
永定河干涸段(卢梁段)河流廊道植物群落数量分类及多样性特征
2021-01-15 10:18刘志成杨瑞莹张任菲许晓明
生态学报 2020年24期
刘 璐,刘志成,杨瑞莹,张任菲,李 豪,许晓明
北京林业大学园林学院, 北京 100083
植物群落是指处于同一环境、具有不同于周边植被的相对统一的组成及结构的植被[1],其植被分类和物种多样性特征是群落生态学研究的重要内容[2]。在植被分类方法中,数量分类是最主要的、应用最为广泛的技术[3],对某一区域的植被进行数量分类,能够更准确地了解该区域植物群落结构特征[4]。植物群落数量分类方法众多,常用的有聚类分析、双向指示种分析法(TWINSPAN)、多元回归树(MRT)等[4- 6],其中, TWINSPAN是20世纪80年代起植被研究中使用最多的方法[7]。物种多样性是生物多样性在物种水平上的表现形式[8],被认为是生物多样性的核心[9- 11],使用一定空间范围物种数量和分布特征来衡量[7],能直接或间接地体现植物群落的生态结构[12, 13]。目前国内外对物种多样性的研究大多集中在物种多样性发生和维持的机制、物种多样性变化的影响因子[8]、生物多样性格局的自然梯度[14]等方面。
永定河是海河北系最大的河流[15]、“六河五湖”中的重要河流之一,是京津冀区域重要的水源涵养区、生态屏障和生态廊道,然而,长期以来永定河流域的生态环境十分脆弱:1954年官厅水库建成后,三家店以下河道逐渐干涸、砂质河床裸露,80年代起下游开始出现断流现象,生态环境逐渐恶化。近些年,为遏制永定河流域生态环境的恶化,相关部门采取了建设京冀水源保护林等一系列措施,对永定河进行生态修复和治理[16],并于2010年正式启动了永定河(北京段)生态治理工作,先期完成永定河“五湖一线”的治理任务,取得了一定的生态效益[17],但中下游的生态问题仍然十分突出。
2016年底,国家发改委等部门印发了《永定河综合治理与生态修复总体方案》,要求落实永定河河道综合整治、河道防护林建设、水源涵养林建设等治理措施,于2025年基本建成永定河绿色生态河流廊道,其中,中下游干涸河段为重点治理河段。由于以往对于永定河进行的植物调查及研究相对较少,并且主要集中在植被历史变迁、构成规律、区系特征等方面[18-20],对河流廊道植被分类和多样性特征的研究较少,要为永定河干涸河段重塑河滩地,进行防护林、水源涵养林的物种选择及位置确定等提供数据支持科学的理论依据,就需要对区域内植被现状进行详细的调查与分析,研究其物种构成特征、群落植被分类和多样性特征等。基于此,本研究采用TWINSPAN分类对永定河河流廊道植物群落进行数量分类,对不同层次间的物种多样性进行比较,并尝试借助回归分析研究植物群落物种多样性在垂直(纵向)及水平(横向)梯度上的空间分布格局,以期为永定河的生态修复的植被规划和工程实施提供一定的数据支持和理论依据。
1 研究区概况
永定河发源于内蒙古高原的南缘和山西高原的北部,全长747 km[17],其中北京段按河道特征和防汛特点分为官厅山峡段、平原城市段和平原郊野段。官厅山峡段从幽州入境至三家店拦河闸、平原城市段从三家店拦河闸至南六环路、平原郊野段从南六环路至梁各庄[21]。
研究选择基本全年干涸的卢沟桥至梁各庄段(以下简称干涸段),范围为东经116°12′41″—116°27′05″、北纬39°50′55″—39°26′12″,位于北京市丰台、大兴、房山区和河北省保定、廊坊市内。气候属于暖温带半湿润大陆季风气候,四季分明,年平均气温为11.6℃、年平均降水量556 mm,降水时间主要集中在7—8月份。研究区域内海拔由西北向东南逐渐降低,但海拔变化小;土壤沙化严重,以细粉砂为主。由于该段两岸均靠堤防约束,将河流廊道的范围界定为左右两堤之内。
现状河流廊道内水域空间消失,分布有大量草滩、林地、滥采砂石形成的裸露砂石坑及部分农田,另有多处高尔夫球场、农业设施、建筑物等,环境杂乱。由于北京市为治理永定河风沙带自1990年起开始在平原区植树造林[16],2012年又进行了百万亩造林工程,于永定河沿线新增造林约3000 hm2,因此河流廊道内现状植被以人工林为主、自然林较少,群落构成较为单一。
2 调查及研究方法
2.1 调查方法
调查工作于2019年7月进行,在研究区域内每间隔约5 km设置垂直于河堤的横断面为一个样带,每个样带采用“样方法”,从两堤起向内每间隔约100 m选择1个样方(如图1),样方大小为10 m×10 m,进行乔木层的测量,样方一角为5 m×5 m的灌木层小样方,四角和中心点共5个1 m×1 m的草本层小样方[22]。对样方内的植物进行详细调查,记录物种的种类、数量、高度、冠幅、盖度等,同时记录样方距堤距离、海拔等信息。实际调查中,共选取了12个样带,124个样方。
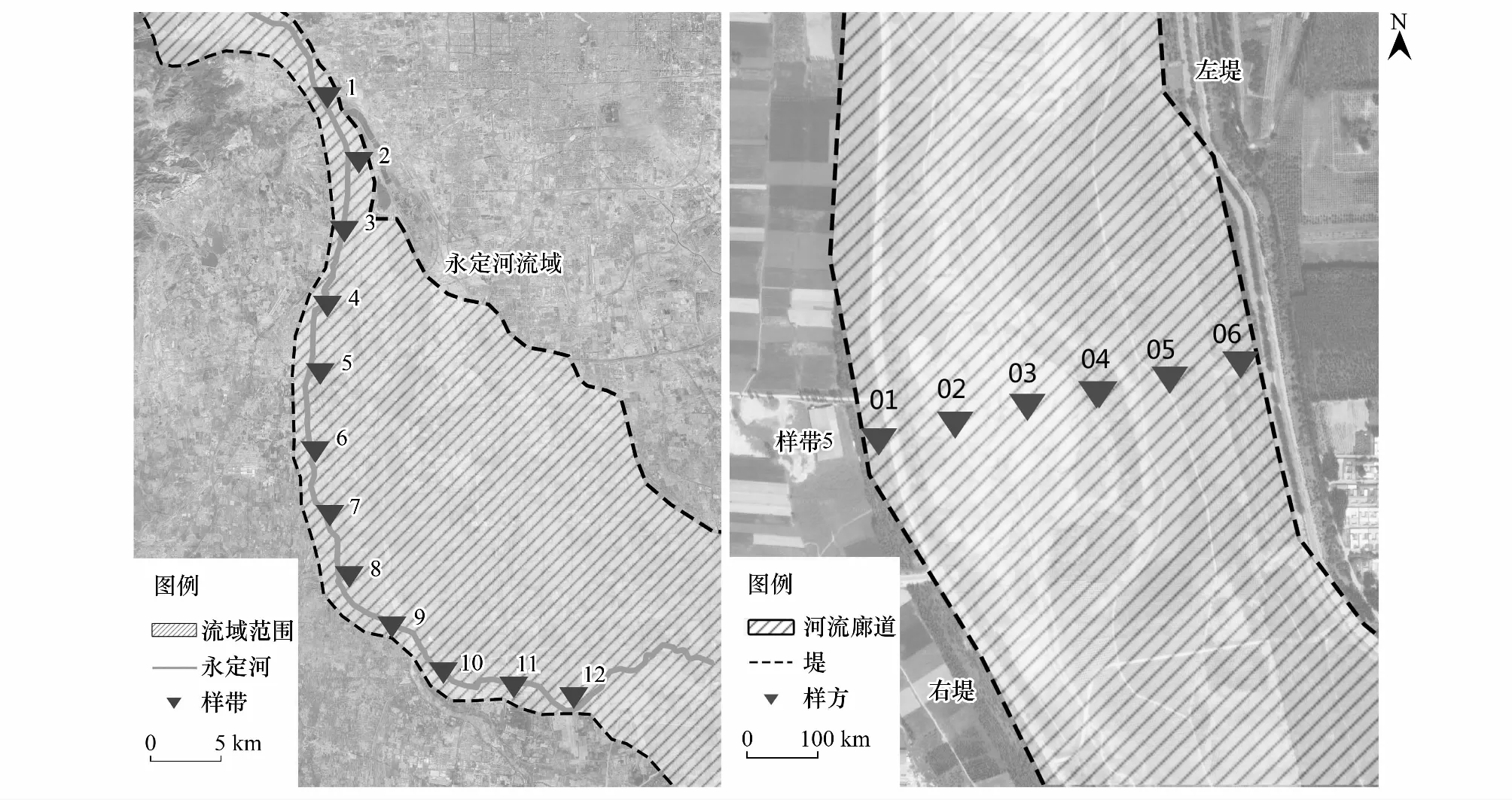
图1 样带及样方选取Fig.1 Selection of sample strips and plots
2.2 数据处理
2.2.1物种频度及重要值计算
(1)各物种频度计算公式为:
F=n/N×100%
式中,n为某一物种在调查中出现的样方数量和,N为全体样方的总数量。
(2)采用重要值(important value, IV)作为评价各物种在群落中的重要性指标[22],各物种重要值计算公式为:
乔木重要值=(相对密度+相对优势度+相对频度)/3
灌木重要值=(相对密度+相对盖度+相对频度)/3
草本重要值=(相对高度+相对盖度+相对频度)/3
2.2.2TWINSPAN分类
以124个植物群落各物种的重要值为基础数据,建立物种-样方矩阵,利用TWINSPAN分类方法进行群落数量分类。对数据进行CR/RA排序,同时得到样方和种类第一排序轴,分别用于样方分类和种类分类,获得双向分类矩阵[7]。TWINSPAN分析由PC-ORD Version 5完成,分类结果用树状图表示。
2.2.3群落的物种多样性计算及回归分析
群落的物种多样性采用Patrick丰富度指数(R)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(E)3个测定指标[7],对群落的物种多样性进行分析。计算公式如下:
(1)Patrick丰富度指数:
R=S
(2)Shannon-Wiener多样性指数:
(3)Pielou均匀度指数:
E=H/ln(S)
式中,S为样方内物种总数。Pi为第i个物种的相对重要值。
此外,运用Origin 8.0软件对样方的物种多样性数据与垂直、水平梯度的空间数据进行回归拟合与散点图绘制。
3 结果及分析
3.1 群落的物种构成
调查样方中共测得植物119种,隶属40科、97属。其中,乔木29种、灌木9种、草本81种;根据《北京植物志》,共有本地植物100种(84.03%)、国内引种植物15种(12.60%)、国外引种植物4种(0.03%)。从各物种的科属来看,研究区域的河流廊道植物主要分布于禾本科(Gramineae)(18种,占15.13%)、菊科(Compositae)(16种,占13.45%)、豆科(Leguminosae)(11种,占9.24%)和杨柳科(Salicaceae)(6种,占5.04%);含3- 5个物种的共有8科,分别为旋花科、藜科、萝藦科、桑科、百合科、柏科、大戟科和苋科。统计结果表明,永定河干涸段河流廊道灌木物种较少,草本植物集中的禾本科和菊科有明显突出的地位。
通过对各物种的频度进行统计,得到乔木物种频度大于10%的有刺槐(Robiniapseudoacacia)(0.12)、毛白杨(Populustomentosa)(0.12)和旱柳(Salixmatsudana)(0.11);草本频度大于10%的有狗尾草(Setariaviridis)(0.757)、猪毛菜(Salsolacollina)(0.29)、尖头叶藜(Chenopodiumacuminatum)(0.27)、蒺藜(Tribulusterrester)(0.26)、马唐(Digitariasanguinalis)(0.16)、灰菜(Chenopodiumalbum)(0.15)、地梢瓜(Cynanchumthesiodes)(0.14)和虎尾草(Chlorisvirgata)(0.10);灌木频度均小于3%。其中,狗尾草的频度远大于其他物种,在分布上呈绝对优势。
3.2 群落的TWINSPAN分类
对永定河断流区河流廊道的124个样方进行TWINSPAN分类,去除频度小于5%的偶见种,共计20个物种。根据各物种重要值设定分割值(cut level)分别为0.2、0.4、0.6、0.8、1,最终采用TWINSPAN第五层次水平的划分结果,分类结果见图2。

图2 124个样方TWINSPAN分类树状图Fig.2 TWINSPAN classification tree of 124 plots N表示样方总数; 图下方矩形框内的数字为124个样方的编号;横线上为各分类水平正负指示种的拉丁名缩写:SV:狗尾草Setaria viridis;TT:蒺藜Tribulus terrester;CAc:尖头叶藜Chenopodium acuminatum;UP:榆树Ulmus pumila;SM:旱柳Salix matsudana;SC:猪毛菜Salsola collina;CV:虎尾草Chloris virgata;RP:刺槐Robinia pseudoacacia;PTo:毛白杨Populus tomentosa;PC:加杨Populus × canadensis;DS:马唐Digitaria sanguinalis;PH:河北杨Populus hopeiensis;EI:牛筋草Eleusine indica;CT:地梢瓜Cynanchum thesioides;Cal:灰菜Chenopodium album
根据TWINSPAN数量分类结果,结合调查的实际结果和群落特征,按照《中国植被》的命名原则[23]将调查样方植被的主要群落类型进行了分类,共分为以下9个群丛类型:
Ⅰ.旱柳-狗尾草群丛(Ass.Salixmatsudana-Setariaviridis)。乔木层优势种为旱柳,少量伴生有加杨(Populus×canadensis)及国槐(Sophorajaponica),郁闭度为0.7—0.9;草本层优势种为狗尾草,平均盖度为0.1—0.9,次优种为猪毛菜、尖头叶藜,伴生种有蒺藜、马唐、牛筋草(Eleusineindica)等。该类群丛分布范围较广,调查区域内河流廊道各垂直梯度及水平梯度均有分布。
Ⅱ.河北杨-狗尾草群丛(Ass.Populushopeiensis-Setariaviridis)。乔木层优势种为河北杨,少量伴生有刺槐、油松(Pinustabuliformis),郁闭度为0.5—0.9;草本层优势种为狗尾草,平均盖度为0.1—0.8,主要伴生种为猪毛菜、尖头叶藜、灰菜、虎尾草及牛筋草。该类群丛大多分布在靠近河堤的位置。
Ⅲ.狗尾草+猪毛菜+尖头叶藜群丛(Ass.Setariaviridis+Salsolacollina+Chenopodiumacuminatum)。优势种为狗尾草、猪毛菜和尖头叶藜,盖度变化较大,次优种为蒺藜,伴生种为灰菜、虎尾草。该类群丛分布范围广泛且数量多,调查区域内几乎各样带均有分布。
Ⅳ.加杨-狗尾草+蒺藜群丛(Ass.Populus×canadensis-Setariaviridis+Tribulusterrester)。乔木层优势种为加杨,郁闭度为0.6—0.9,伴生种为毛白杨;草本层优势种为狗尾草和蒺藜,盖度分别为0.2—0.9和0.1—0.5,次优种为猪毛菜和虎尾草,另外伴生种有少量灰菜、牛枝子(Lespedezapotaninii)、尖头叶藜等。该类群丛主要分布在左右堤两侧,多作为防护林出现。
Ⅴ.毛白杨-狗尾草群丛(Ass.Populustomentosa-Setariaviridis)。乔木层优势种为毛白杨,郁闭度为0.5—0.9;草本层优势种为狗尾草,平均盖度为0.1—0.7,次优种为灰菜,另有少量猪毛菜、蒺藜。该类群丛基本分布在断流区上段(样带1—4)、北京区段内。
Ⅵ.蒺藜+狗尾草+葎草群丛(Ass.Tribulusterrester+Setariaviridis+Humulusscandens)。优势种为蒺藜、狗尾草和葎草,盖度变化较大,伴生种为马唐和地梢瓜。该类群丛基本分布在断流区中段至下段,距堤距离集中在100—300 m以内。
Ⅶ.刺槐-狗尾草群丛(Ass.Robiniapseudoacacia-Setariaviridis)。乔木层优势种为刺槐,郁闭度为0.4—0.9;草本层优势种为狗尾草,平均盖度为0.1—0.6,次优种为猪毛菜,伴生种有灰菜、地梢瓜等。该类群丛分布较为广泛,在河流廊道各垂直梯度与水平梯度均有少量分布。
Ⅷ.狗尾草群丛(Ass.Setariaviridis)。草本层优势种为狗尾草,平均盖度为0.1—0.8,次优种为马唐、地梢瓜,伴生种有葎草、灰菜、苣荬菜(Sonchusarvensis)等。该类群丛分布较为广泛,在河流廊道各垂直梯度与水平梯度均有一定数量的分布。
Ⅸ.榆树群丛(Ass.Ulmuspumila)。乔木层优势种为榆树,郁闭度为0.4—0.9;草本层有少量尖头叶藜、马唐、灰菜、地梢瓜和苣荬菜。该类群丛数量少,仅在样带1及样带5少量出现。
在以上9类群丛中,狗尾草+猪毛菜+尖头叶藜群丛(群丛Ⅲ)包含最多的样方,占总样方数量的28.2%;狗尾草群丛(群丛Ⅷ)占比12.1%,加杨-狗尾草+蒺藜群丛(群丛Ⅳ)占比11.3%,以上三类群丛共占调查样方数量的一半以上。此外,9类群丛中共有乔木群落1类,占总样方数量的2.4%;草本群落3类,占比46.8%;乔木-草本群落5类,占比50.8%。从整体上看,永定河干涸段河流廊道植物群落结构较为简单,以乔木-草本结构和草本结构为主,灌木层十分缺失。
3.3 群落的物种多样性特征
3.3.1群落不同层次间物种多样性的比较
经统计,研究区域内各样方乔木层的Patrick指数变化范围在0—4之间、平均值为1.16,Shannon-Wiener指数变化范围在0—0.9之间、平均值为0.35,Pielou指数变化范围在0—0.9之间、平均值为0.21;草本层的Patrick指数变化范围在1—14之间、平均值为7.04,Shannon-Wiener指数变化范围在0.3—2.4之间、平均值为1.48,Pielou指数变化范围在0.0—1.0之间、平均值为0.78;灌木层由于样方过少不再单独进行分析。整体来看,草本层的三类物种多样性指数均大于乔木层,且丰富度指数和多样性指数变化幅度更大。
3.3.2群落物种多样性在垂直梯度上的空间分布特征
分别对干涸段上游至下游12个样带(海拔由53m降至29m)中各样方乔木层和草本层的物种多样性指数进行统计,以多样性指数为响应变量、样带号为解释变量进行回归分析[24],分析结果如图3所示,趋势线经四阶多项式回归拟合得到。
从图3中可以看出,乔木层各样方的Patrick指数和Shannon-Wiener指数在垂直梯度上的变化趋势基本一致,从指数最高的样带1开始先缓慢减小,在样带8、9达到最小值后又小幅度增大;Pielou指数的变化趋势稍有不同,呈现持续减小的趋势,仅在最后两个样带有小幅度的增大。草本层的多样性指数在垂直梯度上呈现更加波动的变化趋势,Patrick指数和Shannon-Wiener指数的变化基本一致,呈现先减小,到达样带3、4后增大,到达样带9、10后再次减小的趋势;Patrick指数的变化趋势稍有不同,在样带2、3达到最小值。对比乔木层和草本层的多样性指数变化,可以看到乔木层Patrick指数和Shannon-Wiener指数最小的样带8—10却是草本层两类指数最大的样带,说明乔木层物种组成越丰富、草本层物种的复杂程度越小;草本层各样带的Patrick指数整体较乔木层数值更大且变化更小,即草本层植物均匀程度更大、变化更小。
3.3.3群落物种多样性在水平梯度上的空间分布特征
由于研究区域内河流廊道宽度变化较大,为更直观地看出物种多样性在两堤内水平梯度的变化趋势,以样方中乔木层和草本层的物种多样性指数为响应变量、样方距左堤距离同河流廊道宽度的比值为解释变量进行回归分析。分析结果如图4所示,趋势线经四阶多项式回归拟合得到,展示了从左堤至右堤物种多样性指数的变化。

图4 乔木层、草本层的丰富度指数、多样性指数及均匀度指数在水平梯度上的变化趋势Fig.4 The variation trend of Patrick Index, Shannon-Wiener Index and Pielou Index of arbor layer and herbaceous layer on horizontal gradientD/W: 样方距左堤距离与河流廊道宽度的比值
如图4所示,乔木层的三类多样性指数在水平梯度上的变化趋势基本一致:沿左堤向内先大幅减小,Patrick指数和Shannon-Wiener指数在距左堤约3/10距离处达到低谷值后又逐渐增大、Pielou指数在距左堤约2/5距离处达到低谷值,三类指数在距右堤1/5距离(距左堤4/5距离)处开始几乎不再增大。草本层的Patrick指数和Shannon-Wiener指数呈现基本一致的“W”型变化趋势,从指数最高的左堤处开始,三个拐点分别为距左堤约1/5距离、距左堤约1/2距离和距右堤约1/10距离处; Pielou指数则呈现差别较大的变化趋势,先逐渐增大,在距左堤约3/10距离处时开始缓慢减小,在右堤处达到最小值。对比乔木层和草本层的物种多样性变化,可以看出干涸区河流廊道乔木层和草本层的Patrick指数和Shannon-Wiener指数在靠近左堤一侧呈现较为相似的变化趋势,但Pielou指数呈现相反的变化趋势。
4 结论和讨论
(1)本次研究共调查到永定河断流区河流廊道植物119种,其中乔木29种、灌木9种、草本81种,优势科为禾本科、菊科、豆科和杨柳科。测得的物种数少于滕依辰2013年对永定河山峡段河岸带测得的220种[19],可见永定河干涸段河流廊道物种的丰富程度相对匮乏,推测这与该段历史上河道泛滥改道造成的土地沙化以及常年断流造成的生态环境持续恶化[16]有关。经TWINSPAN分类,将河流廊道植物分为9个主要群丛类型,其中优势群丛为“狗尾草+猪毛菜+尖头叶藜群丛”、“狗尾草群丛”和“加杨-狗尾草+蒺藜群丛”,说明永定河干涸段河流廊道植被部分还处于演替初期,环境适应能力强的一年生草本群落分布十分广泛,乔木层物种较为单一。
(2)根据以往的研究,物种多样性在垂直梯度上的变化主要有负相关、在中等海拔高度最大等五种形式[25],但由于干涸段海拔变化较小,且受人类活动干扰较大,不适于用以上五种模式进行分析。通过分析干涸段乔木层的多样性指数在垂直梯度的变化趋势,可以看出上游植物群落乔木层的物种结构组成较下游来说更加丰富、复杂程度更高、物种分布更加均匀且优势种不明显。由于样带1—4位于平原城市段,受城市建设影响较大,在调研过程中发现河流廊道内存在较多大片的水源保护林、公益林等,并存在观赏性植被群落;样带5—12位于平原郊野段,人为干扰以农业干扰类型为主[26],乔木主要为堤侧防护林,以杨、柳为主。因此,越靠近城市人为干扰程度越大,乔木层的物种多样性越高。这与许多研究得出的人为干扰程度越大、物种多样性指数越低的结论相反[27-28],是因为永定河干涸段的人为干扰以生态破坏后的修复性造林活动为主。除人为干扰外,环境条件、种子传播等也会影响物种多样性的空间分布[29-30],但自然因素在干涸段对乔木物种多样性的影响程度相对较小。
通过分析干涸段草本层的多样性指数在垂直梯度的变化趋势,可以看出草本层从上游到下游物种组成的丰富程度和均匀程度先降低后升高、再降低,其中样带8—10的草本层物种的组成最复杂、分布最均匀。草本植物作为冠下层物种,其分布格局除受大的水热环境的影响之外,也受到其上层的乔木结构(如林冠郁闭度)、林下土壤、地形及其它局部小环境的影响[31],另外与农业种植等人类活动也有一定关系,因此要分析导致其变化的具体原因还需对草本群落的多样性指数和环境因子等做进一步的排序分析等相关性分析。
(3)理论上,河流廊道内离河的较近地区干旱胁迫越弱,物种多样性应越高[32],但由于研究区域内河道干涸已久,因此不再表现为自然的河流廊道植物分布特征。通过分析干涸段乔木层的多样性指数在水平梯度的变化趋势,可以看出越靠近左、右两堤,乔木层的物种结构组成越丰富、分布越均匀,左堤的物种组成较右堤更加复杂且分布更加均匀。由于历史上永定河多次改道,加上人类的非法采砂等活动,河流廊道内部留下大面积的沙地[16],现状以草本植被为主,乔木较少;另外,调研时发现北京市的造林工程集中分布于左堤一侧,以成片的防风固沙林为主,右堤虽然也分布有连续的带状防护林,但林带较窄且物种组成较为单一,因此,人类的破坏和造林活动范围是影响乔木层物种多样性分布的主要原因。草本层的物种多样性在水平梯度上变化较大,但与乔木层一样,在靠近左堤的位置物种组成最丰富,要具体解释导致其多样性格局变化的原因还需对草本群落的多样性指数与水分、土壤、地形等环境因子及人为干扰等做进一步的相关性分析。
致谢:感谢北京林业大学园林学院朱妙馨、王琛瑶、孟子卓、郭若木在实地调研工作中给予的帮助。
猜你喜欢
中国野生植物资源(2022年12期)2023-01-09
生态学报(2022年19期)2022-10-27
中国水利(2022年9期)2022-06-02
作文周刊·小学一年级版(2020年40期)2020-10-19
中华诗词(2020年2期)2020-09-21
绿色中国(2019年24期)2019-11-26
西南农业学报(2018年6期)2018-08-04
中国水土保持科学(2016年3期)2016-07-19
小学生作文选刊·低年级版(2016年6期)2016-06-24
中国水利(2015年16期)2015-02-28
