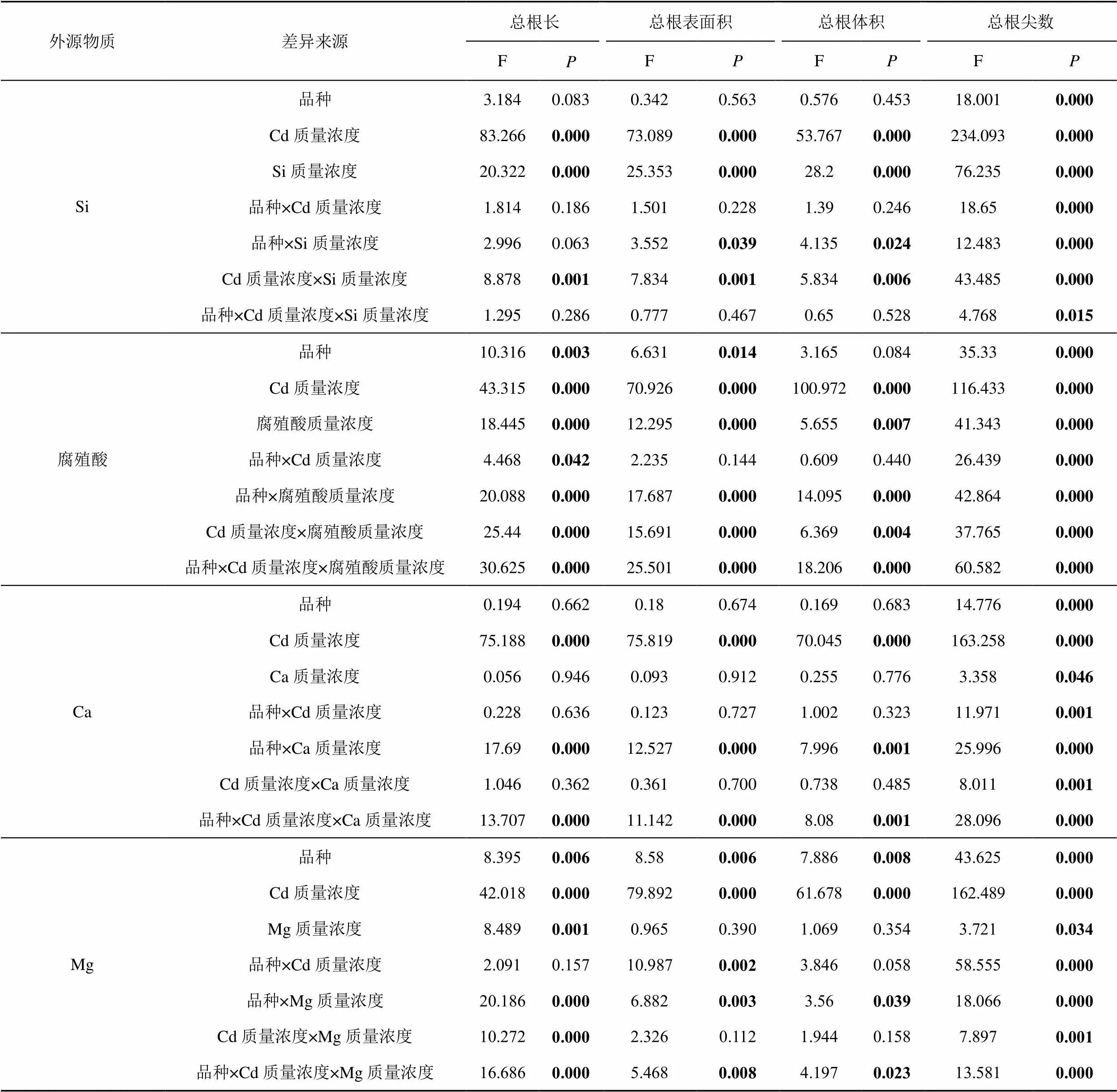
表4 根系形态指标的三因素方差分析
注 加粗的数值代表有显著差异,其中<0.05代表差异显著,<0.01代表差异极显著。
2.2 不同外源物质对冬小麦植株Ca和Mg量的影响
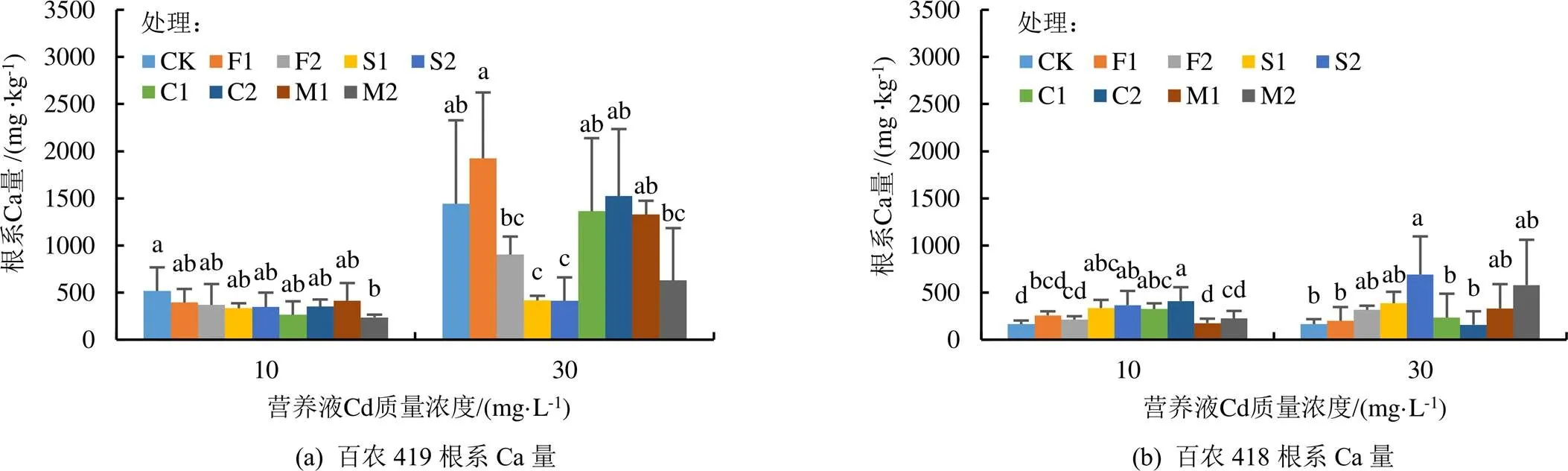
不同处理条件下小麦根系Ca和Mg量图1(a)—图1(d)所示。在正常营养液中生长的百农419和百农418根系Ca量分别为610 mg/kg和360 mg/kg,根系Mg量分别为999 mg/kg和583 mg/kg,百农419根系的Ca和Mg量明显高于百农418,说明百农419相比于百农418更喜Ca和Mg。在低Cd条件下,与CK相比,添加外源物质均降低了百农419根系Ca量,对百农419和百农418根系Mg量基本无影响(除了高Si处理),却增加了百农418根系Ca量(除个别例外)。高Cd条件下,与CK相比,2种质量浓度Si处理均降低了百农419根系Ca和Mg量,而增加了百农418根系Ca量;而所有外源物质添加对百农418根系Mg量无显著影响。
不同处理条件下小麦茎叶Ca和Mg量图1(e)—图1(h)所示。在正常营养液中生长的百农419和百农418茎叶的Ca量分别为1 082 mg/kg和838 mg/kg,茎叶Mg量分别为3 214 mg/kg和2 931 mg/kg。与CK相比,2种Cd质量浓度下不同外源物质添加对百农419茎叶Ca和Mg量基本无显著影响,除了高Cd条件下高Si处理显著增加了百农419茎叶Ca量。2种Cd质量浓度下,添加外源物质的百农418茎叶Mg量与CK无显著差异。在低Cd条件下,高Si和低Ca处理的百农418茎叶Ca量显著高于CK;高Cd条件下,低Ca和高Ca处理的百农418茎叶Ca量显著高于CK。
2.3 不同外源物质对冬小麦吸收重金属Cd的影响
添加不同外源物质对小麦根系和茎叶Cd量的影响如图2所示。当营养液中不添加外源物质与Cd时,百农419和百农418根系和茎叶中均未检测到Cd。添加不同的外源物质对2种小麦根系和茎叶Cd量影响不同。随着Cd质量浓度的升高,2种冬小麦根系和茎叶Cd量均上升。从图2(a)可以看出,在Cd质量浓度为10 mg/L时,与CK相比,外源Si显著抑制了百农419根系对Cd的吸收,高Si降幅最大为85.62%;加Ca、Mg的根系Cd量与CK无显著差异;加腐殖酸反而促进了根系对Cd的吸收,高腐殖酸时根系Cd量相比CK增加最多(43.48%),加重了Cd毒害作用,与根系形态结果一致。在Cd质量浓度为30 mg/L时,添加2种质量浓度的Si以及高质量浓度Mg显著抑制了百农419根系对Cd的吸收,高Si抑制作用最强,相比CK降低了79.17%;其余处理对根系Cd量无显著影响。这与根系形态结果不一致,说明高Si虽然没有明显改善百农419的根系形态,却显著降低了根系吸收的Cd。
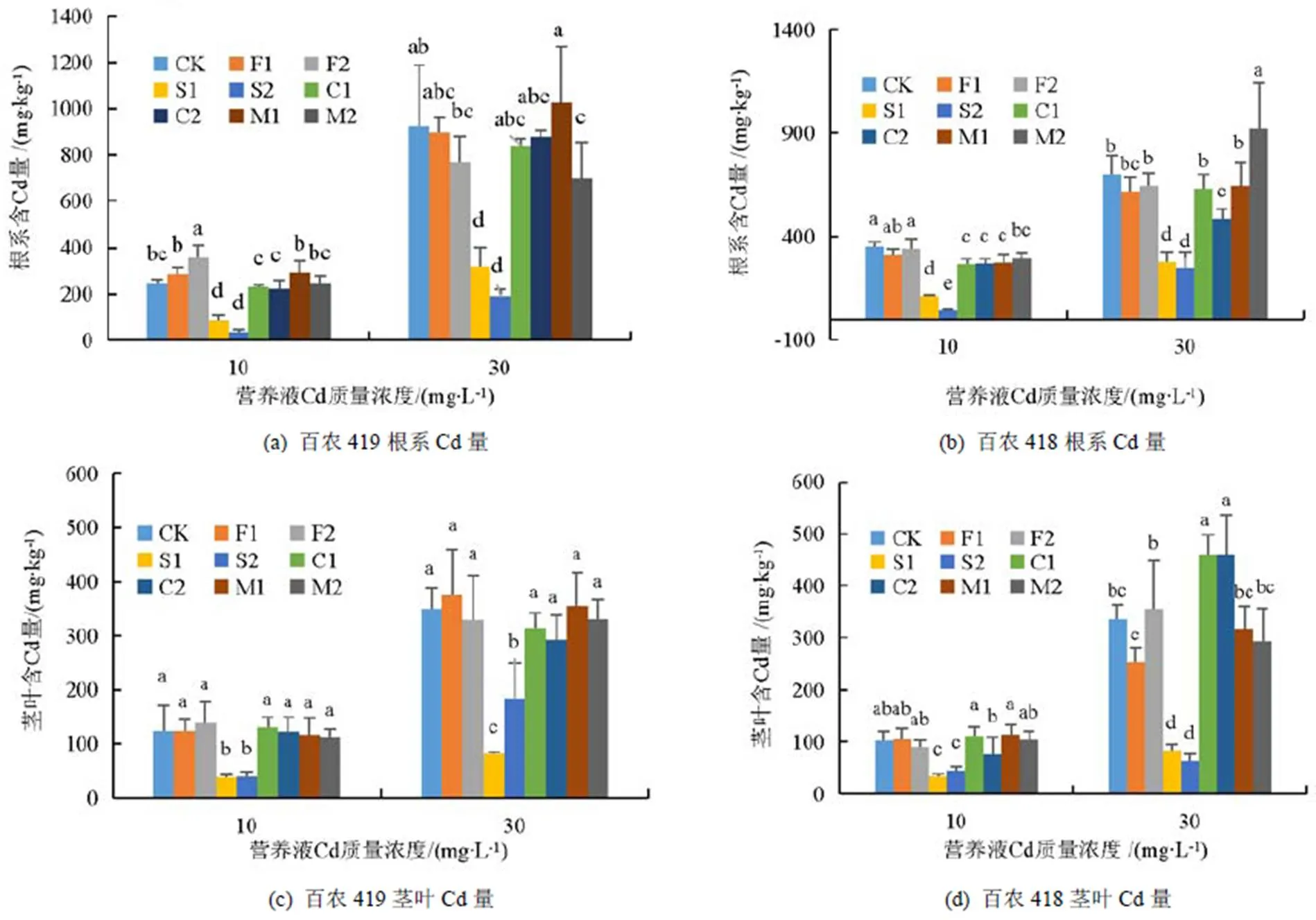
图2 添加不同外源物质时小麦根系和茎叶Cd量
不同外源物质对百农418根系Cd量影响的结果如图2(b)所示。在Cd质量浓度为10 mg/L时,除腐殖酸外,其他处理相比CK均显著降低了百农418根系Cd量,高Si的降幅最大为87.59%。在Cd质量浓度为30 mg/L时,添加2种质量浓度Si相比CK显著降低了百农418根系Cd量,高Si降幅最大为64.24%;另外高Ca处理显著降低了根系Cd量、高Mg处理显著增加了根系Cd量。从图2(c)可以看出,在Cd质量浓度为10 mg/L时,相比CK加Si显著降低了百农419茎叶Cd量,但2种Si质量浓度无显著差异,其他处理影响不显著;Cd质量浓度为30 mg/L时不同处理与CK的差异与低质量浓度Cd类似,但低Si对百农419茎叶含Cd量的降低作用(76.87%)显著大于高Si。不同外源物质对低质量浓度Cd条件下百农418茎叶Cd量的影响与百农419相似(图2(d)),相比CK低Si处理对茎叶Cd量降低作用(68.07%)最明显。在Cd质量浓度为30 mg/L时,相比CK加Ca显著增加了百农418茎叶Cd量,最大增幅为36.32%(C1);加Si处理显著降低了茎叶含Cd量,最大降幅为88.18%(S2);其余处理条件下无显著影响。
植株Cd量的三因素方差分析结果如表5所示。由表5可知,添加Si条件下,根系Cd量差异主要是Cd质量浓度和Si质量浓度造成的,茎叶Cd量差异主要是由品种、Cd质量浓度和Si质量浓度造成的。添加腐殖酸条件下,根系Cd量差异主要由是品种和Cd质量浓度引起的,而茎叶Cd量差异主要是由Cd质量浓度引起的。添加Ca条件下,根系Cd量差异主要是由品种、Cd质量浓度和Ca质量浓度导致的,茎叶Cd量差异主要是由品种和Cd质量浓度导致的。添加Mg条件下Cd质量浓度是造成植株Cd量差异的主要因素。

表5 植株Cd量和Cd转运系数的三因素方差分析
注 加粗的数值代表有显著差异,其中<0.05代表差异显著,<0.01代表差异极显著。
2.4 不同外源物质对冬小麦Cd转运系数的影响
由图3可知,在Cd质量浓度为10 mg/L时,百农419的转运系数普遍高于百农418,高Si处理条件下百农419和百农418转运系数>1,显著高于其他处理条件下百农419和百农418转运系数。在Cd质量浓度为30 mg/L时,只有高Si处理百农419转运系数>1,显著高于其他处理条件下百农419转运系数,百农418在所有处理条件下转运系数均<1。
植株Cd转运系数的三因素方差分析结果如表5所示。由表5可知,添加Si条件下,转运系数差异主要是由品种、Cd质量浓度和Si质量浓度造成的。添加腐殖酸条件下,转运系数差异主要是Cd质量浓度引起的。添加Ca条件下,转运系数差异主要是源于Cd质量浓度和Ca质量浓度差异。
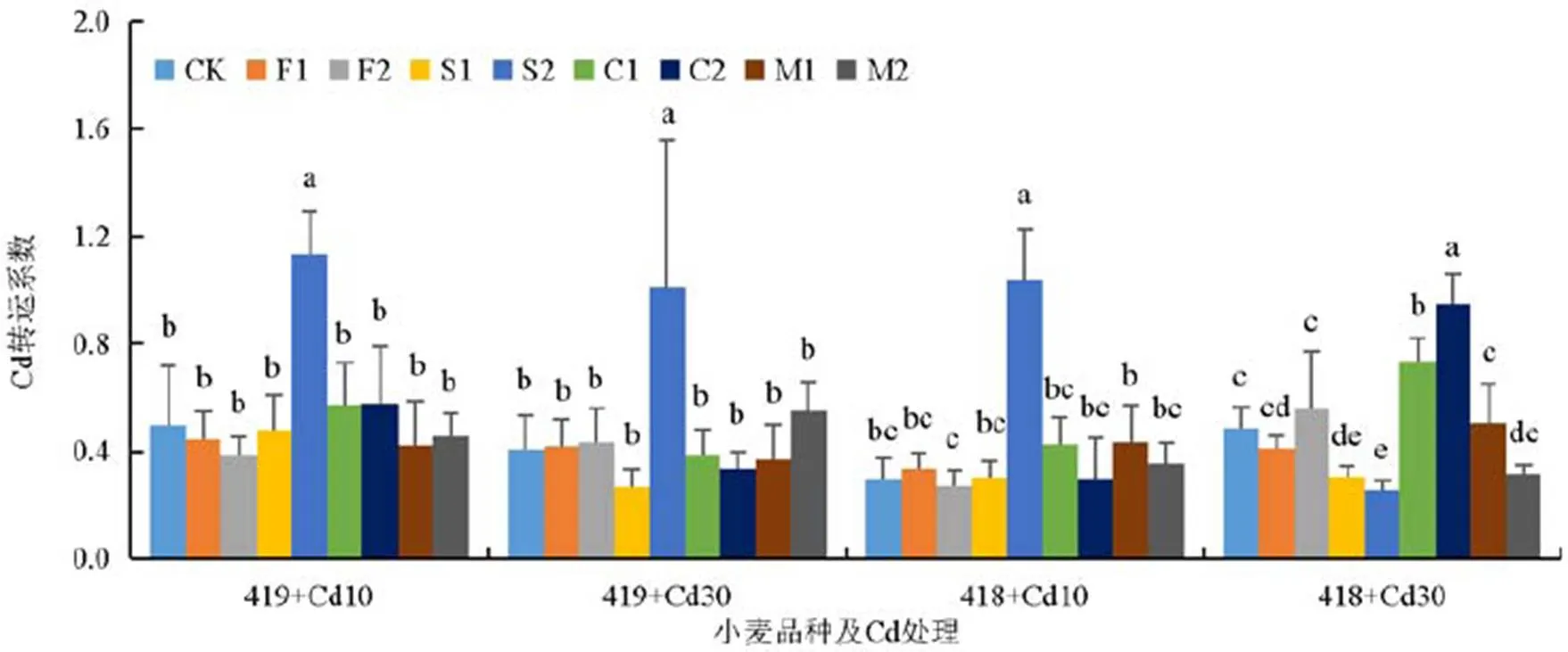
图3 添加不同外源物质时不同品种冬小麦Cd转运系数
3 讨论
根系形态指标是评价作物对Cd耐性的重要指标。当作物生长环境中Cd质量浓度过高会导致作物生长缓慢并矮化,而添加不同外源物质可以缓解这一现象。本研究中2种小麦都是敏感品种,表现为在Cd处理条件下根系生长都受到了明显的抑制,且随着Cd质量浓度的升高,抑制作用更明显,根系以及茎叶含Cd量明显上升。在低Cd质量浓度时,与CK相比,百农418在各处理条件下各根系形态指标均改善,改善效果表现为:Si>Ca>Mg>腐殖酸;而百农419只在Si处理条件下各根系形态指标优于CK。在高Cd质量浓度时,不同外源物质对2种小麦品种的影响也不同。造成这种差异的原因主要是种内差异[30]。二者对Cd毒害的解毒机制差异可能是导致这种差异的原因之一。吸收到植物体内的Cd会启动植物的自我保护系统以降低Cd在植株体内的转运和累积,而这种自我保护系统在不同植物或品种的差异会造成植物的不同器官和不同品种间Cd的积累量不同[31]。在本研究中体现为低Cd无外源物质的CK下,百农418根系Cd量明显高于百农419,而百农419的转运系数高于百农418。另一个可能的原因是二者对Ca的需求量不同。百农419需Ca量较多,但所有外源物质添加均导致了根系Ca量减少,其中腐殖酸处理还导致根系累积Cd增加,而百农418根系在添加外源物质后Ca量未下降且Cd量下降,因此百农419的根系生长相对于CK受抑制作用明显,百农418的根系未受到明显影响。另外,不同植物或品种根系阳离子交换量、Zeta电位等带电性质的差异会影响根系对Cd的吸附和吸收[32],环境条件的改变也会导致植物不同品种对Cd的耐受能力的差异[8]。因此,依据小麦不同品种Cd吸收的差异特性[10-11,33]可以在筛选重金属低积累小麦品种的基础上施用合适的外源物质并配合适当的栽培措施降低小麦重金属超标的风险[34]。
本研究比较了4种外源物质对Cd毒害的缓解效果,但4种物质的效果在2种小麦品种表现不尽相同。不同外源物质中Si对不同品种冬小麦根系和茎叶Cd量降低效果最好,但添加Ca、Mg和腐殖酸并不总是缓解Cd毒害。研究表明,水稻幼苗在Ca质量浓度较低时,Cd在根部和茎叶的量高于Cd单独处理时[35],说明Ca在一定质量浓度会促进作物对Cd的吸收,加重Cd的毒害。熊礼明等[36]研究也证明了上述结果。朱华兰[37]研究表明,缺Mg和Mg过量均不能缓解玉米对Cd的吸收,即Mg对Cd毒害的缓解作用取决于添加Mg的质量浓度。孙枭琼等[38]研究表明,腐殖酸钠对冬小麦种子Cd毒害的缓解作用也依赖于添加的质量浓度。这也部分解释了本研究在一定Cd质量浓度胁迫下添加Ca、Mg和腐殖酸反而降低了各根系形态指标。
转运系数体现出植物地上部转运根部重金属的能力,系数的高低将直接影响植物抵御重金属毒害的能力。由图3可知,在CK、F1、F2、S1、C1、C2、M1和M2处理下2种小麦转运系数均<1,说明Cd主要积累在根系中,这与前人研究结果[12]相同。而在高质量浓度Si处理下不同品种冬小麦转运系数>1,说明加入高质量浓度Si抑制了根系对Cd的吸收,却提升了Cd向茎叶转运的能力。也有研究表明加Si可降低Cd转运系数[39],与本研究结论不一致,可能是由于本试验Si添加量较高,而且本研究也证实2种Si添加量下Cd转运系数有显著差异。Si并不总是降低Cd转运系数。有研究表明加Si后Cd在水稻各部位的转运系数因土壤pH而异[40]。另一个关于大蒜的研究中也发现加Si提高了Cd的转移系数[41]。另外,加Si对重金属转运系数的影响随着生育期的推进而改变[42]。说明Si对Cd在植物体内转运的影响还有待进一步研究。另外,低Cd时百农419表现出较强的Cd转运能力,这与前期大田试验结果[43]一致,说明在Cd污染土壤中种植百农419有较高风险,可能会导致地上部分和籽粒积累较高的Cd。
4 结论
百农419和百农418均为Cd敏感小麦品种,百农419比百农418对Ca的需求更高。
在Cd质量浓度为10 mg/L时,对于百农419来说,加Si缓解了Cd毒害,但加其他物质无缓解作用;对于百农418来说,加Si同样缓解了Cd毒害,加Ca促进了根系生长且降低了根系Cd量,加Mg只降低了根系Cd量,加腐殖酸无明显作用。
在Cd质量浓度为30 mg/L时,加Si促进了2种小麦根系生长并降低了植株Cd量,其中低Si和高Si分别对百农419和百农418根系生长促进效果更好;而其他物质添加对Cd毒害基本无明显缓解效果。
高Si显著增加了低Cd条件下2种小麦品种的Cd转运系数以及高Cd条件下百农419的Cd转运系数。
因此,不同外源物质对2种小麦品种Cd毒害缓解效果最好的为Si,且存在品种、Cd质量浓度和Si质量浓度的明显交互作用。
[1] 环境保护部和国土资源部发布全国土壤污染状况调查公报[J]. 资源与人居环境, 2014 (4): 26-27.
Ministry of Environmental Protection and Ministry of Land and Resources release national survey bulletin on soil pollution[J]. Resources Environment Inhabitant. 2014 (4): 26-27.
[2] 薛永, 王苑螈, 姚泉洪, 等. 植物对土壤重金属镉抗性的研究进展[J]. 生态环境学报, 2014, 23(3): 528-534.
XUE Yong, WANG Yuanyuan, YAO Quanhong, et al. Research progress of plants resistance to heavy metal Cd in soil[J]. Ecology and Environment Sciences, 2014, 23(3): 528-534.
[3] 王科, 李浩, 张成, 等. 不同类型土壤下水稻镉的富集特征及与土壤镉量的关系[J]. 四川农业科技, 2018 (11): 38-40.
WANG Ke, LI Hao, ZHANG Cheng, et al. Accumulation of cadmium in rice in different types of soil and its relationship with soil cadmium content[J]. Sichuan Agricultural Science and Technology, 2018(11): 38-40.
[4] 骆翠红. 浅谈土壤镉污染的修复技术[J]. 中国资源综合利用, 2018, 36(2): 73-75.
LUO Cuihong. Discussion on remediation technology of soil cadmium pollution[J]. China Resources Comprehensive Utilization, 2018, 36(2): 73-75.
[5] 张伟杰, 徐建新. 三峡库区干流沉积物重金属质量分数及污染评价[J]. 灌溉排水学报, 2018, 37(5): 99-105.
ZHANG Weijie, XU Jianxin. Assessing contents and pollution of heavy metals within the sediment deposits in the main streams of the Three Gorges Reservoir[J]. Journal of Irrigation and Drainage, 2018, 37(5): 99-105.
[6] MARYAM Haghighi, M. Kafi, P. Fang, et al. Humic acid decreased hazardous of cadmium toxicity on lettuce (Lactuca sativa L.)[J]. Vegetable Crops Research Bulletin, 2010, 72, 49-61.
[7] HUANG Fei, WEN Xiaohui, CAI Yixia, et al. Silicon-mediated enhancement of heavy metal tolerance in rice at different growth stages[J]. International Journal of Environmental Research and Public Health, 2018, 15(10): 2193.
[8] KANU Adam Sheka, ASHRAF Umair, MO Zhaowen, et al. Calcium amendment improved the performance of fragrant rice and reduced metal uptake under cadmium toxicity[J]. Environmental Science and Pollution Research, 2019, 26(24): 24 748-24 757.
[9] 张玉烛, 方宝华, 滕振宁, 等. 应急性镉低积累水稻品种筛选与验证[J]. 湖南农业科学, 2017 (12): 19-25.
ZHANG Yuzhu, FANG Baohua, TENG Zhenning, et al. Screening and verification of rice varities with low cadmium accumulation[J]. Hunan Agricultural Sciences, 2017(12): 19-25.
[10] 肖亚涛, 吴海卿, 李中阳, 等. 不同基因型冬小麦镉累积差异及其与根系形态的关系[J]. 水土保持学报, 2015, 29(6): 276-280.
XIAO Yatao, WU Haiqing, LI Zhongyang, et al. Difference of cadmium accumulation by different genotypes of winter wheat and its relationship with root morphology[J]. Journal of Soil and Water Conservation, 2015, 29(6): 276-280.
[11] 林小兵, 周利军, 王惠明, 等. 不同水稻品种对重金属的积累特性[J]. 环境科学, 2018, 39(11): 5 198-5 206.
LIN Xiaobing, ZHOU Lijun, WANG Huiming, et al. Accumulation of heavy metals in different rice varieties[J]. Environmental Science, 2018, 39(11): 5 198-5 206.
[12] 邓洪, 刘惠见, 牛婧, 等. 玉米重金属低累积品种的筛选与研究[C]// 中国土壤学会土壤环境专业委员会第二十次会议暨农田土壤污染与修复研讨会摘要集, 合肥, 2018: 34.
DENG Hong, LIU Huijian, NIU Jing, et al. Screening and Study of Maize Heavy Metal Low Accumulation Varieties[C]// The 20th Meeting of Soil Environment Professional Committee of Chinese Soil Society and Symposium on Farmland Soil Pollution and Remediation. Hefei, 2018: 34.
[13] 周相玉, 冯文强, 秦鱼生, 等. 镁、锰、活性炭和石灰及其交互作用对小麦镉吸收的影响[J]. 生态学报, 2013, 33(14): 4 289-4 296.
ZHOU Xiangyu, FENG Wenqiang, QIN Yusheng, et al. Effects of magnesium, manganese, activated carbon and lime and their interactions on cadmium uptake by wheat[J]. Acta Ecologica Sinica, 2013, 33(14): 4 289-4 296.
[14] 师振亚. 硅对小麦幼苗镉毒害的缓解作用研究[D]. 郑州: 河南农业大学, 2018.
SHI Zhenya. Allevition of exogenous silion on cadmium toxicity in wheat seedlings[D]. Zhengzhou: Henan Agricultural University, 2018.
[15] 王巧玲, 邹金华, 刘东华, 等. 外源钙(Ca)对毛葱耐镉(Cd)胁迫能力的影响[J]. 生态学报, 2014, 34(5): 1 165-1 177.
WANG Qiaoling, ZOU Jinhua, LIU Donghua, et al. Effects of exogenous calcium (Ca) on tolerance of Allium cepa var. agrogarum L. to cadmium (Cd) stress[J]. Acta Ecologica Sinica, 2014, 34(5): 1 165-1 177.
[16] 杜文琪. 外源镁对镉在稻田系统中生物有效性与转运累积的影响[D]. 长沙: 中南林业科技大学, 2018.
DU Wenqi. Effects of exogenous magnesium on bioavailability, transportation and accumulation of cadmium in rice-soil system[D]. Changsha: Central South University of Forestry and Technology, 2018.
[17] KASHEM Md Abul, KAWAI Shigenao. Alleviation of cadmium phytotoxicity by magnesium in Japanese mustard spinach[J]. Soil Science and Plant Nutrition, 2007, 53(3): 246-251.
[18] WU Jiawen, CHRISTOPH-MARTIN Geilfus, BRITTA Pitann, et al. Silicon-enhanced oxalate exudation contributes to alleviation of cadmium toxicity in wheat[J]. Environmental and Experimental Botany, 2016, 131: 10-18.
[19] 宋阿琳. 小白菜对镉胁迫的响应及硅缓解镉毒害的机制[D]. 南京: 南京农业大学, 2009.
SONG Alin, Responses of Brassica chinensis L. to cadmium stress and silicon-alleviated cadmium toxicity[D]. Nanjing: Nanjing Agricultural University, 2009.
[20] NAGASAWA Kenya, WANG Binhui, NISHIYA Kazuki, et al. Effects of humic acids derived from lignite and cattle manure on antioxidant enzymatic activities of barley root[J]. Journal of Environmental Science and Health, Part B, 2016, 51(2): 81-89.
[21] KHAN Kiran Yasmin, ALI Barkat, CUI Xiaoqiang, et al. Effect of humic acid amendment on cadmium bioavailability and accumulation by pak choi (Brassica rapa ssp chinensis L.) to alleviate dietary toxicity risk[J]. Archives of Agronomy and Soil Science, 2017, 63(10): 1 431-1 442.
[22] ZHU Hanhua, CHEN Cheng, XU Chao, et al. Effects of soil acidification and liming on the phytoavailability of cadmium in paddy soils of central subtropical China[J]. Environmental Pollution, 2016, 219: 99-106.
[23] 李丽君, 张强, 白光洁, 等. 改良剂与油菜对土壤重金属有效态的影响[J]. 水土保持学报, 2014, 28(1): 246-252.
LI Lijun, ZHANG Qiang, Bai Guangjie, et al. The Influence of amendments and rape on available heavy metals content in soil[J]. Journal of Soil and Water Conservation, 2014, 28(1): 246-252.
[24] 贾瑞星, 丁鑫超, 汤丹峰, 等. 镉对两个同核异质红麻杂交种种子萌发及幼苗生长的影响[J]. 南方农业学报, 2019, 50(8): 1 688-1 694.
JIA Ruixing, DING Xinchao, TANG Danfeng, et al. Effects of seed germination and seedling growth of two homonuclear- heterocytoplasmic kenaf hybrid cultivars under cadmium stress[J]. Journal of Southern Agriculture, 2019, 50(8): 1 688-1 694.
[25] 李艳玲, 卢一富, 陈卫平, 等. 工业城市农田土壤重金属时空变异及来源解析[J]. 环境科学, 2020, 41(3): 1 432-1 439.
LI Yanling, LU Yifu, CHEN Weiping, et al. Spatial-temporal variation and source change of heavy metals in the cropland soil in industrial city[J]. Environmental Science, 2020, 41(3): 1 432-1 439.
[26] 王敏, 蒋泽海. 环境卫生监测项目土壤中镉的检测结果分析[J]. 世界有色金属, 2019, 19: 283-284.
WANG Min, JIANG Zehai. Analysis on the detection results of cadmium in soil of 2018 rural environmental health monitoring project in Liupanshui City[J]. World Nonferrous Metal, 2019, 19: 283-284.
[27] 陈兆进, 李英军, 邵洋, 等. 新乡市镉污染土壤细菌群落组成及其对镉固定效果[J]. 环境科学, 2020, 41(6): 2 889-2 897.
CHEN Zhaojin, LI Yingjun, SHAO Yang, et al. Bacterial Community composition in cadmium-contaminated soils in Xinxiang city and its ability to reduce cadmium bioaccumulation in Pak Choi (Brassica chinensis L.)[J]. Environmental Science, 2020, 41(6): 2 889-2 897.
[28] 夏雪姣, 菅明阳, 韩玉翠, 等. 镉胁迫对小麦形态发育及生理代谢的影响[J]. 农业生物技术学报, 2018, 26(9): 1 494-1 503.
XIA Xuejiao, JIAN Mingyang, HAN Yucui, et al. Effects of cadmium stress on morphological development and physiological metabolism in wheat (Triticum aestivum)[J]. Chinese Journal of Agricultural Biotechnology, 2018, 26(9): 1 494-1 503.
[29] KIKUCHI Tetsuro, OKAZAKI Masanori, MOTOBAYASHI Takashi. Suppressive effect of magnesium oxide materials on cadmium accumulation in winter wheat grain cultivated in a cadmium- contaminated paddy field under annual rice–wheat rotational cultivation[J]. Journal of Hazardous Materials, 2009, 168, 89-93.
[30] 陈亮妹, 李江遐, 胡兆云, 等. 重金属低积累作物在农田修复中的研究与应用[J]. 作物杂志, 2018(1): 16-24.
CHEN Liangmei, LI Jiangxia, HU Zhaoyun, et al. Review on application of low accumulation crops on remediation of farmland contaminated by heavy metals[J]. Crops, 2018(1): 16-24.
[31] 王丹, 戴绍军. 植物响应金属胁迫重要蛋白质的细胞定位[J]. 生物技术通报, 2010(7): 7-13.
WANG Dan, DAI Shaojun. Cellular localization of proteins in plant in response to heavy metals[J]. Biotechnology Bulletin, 2010(7): 7-13.
[32] ZHOU Qin, LIU Zhaodong, LIU Yuan, et al. Relative abundance of chemical forms of Cu(II) and Cd(II) on soybean roots as influenced by pH, cations and organic acids[J]. Scientific Reports, 2016(6): 36373.
[33] 徐红宁, 杨居荣, 许嘉琳. 作物对Cd的吸收与根系阳离子交换容量[J]. 农业环境保护, 1995(4): 150-153, 177, 193.
XU Hongning, YANG Jurong, XU Jialin. Cd uptake by crops and root cation exchange capacity[J]. Journal of Agro-Environment Science, 1995(4): 150-153, 177, 193.
[34] 何冠华. 不同基因型小麦对土壤重金属污染响应及抗性筛选研究[D]. 郑州: 河南农业大学, 2012.
HE Guanhua. Study on response of different genotypes of wheat to heavy metal pollution soil and resistance screening[D]. Zhengzhou: Henan Agricultural University, 2012.
[35] CHO ShihChueh; CHAO Yunyang, KAO Ching Huei. Calcium deficiency increases Cd toxicity and Ca is required for heat-shock induced Cd tolerance in rice seedlings[J]. Journal of Plant Physiology, 2012, 169(9): 892-898.
[36] 熊礼明, 鲁如坤. 几种物质对水稻吸收镉的影响及机理[J]. 土壤, 1992 (4): 197-200.
XIONG Liming, LU Rukun. Effects of several substances on cadmium uptake in rice and its mechanism[J]. Soils, 1992 (4): 197-200.
[37] 朱华兰. 镉胁迫下不同镁水平对玉米幼苗生长的影响及生理机制的研究[D]. 重庆: 西南大学,2013.
ZHU Hualan. Effect of magnesium levels on growth of cornunder cadmium stress and its physiological mechanisms[D]. Chongqing: Southwest University, 2013.
[38] 孙枭琼, 陈永亮, 孙慎丽, 等. 腐殖酸钠对镉胁迫下冬小麦种子萌发及根系生长的影响[J]. 安徽农业科学, 2017, 45(13): 34-35, 71.
SUN Xiaoqiong, CHEN Yongliang, SUN Shenli, et al. Effect of sodium humate on winter wheat seed germination and root growth under cadmium stress[J]. Journal of Anhui Agricultural Sciences, 2017, 45(13): 34-35, 71.
[39] ZHANG Chaochun, WANG Lijun, NIE Qing, et al. Long-term effects of exogenous silicon on cadmium translocation and toxicity in rice (Oryza sativa L.). Environmental and Experimental Botany, 2008, 62(3): 300-307.
[40] 郭磊. 外源硅影响镉化学形态及其生物有效性的土壤化学机制[D]. 沈阳: 沈阳农业大学, 2018.
GUO Lei. The soil chemistry mechanisms of influences on cadmium chemical speciation and bioavailability with exogenous silicon[D]. Shenyang: Shenyang Agricultural University, 2018.
[41] 王义超. 硅对镉和汞胁迫下大蒜生理生化代谢的影响及机理研究[D]. 杨凌: 西北农林科技大学, 2014.
WANG Yichao. Effects and mechanisms of exogenous silicon on physiological and biochemical metabolisms in garlic under cadmium and mercury stresses[D]. Yangling: Northwest A&F University, 2014.
[42] 苏秀伟. 硅缓解苹果植株高锰毒害的研究[D]. 泰安: 山东农业大学, 2011.
SU Xiuwei. The study of silicon alleviating manganese toxicity in apple trees[D]. Taian: Shandong Agricultural University, 2011.
[43] 李乐乐, 刘源, 李宝贵, 等. 镉低积累小麦品种的筛选研究[J]. 灌溉排水学报, 2019, 38(8): 53-58, 72.
LI Lele, LIU Yuan, LI Baogui, et al. Screening of low-accumulation wheat varieties with cadmium[J]. Journal of Irrigation and Drainage, 2019, 38(8): 53-58, 72.
Cadmium Accumulation in Wheat of Different Varieties at Seedling Stage as Impacted by Addition of Exogenous Elements
LI Lele2, LI Zhongyang1, WU Dafu2, BAN Zhuohao3, LI Baogui1, FAN Tao1, HU Chao1, ZHAO Zhijuan1, LIU Yuan1*
(1. Farmland Irrigation Research Institute, Chinese Academy of Agricultural Sciences, Xinxiang 453002, China; 2. Henan Institute of Science and Technology, Xinxiang 453003, China;3. Changyuan Vocational Secondary Professional School, Xinxiang 453400, China)
【】Cadmium (Cd) is one of contaminants found in agricultural soils caused by anthropogenic activities including wastewater irrigation and application of phosphate fertilizers rich in Cd impurities, sludges and composts. In China, Cd contamination comes to the top in soils contaminated by all heavy metals and their metalloids. Since Cd is toxicto all organisms and highly mobile in soil for plants to take up, excessive Cd accumulation in crop tissues could impede its growth and even lead to mortality. Numerous studies showed that adding exogenous substances to soil could alleviate toxic effects of Cd on crops, but if and how their efficacy varies with crop variety remains poorly understood.【】Taking winter wheat as an example, this paper aimed to investigate the effects of exogenous Si, Ca, Mg and humic acid on uptake of Cd by different cultivars and its subsequent translocation at seedling stage.【】Wheat varieties Bainong 419 (419) with high Cd accumulation in grain and Bainong 418 (418) with low Cd accumulation in grain were taken as the model plants. They were grown in hydroponic culture with the Cd content in it spiked to 10 mg/L or 30 mg/L respectively. We added Si, Ca, Mg and humic acid at different rates to the medium and harvested the crops 30 days later. We then measured Cd accumulation and transportation in roots and shoots, as well as root morphology traits.【】Crop absorption of Cd varied with the wheat varieties, and the total length, surface area, volume and tip number of the roots in both varieties decreased with the increase in Cd concentration. Compared to variety 418, variety 419 took more Ca for its root developments. At low Cd concentration and compared to CK, adding Si at low dose improved root growth of the variety 419 and reduced Cd accumulation in its roots and shoots, while adding other elements inhibited root growth; applying humic acid at high dose enhanced Cd accumulation in the roots. It was found that compared to CK, adding any exogenous element reduced Ca content in the roots of the variety 419 when Cd concentration was low. For the variety 418 grown in medium with low Cd concentration, adding Si and Ca was more effective to promote root growth than adding Si alone, while adding Mg and humic acid did not show noticeable effects. Adding Si reduced Cd accumulation in roots and shoots at significant level, while adding Ca and Mg only impeded Cd accumulation in the root. Humic acid did not appear to have a noticeable impact on plant Cd. For the crops growing in medium with high Cd concentration, adding Si boosted root growth of both varieties regardless of its application rate, while in contrast, adding other elements were unable to alleviate Cd toxicity to plants at significant level. Compared with other treatments, adding Si at high does significantly increased the translocation factor (TF) for both varieties growing in medium with low Cd concentration, and it was also effective at boosting the TF for the variety 419 growing in medium with high Cd concentration.【】The most effective conditioner to alleviate Cd toxicity to winter wheat was Si, although its efficacy varies with wheat cultivar, Si application rate and Cd concentration in the medium where the crop grows.
winter wheat; Si; Cd; Ca; Mg; humic acid
X703.5
A
10.13522/j.cnki.ggps.2019467
1672 - 3317(2021)01 - 0079 - 12
2019-12-26
“十三五”国家重点研发计划项目(2017YFD0801103-2);中国农业科学院基本科研业务费专项所级统筹项目(FIRI2016-14);中国农业科学院基本科研业务费专项院级统筹项目(Y2016XT02);国家自然科学基金青年科学基金项目(41701265)
李乐乐(1994-),男。硕士研究生,主要从事重金属污染土壤修复研究。E-mail: 13939805310@163.com
刘源(1988-),女。副研究员,博士,主要从事非常规水资源安全利用研究。E-mail: liuyuanfiri88@163.com
李乐乐, 李中阳, 吴大付, 等. 外源物质对镉胁迫下不同品种冬小麦苗期镉吸收特征的影响[J]. 2021, 40(1): 79-90.
LI Lele, LI Zhongyang, WU Dafu, et al. Cadmium Accumulation in Wheat of Different Varieties at Seedling Stage as Impacted by Addition of Exogenous Elements[J]. Journal of Irrigation and Drainage, 2021, 40(1): 79-90.
责任编辑:赵宇龙
猜你喜欢
现代园艺(2022年7期)2022-11-19
林业科技(2022年5期)2022-10-08
热带作物学报(2022年7期)2022-08-06
舰船科学技术(2022年11期)2022-07-15
波谱学杂志(2021年3期)2021-09-07
河南农业·综合版(2021年6期)2021-07-16
湖北农业科学(2019年22期)2019-12-23
农民致富之友(2019年8期)2019-05-22
农民致富之友(2019年6期)2019-03-25
食品工业科技(2014年21期)2014-03-11
