
徽州臭鳜鱼微生物多样性、品质特性及其酶解产物抗氧化能力分析
2021-01-20 08:16吴永祥王婷婷张梦婷郭孝成孙汉巨陈向阳
食品科学 2021年2期
吴永祥,王婷婷,张梦婷,张 瑶,郭孝成,孙汉巨,陈向阳,*
(1.黄山学院生命与环境科学学院,安徽 黄山 245041;2.合肥工业大学食品与生物工程学院,安徽 合肥 230009)
臭鳜鱼是我国安徽黄山地区的传统自然发酵食品,是以新鲜鳜鱼为原料,配以一定量的香辛料,经低盐、低温、短期腌制发酵而成,具有悠久的历史和独特的地域风味。经过发酵作用,鳜鱼体内的蛋白质、脂肪及核酸等分子被降解成氨基酸、多肽、脂肪酸、核苷酸及醛、醇、酮等风味物质,营养价值得到提高,风味品质发生变化,并赋予了发酵后鳜鱼似臭非臭、鲜香透骨、鱼肉酥烂的特点[1-2]。目前,徽州臭鳜鱼的加工方法以传统的自然发酵为主,即通过鱼体自身携带的微生物在适宜温度条件下自然发酵形成的,臭鳜鱼的质构特性、特征气味形成与发酵微生物有关,但参与发酵的微生物多样且群落结构复杂,使得发酵过程中的工艺参数不可控、发酵后臭鳜鱼的质量品质不稳定[3]。因此,分析传统发酵臭鳜鱼的微生物多样性对于了解其品质质量至关重要。李燕等[4]采用聚合酶链式反应-变性梯度凝胶电泳(polymerase chain reaction-denaturing gradient gel electrophoresis,PCR-DGGE)技术分析了臭鳜鱼发酵过程中的细菌群落结构,结果发现不同样本臭鳜鱼的微生物群落结构存在一定差异。罗靓芷等[5]采用传统培养的方法分离纯化得到屎肠球菌、弯曲乳杆菌、坚强肠球菌、干酪乳杆菌4 株优良乳酸菌。杨培周等[6]采用传统培养的方法筛选鳜鱼主要发酵微生物,分离鉴定出蜡样芽孢杆菌和苏云金芽孢杆菌。采用传统培养方法获得臭鳜鱼的纯培养菌株非常有限,存在一定的局限性和片面性。高通量测序技术通量高、准确率高,可定性及相对定量解析复杂微生物群落结构体系,是一种有效、便捷的方法,被广泛应用于豆酱、米酒、酒曲、酸奶、泡菜等发酵食品中微生物多样性的研究[7-9]。本研究采用高通量测序技术对发酵前后鳜鱼中细菌群落结构进行对比分析,解析各类微生物的结构比例,明确优势菌群,并从营养成分、理化性质、质构特性等角度分析发酵品质,为准确认识臭鳜鱼的微生物群落结构组成及发酵品质提供数据参考。
酶解技术是食品精深加工的一项绿色加工手段,通过选择性的可控酶解,促进被多种共存杂质干扰的多糖、多肽等活性成分的释放,并可协同微生物发酵,应用于提高发酵后产品的品质[10-11]。包汭琪等[12]研究了木瓜蛋白酶添加对臭鳜鱼挥发性成分和细菌群落结构之间的影响,表明木瓜蛋白酶的添加能显著提高游离氨基酸含量,并促进臭鳜鱼中风味物质的形成。王翠等[13]采用中性酶对臭鳜鱼肉糜进行蛋白质水解,并对影响水解反应的因素如酶的添加量、酶解温度、pH值和液固比进行研究,确定了最佳工艺条件。然而,目前缺乏不同蛋白酶酶解处理对发酵后臭鳜鱼品质影响的深入研究。本研究采用酸性蛋白酶、中性蛋白酶、碱性蛋白酶和木瓜蛋白酶对发酵后臭鳜鱼进行酶解,拟通过微生物发酵、酶解两步法提高不同酶解产物的可溶性多肽含量、增强不同酶解产物清除2,2’-联氮双(3-乙基苯并噻唑啉-6-磺酸)(2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)阳离子自由基、1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基能力及还原能力,以期为徽州臭鳜鱼的微生物与酶协同发酵及提高臭鳜鱼应用价值提供科学依据。
1 材料与方法
1.1 材料与试剂
未发酵鳜鱼和发酵后徽州臭鳜鱼,由安徽徽字一号投资有限公司提供。
DPPH、VC、牛血清白蛋白(bovine serumalbumin,BSA)、ABTS 美国Sigma公司;酸性蛋白酶(100 000 U/g)、碱性蛋白酶(200 000 U/g)沧州夏盛酶生物技术有限公司;中性蛋白酶(50 000 U/g)河南仰韶生化工程有限公司;木瓜蛋白酶(200 000 U/g)南宁庞博生物工程有限公司;蒽酮、铁氰化钾、三氯乙酸、三氯化铁、磷酸氢二钠、磷酸二氢钠 国药集团化学试剂公司。
1.2 仪器与设备
CR-10plus色差仪 日本柯尼卡美能达公司;GY-4数显式硬度计 北京金科利达电子科技有限公司;SpectraMax-190全波长酶标仪 美国Molecular Devices公司;S3400N扫描电子显微镜 日本日立公司;DL-CJ-1N型超净工作台 北京东联哈尔仪器制造有限公司;SQ510C型高压灭菌器 重庆雅马拓科技有限公司;MiSeq高通量测序平台 美国Illumina公司;LabStart-Aw便携式水活度测定仪 瑞士Novasina公司。
1.3 方法
1.3.1 发酵前后鳜鱼微生物宏基因组DNA提取及检测
在无菌超净工作台中,分别剪取发酵前后鳜鱼不同部位鱼肉组织,各自随机混合。称取20 g鱼肉组织,加入50 mL生理盐水,均质5 min。过滤后,取上清液于10 000 r/min离心10 min,弃去上清液,沉淀重悬浮于1 mL TE buffer,-80 ℃保存备用[14]。用Omega细菌基因组DNA提取试剂盒分别提取发酵前后鳜鱼中细菌的总DNA,具体操作步骤见说明书。提取后,用1%的琼脂糖凝胶电泳检测提取效果。
1.3.2 PCR扩增以及高通量测序
细菌16S rDNA的V3-V4区域扩增采用引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)。PCR扩增及高通量测序工作委托上海美吉生物医药科技有限公司进行。
1.3.3 发酵前后鳜鱼理化性质的测定
水分含量的测定:参照GB 5009.3—2016《食品中水分测定》方法;水分活度的测定:采用LabStart-Aw便携式水分活度测定仪,具体操作步骤见说明书;灰分含量的测定:参照GB 5009.4—2016《食品中灰分测定》方法;粗蛋白含量的测定:参照GB 5009.5—2016《食品中蛋白质测定》方法。
可溶性多肽含量的测定:采用双缩脲法[15]。绘制BSA标准曲线,以吸光度(Y)为纵坐标,BSA质量浓度(X/(mg/mL))为横坐标,得到回归方程为:Y=0.079 9X+0.088 8(R2=0.996 3)。根据BSA标准曲线计算样品中多肽含量,结果以每100 g鱼肉中含有相当BSA质量表示,即g/100 g。
可溶性总糖含量的测定:采用蒽酮比色法[16]。绘制葡萄糖标准曲线,以吸光度(Y)为纵坐标,葡萄糖质量浓度(X/(mg/mL))为横坐标,得到回归方程为:Y=3.233 4X+0.008 2(R2=0.998 5)。根据葡萄糖标准曲线计算样品中可溶性总糖含量,结果以每100 g鱼肉中含有相当葡萄糖质量表示,即mg/100 g。
1.3.4 发酵前后鳜鱼质构特性的测定
硬度值采用GY-4型数显式硬度计测定,探头直径为7.9 mm,分别在发酵前后鳜鱼表面随机选取10 个点测量,并记录数据,计算其平均值,单位为kg/cm2。
色泽测定利用CR-10 plus型色差仪,在发酵前后鳜鱼表面随机选取10 个点测量,并记录L*、a*、b*值,计算其平均值。
微观组织结构观察利用S3400N型扫描电子显微镜,将发酵前后鳜鱼鱼肉冻干后,固定在样品托上,采用离子溅射仪在样品的横断面上喷金,扫描电子显微镜(500、2 000 倍)观察并拍照[17]。
1.3.5 发酵臭鳜鱼蛋白酶解液的制备流程
臭鳜鱼肉糜→加入一定量的灭菌水搅拌均匀→酶解→灭酶(沸水浴10 min)→冷却→离心(3 500 r/min,20 min)→取上清液→多肽含量测定
1.3.6 酶解条件的选择
选择酸性蛋白酶、碱性蛋白酶、中性蛋白酶、木瓜蛋白酶作为蛋白质水解酶,分别在各种酶最佳温度、pH值条件下对臭鳜鱼行水解。参考文献[13],统一为加酶量2 000 U/g、料液比1∶20(g/mL)、酶解时间1.5 h。同时设立对照组,为未添加酶处理。
1.3.7 酶解产物抗氧化活性的测定
ABTS阳离子自由基清除能力的测定参考文献[18];DPPH自由基清除能力的测定参考文献[19-20];还原能力的测定参考文献[21]。酶解产物的抗氧化能力以半数抑制浓度(IC50)表示,IC50指的是清除率为50%时或OD值达到0.5时,所需要样品的有效质量浓度,可根据不同质量浓度的清除率作曲线方程求出。同时设立VC为阳性对照。
1.4 数据统计与分析
2 结果与分析
2.1 鳜鱼发酵前后的细菌菌群α多样性分析

表1 发酵前后鳜鱼的细菌菌群α多样性分析Table 1 Bacterial α diversity indexes of fermented and fresh mandarin fish
由表1可知,未发酵鳜鱼和发酵后臭鳜鱼细菌菌群的覆盖率都达到99%以上,说明样本文库测序结果能反映样品真实情况,可用于样品细菌多样性分析。未发酵鳜鱼细菌菌群的Chao l指数平均值为27.13,Shannon指数平均值为1.36,ACE指数平均值为29.34;发酵后臭鳜鱼细菌菌群的Chao l指数平均值为68.93,Shannon指数平均值为1.67,ACE指数平均值为67.89。从Chao l指数和ACE指数可以看出,发酵后臭鳜鱼的细菌菌群丰度高于未发酵鳜鱼细菌菌群丰度。发酵后臭鳜鱼中细菌Shannon指数也明显高于未发酵鳜鱼,说明发酵后臭鳜鱼中细菌菌群多样性高于未发酵鳜鱼。因此,传统发酵臭鳜鱼中细菌菌群多样性更加丰富。
2.2 发酵前后鳜鱼的细菌群落结构差异性分析
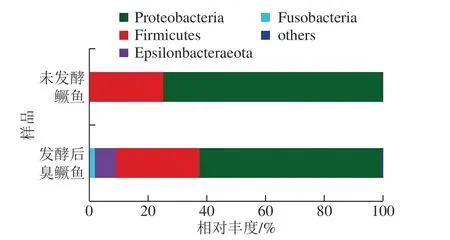
图1 门水平上发酵前后鳜鱼的细菌丰度分析Fig. 1 Relative abundance of bacteria in fermented and fresh mandarin fish at the phylum level
由图1 可知,在门水平上,发酵前鳜鱼总细菌群落主要由变形菌门(Proteobacteria)和厚壁菌门(Firmicutes)组成,相对丰富分别为74.55%、25.45%。发酵后臭鳜鱼的总细菌群落组成仍以变形菌门和厚壁菌门为主,占据菌群总量的90.34%,余下9.37%为Epsilonbacteraeota、梭杆菌门(Fusobacteria)。由此说明,徽州臭鳜鱼是通过鱼体自身携带的微生物在适宜温度条件下自然发酵形成的,变形菌门和厚壁菌门是臭鳜鱼发酵的主要优势细菌。
由图2 可知,在属水平上,发酵前后鳜鱼的微生物多样性与丰度有明显差异。发酵前鳜鱼共分离得到8 个属,相对丰富大于1%的菌群为假单胞菌属(Pseudomonas)、肉杆菌属(Carnobacterium)、沙雷氏菌属(Serratia)、漫游球菌属(Vagococcus)、环丝菌属(Brochothrix)、乳杆菌属(Lactobacillus)及冷杆菌属(Psychrobacter)。发酵前鳜鱼中假单胞菌属和肉杆菌属的优势较为明显,分别为总水平的69.30%和17.33%。发酵后臭鳜鱼共分离得到14 个属,与发酵前鳜鱼相比,发酵后臭鳜鱼的总细菌群落组成发生变化,主要由冷杆菌属、漫游球菌属、弓形杆菌属(Arcobacter)及Psychrilyobacter组成,相对丰度分别为59.96%、26.97%、7.21%及1.23%。冷杆菌属微生物在臭鳜鱼发酵中占有优势,对臭鳜鱼发酵起重要作用。已有报道表明,冷杆菌属是低温发酵食品的主要优势菌群,在冷鲜肉、海产品及牛奶中都十分常见[14,22]。同时,本研究从发酵后臭鳜鱼中检测出乳杆菌属,与罗靓芷[5]和Dai Zhiyuan[23]等传统培养方法的研究结果一致。乳酸菌能赋予发酵臭鳜鱼柔和的酸味,改善产品风味,促进发酵成熟。
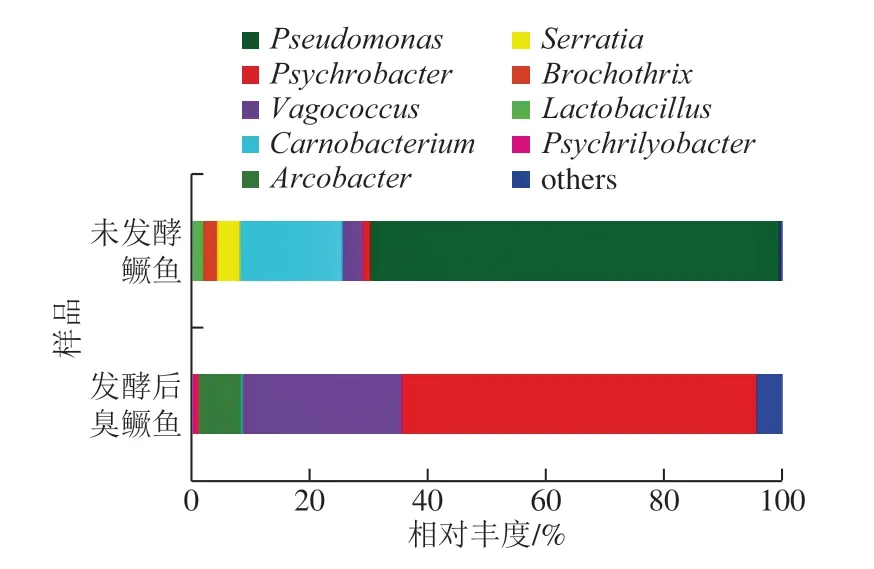
图2 发酵前后鳜鱼的属水平上细菌丰度分析Fig. 2 Relative abundance of bacteria in fermented and fresh mandarin fish at the genus level
2.3 发酵前后鳜鱼的理化性质分析
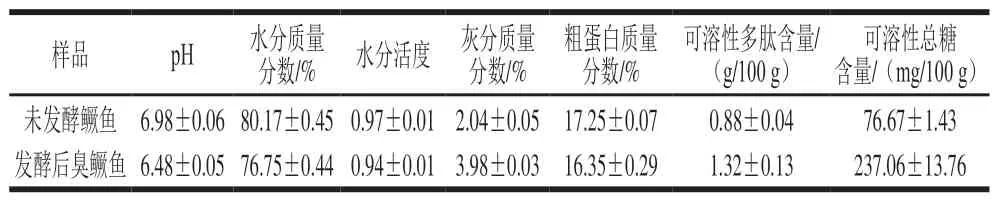
表2 发酵前后鳜鱼理化性质的比较Table 2 Comparison of physicochemical properties of fermented and fresh mandarin fish
由表2可知,相比于未发酵组,发酵后臭鳜鱼的pH值及水分含量、水分活度、粗蛋白含量略降低。臭鳜鱼的水分含量及水分活度下降,可能与发酵前期臭鳜鱼加入食盐腌制中,鱼肉脱水致使含水量降低以及与发酵后期pH值的下降引发了蛋白质变性,使其鱼肉持水能力降低等有关。发酵后臭鳜鱼的灰分、可溶性多肽、可溶性总糖含量较发酵前均明显提高,分别增加了95.10%、50.00%、209.20%。臭鳜鱼灰分含量的增加,与发酵过程中加入食盐腌制有关。
2.4 发酵前后鳜鱼的理化指标与细菌相关性分析
目前已有多篇文献报道了发酵食品微生物群落与其理化指标具有一定的相关性:如姜静[24]研究表明自然发酵豆酱的pH值与乳杆菌、假单胞菌、环丝菌等呈现极显著负相关,蛋白质和脂肪含量与乳杆菌、明串珠菌和环丝菌呈现极显著正相关;Broekaert等[25]研究发现细菌Psychrilyobacterspp.、Pseudoalteromonasspp.是褐虾在保藏过程中挥发性成分(硫化物、酮类、氨类、醇类等成分)产生的主要优势菌群;孙瑛等[26]研究表明海水鱼腌制过程细菌群落多样性和N-亚硝胺变化呈现一定相关性,嗜冷杆菌属为优势菌群。本研究对发酵前后鳜鱼的理化指标与细菌变化进行相关性分析,结果如表3所示。漫游球菌属与pH值、粗蛋白含量呈显著负相关(P<0.05),假单胞菌属、肉杆菌属、沙雷氏菌属和乳杆菌属与pH值呈显著正相关(P<0.05)。漫游球菌属与可溶性多肽、可溶性总糖含量呈显著正相关(P<0.05),相关性系数(r)分别为0.999和0.999,表明在发酵过程中漫游球菌属能合成蛋白酶、碳水化合物活性酶等,促进了蛋白质、碳水化合物的分解,致使发酵后粗蛋白含量下降,而可溶性多肽、可溶性总糖含量显著上升[27]。结果揭示了发酵前后鳜鱼的理化指标与细菌群落呈现显著相关性,而细菌群落代谢的复杂酶体系可能是引起发酵前后鳜鱼理化性质发生变化的主要原因。
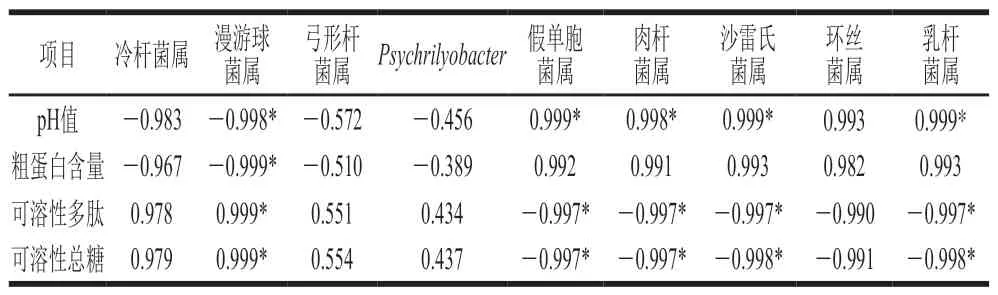
表3 理化指标与细菌相关性分析Table 3 Correlation analysis between physicochemical indexes and bacterial genera in stinky mandarin fish
2.5 发酵前后鳜鱼的质构特性分析

表4 发酵前后鳜鱼硬度、色泽的比较Table 4 Comparison of hardness and color parameters of fermented and fresh mandarin fish
由表4可知,未发酵鳜鱼的硬度值为(2.10±0.10)kg/cm2,发酵后臭鳜鱼的硬度值增加到(3.95±0.09)kg/cm2,提高了0.88 倍。L*值表示发酵前后鳜鱼鱼肉的明暗度,L*值越大表明颜色越白。与未发酵鳜鱼相比,发酵后臭鳜鱼的L*值减少,鱼肉呈现灰白色;发酵前后鳜鱼的b*值亦存在差异,发酵后臭鳜鱼的b*值显著增加,鱼肉颜色呈现淡黄色[28]。如图3所示,与未发酵鳜鱼相比,发酵后臭鳜鱼的组织结构形态发生了显著改变,外表面干燥,脱水严重,组织结构出现严重皱缩,卷曲现象更明显,鱼肉间形成了较为清晰的纹路。发酵前后鳜鱼微观结构扫描电镜观察结果与前期研究的水分含量、水分活度及硬度具有一定的吻合度[29],从微观结构上对发酵前后鳜鱼质构特性差异进行了进一步的验证。
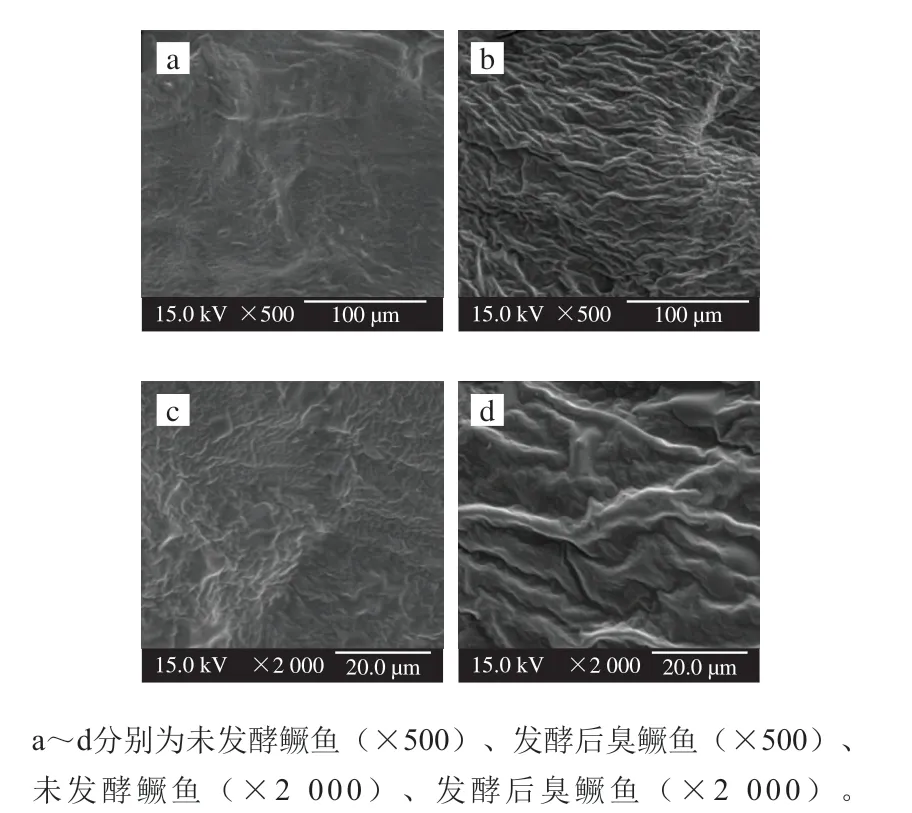
图3 发酵前后鳜鱼微观组织结构的比较Fig. 3 Comparison of microstructure of fermented and fresh mandarin fish
2.6 臭鳜鱼不同蛋白酶酶解产物的可溶性多肽含量分析
选用酸性蛋白酶、中性蛋白酶、碱性蛋白酶及木瓜蛋白酶分别在最适条件下对臭鳜鱼肉糜进行酶解,以可溶性多肽含量为评价指标,结果如表5所示。不同蛋白酶酶解液中可溶性多肽含量差异显著(P<0.05),其中碱性蛋白酶的酶解程度最高,可溶性多肽含量可达(4.97±0.18)g/100 g;未处理组中可溶性多肽含量最低,为(1.32±0.13)g/100 g,显著低于其他4 种蛋白酶处理组(P<0.05)。4 种蛋白酶酶解能力从大到小依次为碱性蛋白酶>中性蛋白酶>木瓜蛋白酶=酸性蛋白酶。包汭琪[12]和王翠[13]等研究表明,中性蛋白酶和木瓜蛋白酶能有效促进臭鳜鱼中蛋白质水解,游离氨基酸含量显著提升。
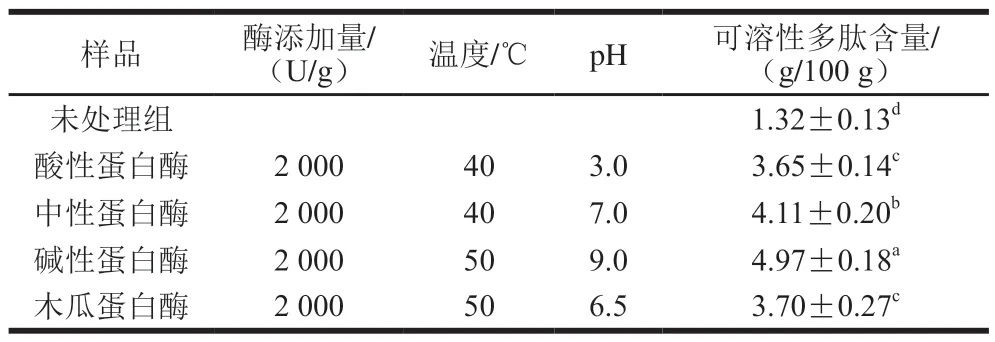
表5 臭鳜鱼的不同蛋白酶酶解产物可溶性多肽含量的比较Table 5 Comparison of soluble polypeptide contents in different protease hydrolysates of stinky mandarin fish
2.7 臭鳜鱼不同蛋白酶酶解产物对ABTS阳离子自由基清除作用
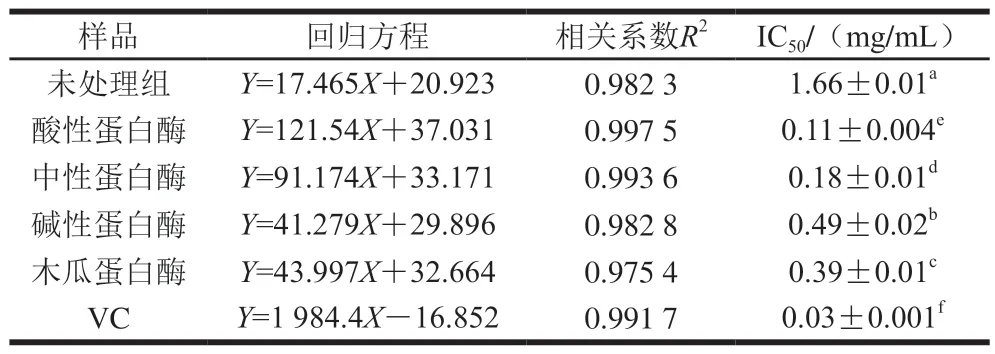
表6 臭鳜鱼酶解产物对ABTS阳离子自由基的清除作用Table 6 ABTS cation radical- scavenging ability of enzymatic hydrolysates of stinky mandarin fish
由表6可知,臭鳜鱼4 种蛋白酶酶解产物对ABTS阳离子自由基清除作用均表现出明显的量效关系,即随着酶解产物质量浓度的增加,ABTS阳离子自由基清除能力随之增强。酸性蛋白酶、中性蛋白酶、碱性蛋白酶及木瓜蛋白酶的酶解产物清除ABTS阳离子自由基能力较未处理组均显著提高(P<0.05),对ABTS阳离子自由基清除作用的IC50值分别为(0.11±0.004)、(0.18±0.01)、(0.49±0.02)、(0.39±0.01)mg/mL,而未处理组的IC50值为(1.66±0.01)mg/mL。臭鳜鱼酶解产物ABTS阳离子自由基清除能力的显著提高,可能与不同蛋白酶酶解产物中可溶性多肽含量的增加有关。有大量研究显示,多肽类物质能显著清除ABTS阳离子自由基,具有良好的抗氧化作用[30-32]。
2.8 臭鳜鱼不同蛋白酶酶解产物对DPPH自由基清除作用
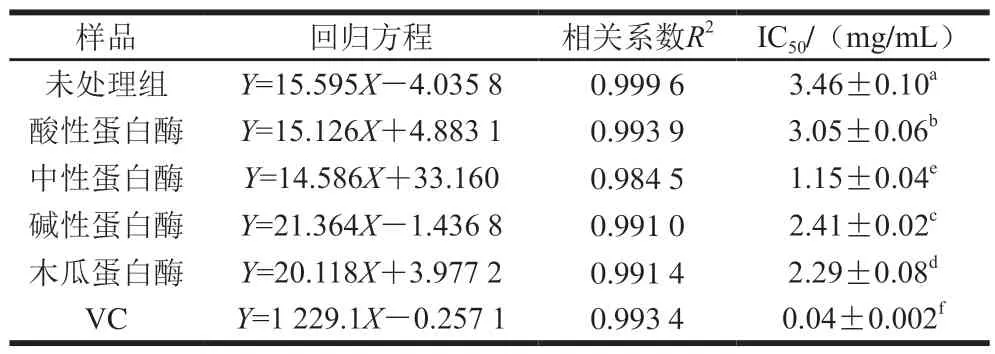
表7 臭鳜鱼酶解产物对DPPH自由基的清除作用Table 7 DPPH radical scavenging ability of enzymatic hydrolysates of stinky mandarin fish
由表7可知,臭鳜鱼的4 种蛋白酶酶解产物均呈现出显著的DPPH自由基清除作用,并呈现明显的量效关系。DPPH自由基清除能力从大到小依次为VC>中性蛋白酶>木瓜蛋白酶>碱性蛋白酶>酸性蛋白酶>未处理组,其DPPH自由基清除作用的IC50值分别为(0.04±0.002)、(1.15±0.04)、(2.29±0.08)、(2.41±0.02)、(3.05±0.06)、(3.46±0.10)mg/mL,说明4 种蛋白酶对臭鳜鱼鱼肉进行蛋白质酶解,可以显著提高其DPPH自由基清除作用(P<0.05)。
2.9 臭鳜鱼不同蛋白酶酶解产物对还原能力的影响
由表8可知,臭鳜鱼不同蛋白酶酶解产物均具有较好的还原能力,随着酶解产物质量浓度的增加,还原能力增强。与未发酵组相比,臭鳜鱼的4 种蛋白酶酶解产物的还原能力显著提高(P<0.05)。结果表明,4 种蛋白酶酶解产物还原能力的变化规律与对ABTS阳离子自由基清除能力一致,说明臭鳜鱼中可溶性多肽可能是其发挥抗氧化作用的主要活性物质。
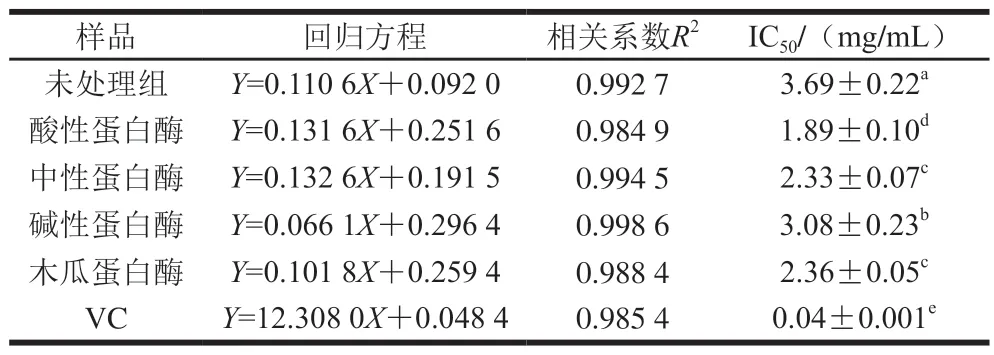
表8 臭鳜鱼酶解产物的还原能力Table 8 Reducing powers of enzymatic hydrolysates of stinky mandarin fish
3 结 论
本研究采用高通量测序技术分析发酵前后鳜鱼中菌群组成及变化情况,解析各类微生物的结构比例,明确优势菌群。结果表明,发酵前鳜鱼的主要优势菌为假单胞菌属、肉杆菌属、沙雷氏菌属、漫游球菌属及冷杆菌属;而发酵后臭鳜鱼的总细菌群落组成发生变化,主要由冷杆菌属、漫游球菌属、弓形杆菌属及Psychrilyobacter组成。由于氧气含量、温度、盐浓度、竞争等因素,发酵后臭鳜鱼的优势菌数量逐步增加变得显著,说明优势菌与臭鳜鱼的发酵品质有密切关系[22],其中冷杆菌属、漫游球菌属、乳杆菌属等为主要优势菌,在臭鳜鱼发酵过程中起重要作用。发酵后臭鳜鱼的营养成分、理化性质及质构特性等发酵品质发生了显著变化,其中灰分、可溶性多肽、可溶性总糖、硬度较发酵前均显著增加,而水分、水分活度、粗蛋白明显减少;发酵后臭鳜鱼鱼肉呈现灰白色,鱼肉组织结构出现严重皱缩,卷曲现象更明显,鱼肉间形成了较为清晰的纹路。相关性分析表明,菌群的种类和丰度对发酵后臭鳜鱼理化指标的影响较大,从而影响产品品质,其中对发酵后臭鳜鱼理化指标影响较大的细菌为漫游球菌属。
采用酸性蛋白酶、中性蛋白酶、碱性蛋白酶和木瓜蛋白酶对发酵后臭鳜鱼肉糜进行蛋白质酶解,并评价4 种不同酶解产物的抗氧化能力。结果发现,臭鳜鱼4 种蛋白酶酶解产物清除ABTS阳离子自由基、DPPH自由基能力及还原能力较未处理组显著提高,这可能与不同蛋白酶酶解产物中可溶性多肽含量的增加有关。本研究阐明了发酵前后鳜鱼微生物群落组成和品质特性的差异及相关性,揭示了发酵后臭鳜鱼蛋白酶解产物具有良好的抗氧化活性,研究结果为准确认识徽州臭鳜鱼的微生物群落结构组成及发酵品质提供数据参考,为臭鳜鱼的微生物与酶协同发酵及提高臭鳜鱼应用价值提供了可能性。
猜你喜欢
化工管理(2022年14期)2022-12-02
河南医学研究(2022年19期)2022-10-19
今日农业(2022年4期)2022-06-01
当代水产(2021年9期)2021-12-02
环境昆虫学报(2021年5期)2021-10-20
当代水产(2021年3期)2021-07-20
国际消化病杂志(2021年1期)2021-03-05
文化交流(2019年1期)2019-01-11
分析化学(2017年12期)2017-12-25
科学中国人(2015年16期)2015-02-28
