
板栗花芽分化中内源激素的变化规律
2021-01-16 03:41:40赵志珩梁文汇廖健明黄晓露蓝振岐覃晓彬
广西林业科学 2020年4期
赵志珩,张 荣,严 蕾,梁文汇,廖健明,黄晓露,蓝振岐,覃晓彬
(1.广西壮族自治区林业科学研究院 广西特色经济培育与利用重点实验室,广西南宁 530002;2.遵义市凤凰山国家森林公园,贵州遵义 563000;3.西南林业大学,云南昆明 650224;4.河池市金城江区大山塘林场,广西河池 547000)
板栗(Castanea mollissima)为壳斗科(Fagaceae)栗属植物,广泛分布于我国26 个省,目前在山区已成为支柱产业,但其单位面积产量不高,制约了板栗产业的发展。根据国家林业和草原局发布的数据,2018年全国板栗产量为227 万t,仅为苹果产量的6.71%[1]。主要原因是板栗为雄单性、两性花或雌雄异花同株植物,雌花和雄花比例严重失调,雌、雄花比例高达1∶2 000 ~1∶3 500,雌、雄花序比例达1∶5[2],对坐果率、果实产量和质量产生了较大的影响。
通过切片观察,发现板栗花器官结构复杂,雄蕊原基存在退化的雌蕊器官,在雌蕊中也观察到了雄蕊。可知,板栗花芽分化具有可塑性,在一定条件下可以转化[3-5]。板栗在花芽分化的过程中,顶芽内的花序原基在休眠期前完成,为冬前雏梢分化期,一般在10月中旬左右;休眠期后,在前者花序原基上部节位,冬前形成的腋芽生长锥原基发育形成冬后花序原基,冬季休眠、芽体萌发或两者共同作用可能会让冬后花序获得发育成雌花序的能力[6]。据此,推测板栗花序性别分化调控的关键是冬后花序原基的分化,这类花序具有转变成雌花序的可能,且形成雌花序的过程具有可调控性[7-10]。本研究在前期解剖观察花芽内部形态特征的基础上,着重研究内源激素在板栗花芽分化期含量的变化,揭示板栗花芽分化的生理特性,对人工调控花芽分化以及提高产量具有重要的指导意义。
1 材料与方法
1.1 样株选择
试验材料为“易门一号”、“易门二号”和“云良”3 个云南本土的主栽品种。2014年高接,2016年开始结果,平均树高约4 m,种植密度为4 m×3 m。选择生长正常、树势和立地条件基本一致的植株取样。
在云南昆明安宁市板栗基地采集3个品种板栗的花芽,随机选取样株树冠外围不同方向的当年生枝条采集花芽,2017年7月10日— 8月25日每15天 采1 次,8月25日— 12月25日 每30 天 采1 次,2018年3月12日— 5月8日 每10 天 采1 次。每 次采集雄花枝和结果枝花芽各30 个,用锡箔纸包好,置于液氮中带回实验室,放入-80 ℃冰箱备用。
1.2 试验方法
用试剂盒法测定板栗混合花芽和不混合花芽中内源激素生长素IAA 、赤霉素GA3、脱落酸ABA和玉米素ZT的含量,试剂盒采用的是双抗体夹心法(ELISA)[11]。
1.3 数据处理
采用Oringe 85软件制图。
2 结果与分析
2.1 IAA含量变化
IAA 含量在完全混合花芽分化发育过程中呈M + 倒V 字形变化,在不完全混合花芽分化发育过程中呈N 字形变化(图1)。3 个品种的IAA 含量变化一致。
冬前分化期,完全混合花芽中的IAA 含量在7月10日— 8月25日呈上升趋势,8月25日达到第一个峰值,之后下降又上升,12月25日(休眠期)达到第二个峰值。说明在分化过程和休眠期,IAA 的作用有所不同,初期IAA 含量升高可促进花芽分化,休眠期IAA 含量升高可促进休眠。IAA 含量在3月中下旬(冬后雄花簇分化期和两性花序原基分化初期)达到谷值,之后稍回升,此时正处于两性花序原基向雌花簇分化的关键时期,说明IAA 含量的升高可能有利于雌花簇的分化。4月17日— 5月8日(花器官分化期),IAA 含量不断下降,说明低浓度的IAA 对完全混合花芽形成雌花有促进作用。
不完全混合花芽中的IAA含量在7月10日—9月25日(冬前分化初期至分化盛期)一直上升,至分化盛期达到峰值。3月12日后(冬后花芽分化期),IAA 含量呈上升趋势,说明高水平IAA 有利于雄花簇和雄花的分化。

图1 IAA含量变化Fig.1 Changes of IAA content
2.2 GA3含量变化
GA3含量在完全混合花芽分化发育过程中,呈N + V 字形变化,在不完全混合花芽分化发育过程中呈W+V字形变化(图2)。
3个品种完全混合花芽中的GA3含量变化一致,均在7月10日—8月10日(冬前分化初期)上升,之后下降,12月25日达到谷值。休眠期后,GA3含量随着分化进程的发展迅速增加,在3月下旬达到第2个峰值。说明GA3对花芽分化的影响较大,在冬前和冬后的分化初期,GA3含量的增加对花芽的启动十分有利。
不完全混合花芽中的GA3含量在7月10日相对较高,说明不完全混合花芽的雄花序原基分化一开始就需要大量GA3。3 个品种板栗花芽的GA3含量峰值和谷值出现的时间不一致,但在12月25日,GA3含量均较低,3月上旬(冬后雄花簇分化期)均较高。可以看出,在冬前和冬后的花芽开始分化时,需要较高水平的GA3参与。
完全混合花芽和不完全混合花芽中的GA3含量均在4月17—4月25日(雌花分化期)上升,说明两性花序的分化均需要较高水平的GA3参与。
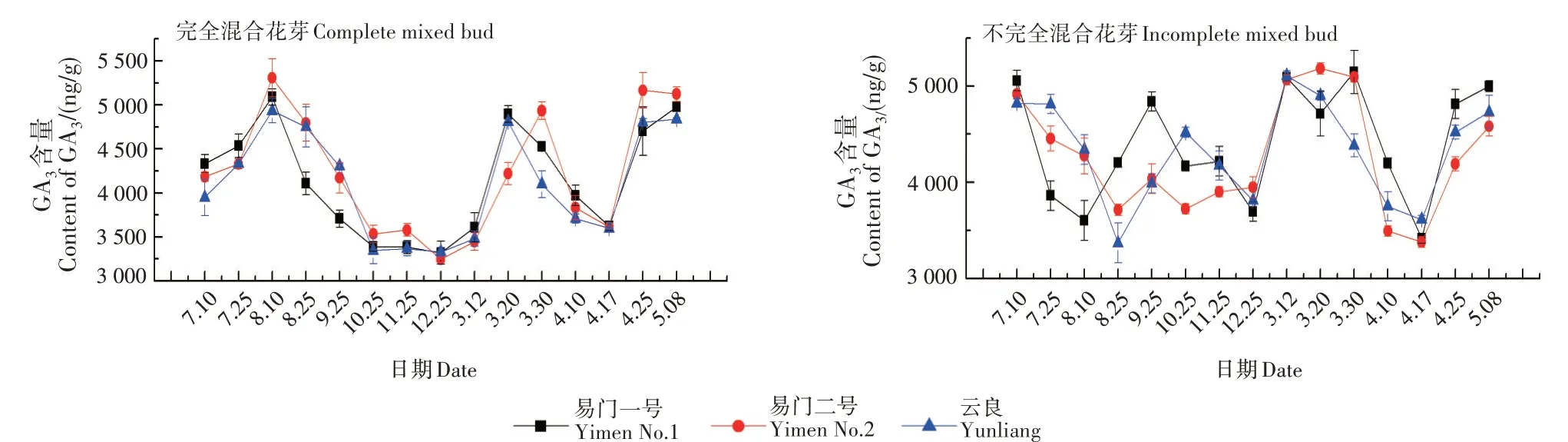
图2 GA3含量变化Fig.2 Changes of GA3 content
2.3 ABA含量变化
ABA 含量在完全混合花芽分化发育过程中呈M + 倒V 字形变化,在不完全混合花芽分化发育过程中呈V+L字形变化(图3)。
3 个品种完全混合花芽中的ABA 含量变化一致。7月10日—9月25日ABA含量先上升后下降,8月10日达到第一个峰值。9月25日—12月25日(冬前分化盛期至休眠期),ABA 含量持续上升,说明ABA 含量的升高有利于植物从分化期进入休眠期。3月12日— 20日(冬后花芽分化初期),ABA含量下降,出现第二个谷值。3月20日— 4月17日,ABA 含量呈上升趋势,此时正处于两性花序原基分化期,可以看出,ABA 含量的升高有利于完全混合花芽两性花序原基的分化。
在不完全混合花芽冬前分化过程中,ABA 含量表现为先下降后上升的趋势,12月25日达到峰值。之后ABA 含量迅速下降,3月12日后趋于稳定。可以看出,冬后较低水平的ABA 有利于冬后雏梢解除休眠及花芽分化。
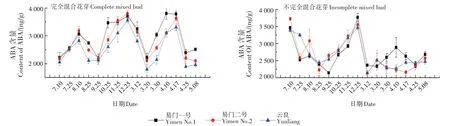
图3 ABA含量变化Fig.3 Changes of ABA content
2.4 ZT含量的变化
ZT 含量在完全混合花芽分化发育过程中呈M+ 倒V 字形变化,在不完全混合花芽分化发育过程中呈W+倒N 字形变化(图4)。3 个品种的ZT 含量变化一致。
完全混合花芽中的ZT 含量在7月10日— 11月25日(冬前分化期)呈上升趋势,11月25日(冬前分化末期)达到峰值,之后迅速下降,12月25日达到谷值。3月12日,ZT 含量处于较高水平,说明冬后两性花序原基以及雄花簇的分化,需要积累较多的ZT,ZT 含量高有利于打破休眠,刺激花芽分化。之后,ZT 含量逐渐下降,3月30日后(两性花序原基分化期)再次上升。说明完全混合花芽的分化在一定程度上都需要大量的ZT。
不完全混合花芽中的ZT 含量冬前和冬后的变化趋势一致,均呈降低-升高-降低的变化趋势。说明冬前、冬后花芽分化前和分化盛期都需要大量的ZT。

图4 ZT含量变化Fig.4 Changes of ZT content
3 结论与讨论
IAA 作为第一个被发现的植物激素在植物成花方面,它的作用一直存在争议。程华等[12]在对不同品种板栗的IAA 进行测定时发现,IAA 含量升高有利于花芽由生理分化向形态分化转变。结合课题组前期的切片试验,发现在完全混合花芽和不完全混合花芽的冬前(7月10日— 8月25日)和冬后花芽分化期(3月12日后),IAA 含量均由低到高,说明高水平IAA可能促进花芽分化;冬后,不完全混合花芽中的IAA 含量持续上升,完全混合花芽中的IAA含量在冬后的雌花簇分化期和花器官分化期下降,说明高水平IAA 可能促进雄花分化,而低水平IAA可能有利于雌花分化。雷新涛等[13]也认为较高水平的IAA 易形成雄花;对板栗进行生长素类的生长调节剂外施和内源生长素测定的结果也表明,低浓度IAA 有利于板栗雌花芽分化,高浓度IAA 抑制雌花芽分化[14-15]。本研究对板栗花芽分化不同时期的IAA 含量进行测定,更加直观地反应出花芽分化不同阶段IAA 的动态变化,为进一步外施激素提供更全面的理论支持。
在果树的花芽分化研究中,普遍认为GA 在花芽分化过程中有重要作用。低浓度GA 能促使板栗雄花芽分化[16]。本研究表明,完全混合花芽在进入雄花序原基分化前(7月10日前),对GA3含量需求不大,分化初期至分化盛期前(7月10日— 8月10日),对GA3需求不断增加,冬后也有相同的变化趋势;郭成圆等[17]的研究也发现,在板栗幼芽分化过程中,GA3含量的峰值出现在冬前花序原基分化期(10月中旬)和冬后花序原基分化期(次年2月中旬),与本研究结果一致,由于本试验样地在云南,具体的花序分化时间与郭成圆等[17]的研究不同。在花器官形成期间(4月17日— 4月25日)GA3含量持续上升,与季志平等[18]研究一致。不完全混合花芽在进入冬前和冬后分化时(7月10日前和3月12日前),均需要较高水平的GA3,此时可以在外部施用GA 类植物激素抑制剂,减少GA3的合成,从而抑制不完全混合芽雄花序的生成。
李从玉等[16]认为雄花芽分化与较高的ABA 含量有关。有些研究者发现ABA 可能对开花有双重影响,一方面,它能与GA3拮抗,使枝条停止生长,促进细胞分裂素、淀粉和糖的积累从而促进开花;另一方面又可诱导休眠,使植物进入休眠期,生长进程减慢,不能成花[19]。樊卫国等[20]对刺梨(Rosa roxburghii)的花芽分化进行研究,发现低水平ABA 更有利于刺梨花芽的分化。本研究发现板栗花芽在休眠期(12月25日)ABA的含量较高;完全混合花芽在冬前(7月10日)和冬后(3月12日)开始分化时的ABA 含量以及不完全混合花芽在冬后开始分化时(3月12日)的ABA 含量均较低,说明高水平ABA 促进休眠,低水平ABA 有利于花芽分化的启动。郭成圆等[17]研究发现板栗花芽中ABA 的含量在冬季花序原基分化期(10月中旬)最高,之后缓慢下降,1月中旬下降至稳定的低水平,并持续到花簇原基分化期(翌年3月中旬)。本研究结果也显示冬后花芽分化时,不完全混合芽中的ABA 含量持续稳定在较低水平。
ZT 是细胞分裂素的一种,具有促进果树花芽分化、细胞分裂和打破休眠的作用,其含量的高低与植物体内细胞分裂及代谢活动的强度有关。黄迪辉等[21]研究显示,暗柳橙(Citrus sinensis)在生理分化转向形态分化期间,对其进行去叶摘花试验,以减少二氢玉米素的上升,花芽数量也明显减少。李秉真等[22]发现在苹果梨(Pyrus pyrifoliacv.pingguoli)的花芽生理分化期(花芽诱导期),ZT 有利于花芽分化,能促进花序原基的形成和发育。本研究中,ZT含量在完全混合花芽发育过程中出现3 个峰值,分别为冬前分化末期(11月25日)、冬后分化期(3月12日)及两性花序原基分化期(4月25日);在不完全混合花芽发育过程中,ZT 含量有4 个峰值,分别是冬前花芽分化期(7月10日和8月25日)和冬后分化期(3月12日和4月10日)。冬后完全混合花芽和不完全混合花芽ZT 含量的变化趋势一致。完全混合花芽在冬前分化盛期到分化末期、冬后开始分化时和花器官分化期均需要大量的ZT,说明ZT 对细胞分裂,快速生长和分化启动均有重要作用。在生产过程中,可考虑在花芽分化开始前施用ZT,多形成花芽,保证冬后花芽数量,取得增产效果。
对板栗花芽分化影响的两个关键时期为冬前花序原基分化期(7月10日—11月25日)和冬后花序原基分化期(3月20日—4月17日)。冬前,高水平IAA 和GA3有助于花芽分化;冬后,低水平IAA 有助于雌花序原基分化,高水平GA3有助于雄花序原基的形成。ZT 在完全混合花芽和不完全混合花芽中表现一致性,在花芽发育的关键时期,高水平ZT 能同时促进雌花和雄花的形成。在生产中,冬前花芽分化期应施用ZT,以增加花芽数量;冬后花芽分化期应前施用GA3抑制剂,抑制不完全混合花芽发育,促进雌花分化,以达到“促雌抑雄”的目的。
猜你喜欢
金沙江文艺(2022年4期)2022-04-26 14:14:20
蚕桑通报(2022年3期)2022-04-10 03:16:00
落叶果树(2020年3期)2020-06-17 05:11:28
河北果树(2020年1期)2020-02-09 12:31:08
计算机应用(2018年10期)2018-11-22 09:37:54
华南师范大学学报(自然科学版)(2017年4期)2017-09-11 09:16:23
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19 07:41:15
广东农业科学(2016年3期)2016-07-27 02:07:13
天然产物研究与开发(2016年6期)2016-06-05 10:29:27
湖北农业科学(2014年6期)2014-07-02 11:16:14
