
干旱对马尾松幼苗光合作用及相关生理的影响
2021-01-16 07:17张金凤陈佩珍孙晓波胡兴峰季孔庶
中国农学通报 2021年1期
张金凤,陈佩珍,孙晓波,胡兴峰,季孔庶
(南京林业大学林木遗传与生物技术教育部重点实验室/南方现代林业协同创新中心,南京 210037)
0 引言
随全球气候变暖,干旱地区、半干旱地区的水分短缺问题日益凸显,干旱问题已严重影响国内干旱地区、半干旱地区的农林生产[1],使植物生长受限,达不到栽培的预期效果,影响其生态、经济及景观价值。通常情况下,植物通过自身的形态变化和生理反应来降低干旱伤害以适应环境[2-7]。比如超氧化物歧化酶、过氧化物酶作为植物的抗氧化性酶,对植物细胞起保护作用[8];丙二醛是衡量膜质过氧化的一个重要指标,能反映膜系统的受损伤程度[9-10];脯氨酸作为植物体内重要的渗透调节物质,具有较强的水合力,其积累可增加植物的抗旱或抗渗透胁迫能力[9,11];气孔导度反映气孔张开的程度,直接影响净光合速率和蒸腾速率[12]。干物质积累量和根冠比作为衡量植物生长的重要指标,反映了植物地上、地下的生长情况[13]。植物在受到干旱胁迫时,保护酶超氧化物歧化酶、过氧化物酶的活性明显提高[14],脯氨酸、丙二醛含量增加[15-16],干物质积累受到抑制[13],根冠比增大[17],净光合速率、气孔导度和蒸腾速率下降[18],从而减少干旱伤害。因此,探明干旱、半干旱地区植物的响应机制和适应性以及培育更多抗旱性植物具有重要的意义。
马尾松(Pinus massoniana)是国内南方主要乡土造林树种和改善环境绿化荒山先锋树种,具有速生、丰产、耐贫瘠等优良特性,在工业用材、林产品加工、森林资源发展及生态服务功能中具不可替代的作用[19]。马尾松产区分布广,年降雨量时空分布不均匀限制了马尾松人工林的大面积扩展。科研人员对不同种源[20]和家系[16]马尾松苗木对干旱的响应作过研究,为其耐旱种质的筛选与评价提供了一定的理论基础和参考依据,但在抗旱指标的选择上仍存在一定的局限性,各种生理响应的机理尚待深入研究。本研究以马尾松1年生实生盆栽幼苗为试验材料,研究不同程度控水胁迫下幼苗的生理伤害,探讨马尾松幼苗对不同土壤水分条件的适应能力,旨在为研究马尾松抗旱机制及抗旱良种选育提供理论依据。
1 材料与方法
1.1 试验时间、地点
马尾松苗于2017年在贵州省都匀市马尾松国家林木良种基地繁育,气候条件为当地正常条件。室内土培盆栽试验于2018年在南京林业大学林木遗传与生物技术教育部重点实验室温室进行。
1.2 试验材料
马尾松是来源于贵州省都匀市马尾松国家林木良种基地1年生马尾松半同胞家系幼苗,于2018年3月移栽至南京林业大学温室,共200盆,每盆1株。土壤取自南京林业大学后山马尾松林下黄壤(pH 5.0~5.5),每盆土壤量一致。
1.3 试验设计
盆栽缓苗3个月后随机选取健康长势一致的幼苗120盆,于6月20日在温室内采用称重法进行土壤含水量控制。水分处理分正常(CK)、轻度(T1)、中等(T2)、严重(T3)4个梯度,为田间持水量的(80±5)%、(65±5)%、(50±5)%和(35±5)%。每个处理10盆,3次重复。每天16 h光培养(光照强度35000 lx),8 h暗培养,室内温度控制在15~22℃,湿度约75%。干旱胁迫持续60天。
1.4 指标测定
1.4.1 生长指标 马尾松幼苗胁迫前0天和胁迫后60天时,每个梯度各选3株苗木,取整株苗木分地下和地上部分烘干称重得苗木干重,处理前后干重之差为苗木干物质积累量,苗木地下、地上部分干重比值为根冠比。苗高和地径每15天测量一次,10次重复,苗高采用直尺测定,地径采用电子游标卡尺测量。
1.4.2 光合指标 选择晴朗天气,在上午10:00左右采用CIRAS-3光合仪随机选择不同处理条件下的马尾松幼苗各3株进行光合测定,6月20日设定为胁迫0天,7月5日为第15天胁迫,7月20日为第30天胁迫,8月5日为第45天胁迫,8月20日为第60天胁迫,测定时运用缓冲瓶使得CO2浓度处于相对平稳状态,测定的主要指标包括净光合速率(net photosynthetic rate,Pn)、蒸腾速率 (transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)等,3次重复。
1.4.3 生理指标 在特定波长下,运用黄嘌呤氧化酶-氮蓝四唑(NBT)法检测超氧化物歧化酶(superoxide dismutase,SOD)活性,过氧化物酶(peroxide,POD)活性用愈创木酚法测定,用酸性茚三酮比色法测定脯氨酸(proline,Pro)的含量,脂类氧化损伤丙二醛(malonaldehyde,MDA)含量用硫代巴比妥酸法检测,所用试剂盒均购自南京建成生物公司并严格按照说明书操作。每个指标3次重复(每10株苗混合采样为1次重复),测定时间与光合指标测定时间一致。
1.5 数据分析
采用Excel和R语言ggcorrplot包对数据进行统计分析。
2 结果与分析
2.1 干旱胁迫对马尾松幼苗生长指标的影响
2.1.1 干旱胁迫对苗高、地径的影响 干旱胁迫下马尾松幼苗生长受到不同程度的抑制。由表1可以看出,干旱胁迫60天后T1与CK苗高、地径的生长均无显著差异(P>0.05),苗高分别增长了1.95、1.76 cm,增长率分别为5.32%、5.03%;地径分别增长0.45、0.41 mm。T2、T3生长缓慢,且与CK均有显著差异(P<0.05),苗高分别增长了0.74、0.53 cm;地径分别增长0.21、0.12 mm,T1的苗高和地径增长量约是T3的3.3倍,说明干旱胁迫对马尾松幼苗生长造成严重影响。
2.1.2 干旱胁迫对干物质积累和根冠比的影响 植物在逆境胁迫下,干物质积累反映了其对外界环境的适应能力。如表1所示,生物量的积累随着胁迫加剧而受到抑制,使马尾松幼苗生长减缓,60天后T3根生物量较CK降低69.49%,茎生物量较CK降低89.69%,说明根系对干旱胁迫的敏感程度略低于茎叶部分。T1干物质积累(0.74 g)与CK(0.86 g)差异性不显著(P>0.05);随干旱胁迫加剧,T2与T3干物质积累均受到抑制,较CK分别降低了53.49%、75.58%。水分充足条件下,地上部分生长旺盛,远大于地下部分的生长,根冠比较小;T1的苗高生长量、地径生长量、干物质积累量及根冠比均略高于CK,但差异性均不显著(P>0.05);T3苗高生长量、地径生长量及干物质积累比CK分别降低了69.89%、70.73%、75.58%,差异性均达到显著水平(P<0.05);T3根冠比为0.42,较CK提高30.95%,说明干旱胁迫能够引起了马尾松地上和根系生物量重新分配,根系生物量积累增多,根冠比增大。
2.2 干旱胁迫对马尾松幼苗光合特性的影响
2.2.1 干旱胁迫对马尾松幼苗气孔导度(Gs)的影响 气体交换参数是判断植物应对不同水分梯度的重要指标之一。通过图1可看出,在0~15天,马尾松幼苗的气孔导度随干旱胁迫程度加剧和胁迫时间延长而急剧下降,且差异性显著(P<0.05);处理45~60天,气孔导度变化相对稳定,可能是经过一段时间的干旱胁迫,苗木能短期适应干旱胁迫的表现;处理60天之后,与0天相比,CK、T1、T2、T3的气孔导度分别下降了5.08%、19.93%、56.12%、73.13%,其中T2和T3下降幅度高于CK和T1。
2.2.2 干旱胁迫对马尾松幼苗蒸腾速率(Tr)的影响 从图1可看出,干旱胁迫下马尾松幼苗蒸腾速率与CK相比呈下降趋势。CK的蒸腾速率基本保持不变,T1与CK之间差异性不显著(P>0.05)。处理15~60天,T2和T3蒸腾速率明显下降,与CK差异性均达到显著水平(P<0.05),说明马尾松幼苗受到干旱胁迫影响,以减少水分蒸发的方式来平衡体内渗透压,维持细胞的正常形态。
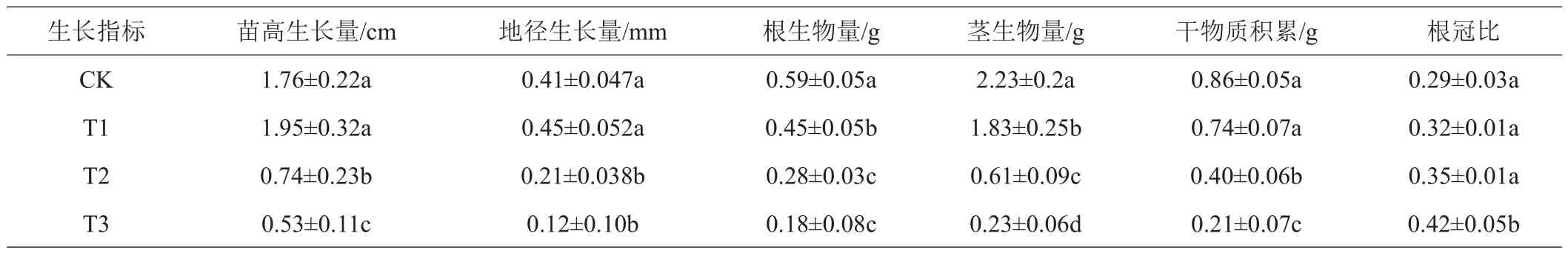
表1 干旱胁迫(60天)马尾松幼苗形态生长指标及生物量的变化

图1 干旱胁迫对马尾松幼苗光合速率及相关参数的影响
2.2.3 干旱胁迫对马尾松幼苗净光合速率(Pn)的影响从图1可看出,干旱胁迫后,马尾松幼苗净光合速率呈均匀下降趋势。整体来看,T1与CK相比下降不明显,T2下降趋势相对缓和,提前进入稳定期,而T3在胁迫0~45天净光合速率急剧下降,与CK呈显著性差异(P<0.05);45天后,T3的净光合速率趋于零水平,说明若长期处于重度干旱状态下,马尾松苗木生产力会下降,甚至出现死亡现象。
2.3 干旱胁迫对马尾松幼苗生理特性的影响
2.3.1 干旱胁迫对超氧化物歧化酶活性的影响 从图2A可以看出,随着干旱胁迫加剧,马尾松幼苗保护酶SOD的活性逐渐增强,随处理时间延长,SOD活性呈先上升后下降的趋势。处理15天时,T2和T3开始显著上升,30天时T2和T3均达峰值699.81、817.82 U/(g·FW),与CK及T1差异显著(P<0.05),比CK分别上升75.57%、104.01%;45天时SOD显著下降(P<0.05),但仍表现出较高的活性,60天时SOD活性依然高于胁迫0天时。因此,随着干旱胁迫时间的延长,马尾松幼苗通过提高SOD活性来清除植物体内产生的自由基,以适应干旱环境。
2.3.2 干旱胁迫对过氧化物酶活性的影响 过氧化物酶作为判断植物被伤害程度和抗性大小的指标。干旱胁迫下,马尾松幼苗POD活性随干旱胁迫程度加剧而增强(图2B),30天后缓慢下降,处理30天时各梯度的差异最大,随后差异逐渐缩小。随胁迫时间延长,各处理POD活性呈先升后降的趋势,与SOD活性变化趋势一致;处理30天时,T2与T3的POD活性均达峰值232.67、319.00 U/(g·min),与CK差异显著(P<0.05),比CK分别高98.30%、171.88%;随后POD活性缓慢下降,处理45天和60天时仍表现出较高酶活性,T2、T3间差异不显著(P>0.05),与CK和T1有显著差异(P<0.05)。
2.3.3 干旱胁迫对丙二醛的影响 丙二醛含量是植物细胞膜质过氧化程度的体现。随干旱胁迫加剧,马尾松幼苗MDA含量逐渐上升(图2C);0~15天MDA含量增速缓慢,差异不显著(P>0.05),15天后显著上升,30天时达至峰值,且T3、T2与CK、T1有显著差异(P<0.05),表明重度干旱胁迫下,细胞膜开始出现过氧化伤害。处理60天时,T3和CK、T1、T2之间仍有显著差异(P<0.05),说明植物体内活性氧的产生与清除的平衡状态遭到了破坏,脂质过氧化程度升高,引起过氧化产物MDA的产生和积累。

图2 干旱胁迫对马尾松幼苗生理特性的影响
2.3.4 干旱胁迫对脯氨酸的影响 作为植物细胞质内渗透调节物质,脯氨酸含量一定程度上反映了植物的抗逆性。由图2D可知,随干旱胁迫程度加剧,15天后马尾松幼苗Pro含量快速升高,且在同一水平处理条件下,Pro含量随胁迫时间延长呈上升趋势。处理15天后,T3、T2与CK、T1差异显著(P<0.05);处理30天后,T3与CK、T1、T2均存在显著性差异(P<0.05),T3比CK高258.86%;处理60天时,T1、T2、T3的Pro含量较45天时有所上升,其中T2和T3尤为明显,T3的Pro含量达峰值,说明马尾松幼苗在逆境中通过产生较多的Pro来维持细胞的渗透调节能力,以此来响应逆境条件下的胁迫,是马尾松实现自我保护的一种策略。
2.4 干旱胁迫下马尾松生长、生理指标与光合特性间的相关分析
干旱胁迫下马尾松生理指标与光合特性间的相关性分析如图3。整体来看,除根冠比外,生理指标与生长指标和光合指标呈均负相关,生长指标与光合指标呈正相关,且大多数呈现显著正相关(P<0.05);正相关的各指标间呈高度线性相关,r均在0.90以上;负相关的各指标相关系数r在-0.82以下,且生理指标POD与除根冠比外的其他生长指标和光合指标均呈显著负相关(P<0.05)。生长指标根冠比与生理指标MDA,Tr、茎生物量、Gs之间,Gs与根生物量均呈高度正相关(r=1);但Pro与Pn呈高度负相关(r=-1)。
3 结论
持续干旱胁迫下,马尾松幼苗通过抑制地上部分生长而促进地下根系生长,调整植物生长和生物量分配以抵御逆境;水分亏缺时,可引起马尾松幼苗叶片气孔开度减少及叶绿体类囊体膜结构受损,致使蒸腾作用和光合作用减弱;干旱胁迫初期,马尾松幼苗快速增加SOD、POD抗氧化性酶活性以保护过氧化伤害,中后期则增高渗透调节物质Pro维持细胞渗透压以降低膜质的损伤;基于生长、生理及光合等各项指标分析,发现生长与光合指标呈正相关,而生长与抗逆生理指标、根冠比呈负相关,光合与抗逆生理指标呈负相关,说明马尾松各部位间通过相互制约与协调应对干旱胁迫。
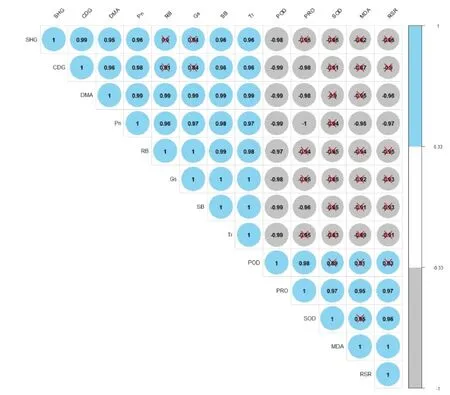
图3 干旱胁迫下马尾松生长生理指标与光合特性间的相关性分析
4 讨论
4.1 马尾松抗旱性与生长指标之间的关系
植物抗旱性是植物适应逆境的一种基本反应。本研究结果表明,干旱胁迫对马尾松幼苗生长指标产生直接影响,随干旱胁迫加剧,马尾松幼苗苗高生长量、地径生长量、根生物量和茎生物量出现了不同程度的下降,根生物量和茎生物量各组间存在显著差异(P<0.05),这与靳月等[21]的研究结果一致。有研究发现,在干旱胁迫下,植物常会通过增加地下部分生长、减少地上部分生长应对干旱造成的伤害[13]。本研究中,根生物量积累比茎生物量积累多,说明马尾松幼苗的根系对干旱胁迫的敏感程度可能略低于茎叶部分,也可能是干旱胁迫引起的生物量重新分配,生物量更多流向根部导致根冠比增大,或者根系深扎汲取土壤深处水分并积累更多的生物量。
4.2 马尾松抗旱性与光合指标之间的关系
气孔导度作为植物叶片与外界气体交换程度的指标,影响着植物的光合作用、呼吸作用和蒸腾作用[22]。马尾松幼苗在不同干旱胁迫下,Gs、Tr和Pn等逐渐下降,与宋学贵等[18]对竹柳光合特性的研究结果一致。这表明干旱一方面引起气孔开度减小甚至关闭,蒸腾作用减弱,影响CO2的吸收,光合速率下降,另一方面低水势造成植物叶绿体类囊体膜结构受损,光系统Ⅱ活力下降,电子传递和磷酸化作用减弱,光合速率下降[23]。
4.3 马尾松抗旱性与生理指标之间的关系
植物受到不同程度干旱胁迫时会通过自身调节机制平衡细胞内环境,使植物免受过分伤害。随干旱胁迫加剧,马尾松幼苗SOD和POD酶活性增强,MDA和Pro含量增加,随胁迫时间延长,SOD和POD酶活性均呈先升后降趋势,MDA和Pro含量升高,与黄华[24]、崔豫川[25]、宋丽梅[26]和 Nguyen-Queyrens等[27]在不同植物中的研究结果相似。这表明干旱胁迫初期会诱导增强SOD、POD酶活性保护马尾松免受伤害,随胁迫程度加剧,30天后酶活性开始降低,自由基大量产生,造成马尾松膜系统受到破坏,膜质过氧化程度加重,MDA含量上升;胁迫后期,Pro仍大量积累,以此增强渗透调节和细胞吸水能力来维持细胞水势,减轻胁迫造成的伤害,从而维持幼苗的生命活动。
4.4 马尾松抗旱性指标间的相关性
植物各部位间存在相互制约与协调的相关性。本研究中,马尾松幼苗茎生物量与根生物量呈高度正相关,其与靳月等[21]对闽南幼苗的研究结果一致,这表明植物地上和地下生长相互促进,地上生物量积累所需的水分、矿物质元素、细胞分裂素及植物碱等含氮化合物由根系提供,对根系的生长及生物量积累有促进作用,糖类及部分维生素只能由地上部分合成运输至根系。但在一些情况中,地下、地上部分的生长也会相互抑制,本研究中也有此结论,随着干旱胁迫的加剧,马尾松幼苗增加了根部物质积累,减少了地上部分的物质积累,使得根冠比值增大。蒸腾速率(Tr)、根生物量与气孔导度(Gs)之间均呈高度的正相关(r=1),这与严青青等[28]对海岛棉的研究结论一致。而脯氨酸(Pro)与净光合速率(Pn)呈很强的负相关(r=-1),与吕春华等[29]对小叶红叶石楠的研究结果一致,这表明马尾松通过调节细胞质基质与环境之间的渗透压,以减缓水分散失速率,维持较弱的光合作用。
本研究以马尾松幼苗为试材,从生长指标、光合特性和生理特性3个方面探究马尾松在受到干旱胁迫时的响应及其两两间的相关性,较全面地分析了不同程度控水胁迫下幼苗的生理伤害,并弥补了马尾松在抗旱指标选择上的不足。在试验过程中欠缺对幼苗生长状态的拍照记录及更多光合指标和生理指标的检测;其次在试验60天结束后,缺少复水处理及其生长、生理指标测定,这样可能利于更全面地了解马尾松幼苗对不同土壤水分条件的适应能力。在今后的研究工作中,在补充上述试验的同时,将研究马尾松抗旱的分子机制,从而更深入地揭示马尾松抗旱机理。
猜你喜欢
世界科学(2020年1期)2020-02-11
绿色科技(2019年5期)2019-11-29
中国生物医学工程学报(2019年5期)2019-07-16
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代农业科技(2017年16期)2017-09-22
Coco薇(2017年5期)2017-06-05
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
