
华中五味子根际土壤真菌群落特征研究
2021-01-15 08:20江林春韦洁敏白尚尚崔浪军刘世鹏
延安大学学报(自然科学版) 2020年4期
江林春,韦洁敏,白尚尚,强 毅,崔浪军,刘世鹏*
(1.延安大学生命科学学院,陕西延安716000;2.西北濒危药材资源开发国家工程实验室药用资源与天然药物化学教育部重点实验室,陕西师范大学生命科学学院,陕西西安710119)
华中五味子(SchisandrasphenantheraRehd.Et Wils.)为木兰科(Magnoliaceae)五味子属(Schisandra)多年生落叶木质藤本植物,以干燥成熟果实入药。中药名为南五味子,为我国传统中药材[1],被收录于《中华人民共和国药典》(2020年版)[2],有收敛固涩、益气生津、补肾宁心之功效[3],食用和药用价值高。在我国主要分布于秦巴山区,且以野生抚育资源为主[4]。华中五味子为须根系藤本植物,须根发达[5],常与伴生树种组成群落,具有较强的截流雨水、减缓雨水冲刷坡面和固土的能力,已成为秦巴山区水土保持和药农脱贫致富的主要药材品种。近年来华中五味子在秦巴山区规模化人工种植和野生抚育的面积逐年增加,如何进行科学田间管理和野生抚育管理是目前亟待解决的问题。了解华中五味子自然状况下根际土壤化学性质、真菌多样性与群落结构的变化对于华中五味子田间管理与野生抚育方案的确定具有重要的指导意义,但目前尚未见这方面的报道。基于此,本文系统研究了华中五味子2年生苗种植3年和6年后根际土壤化学性质和土壤真菌群落结构的变化规律,分析了二者的相互关系,以期为华中五味子的科学田间管理与野生抚育方案的确定提供一定的理论指导。
1 材料和方法
1.1 试验时间、地点
本试验于2013年3月在陕西省商洛市柞水县下梁镇布置试验田。试验田翻土后起隆,按株行距1 m×1.5 m栽种两年生健康华中五味子苗。正常田间管理,包括拔草、灌溉、搭架等,但不施肥,拔草和修剪后将杂草和修剪枝条移出试验田,但秋季落叶保留在试验田不清理。
1.2 试验材料
2013年3月、2016年3月和2019年3月采用5点取样法采集根际土壤样本,标记为CK、Y3和Y6。收集的土壤样品过2 mm孔径筛子后一部分风干用于测定土壤化学性质;另一部分低温保存,用于后续土壤微生物真菌多样性的确定。
1.3 试验方法
1.3.1 土壤化学性质测定
大量与微量元素含量的测定用PW2403型X-Ray荧光光谱仪(德国Bruker公司)。铵态氮、磷酸盐的含量测定用SmartChem300全自动间断化学分析仪(意大利Westco公司)。
1.3.2 土壤样品高通量测序
利用试剂盒(天根生化科技(北京)有限公司)提取土壤样本基因组DNA后,根据保守区设计得到引物,采用真菌ITS1区通用引物[6],在引物末端加上测序接头,以样本DNA为模板,进行PCR扩增并对其产物进行纯化、定量和均一化形成测序文库,并利用1%的琼脂糖凝胶电泳检测总DNA提取质量[7]。选择质量合格的DNA样品进行真菌ITS1区Paired-end测序,由北京百迈客科技有限公司完成,使用Illumina HiSeq平台进行测序,数据分析参考文献方法[8]。
1.4 数据处理与分析
采用Microsoft Excel 2010进行平均数和标准误的计算,采用SPSS19.0软件对数据进行单因素方差处理分析。
2 结果与分析
2.1 不同年限华中五味子根际土壤化学性质含量变化
研究表明,3个样本中铵态氮(NH3-N)、速效磷(A-P)、S含量显著上升(P<0.05),其中铵态氮、S的含量均为Y3>Y6>CK,速效磷的含量为Y6>Y3>CK。Y3中铵态氮、S的含量分别比CK增加108.18%、139.06%。而随华中五味子种植年限延长,土壤中微量元素Fe3+、Mn2+、Zn2+、Pb的含量均低于CK,但无显著差异(P>0.05),总体呈现下降趋势(表1)。
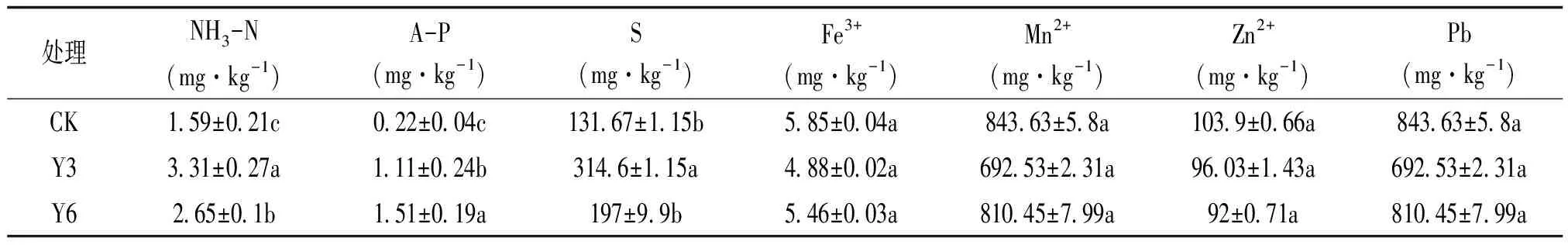
表1 不同年限华中五味子根际土壤化学性质含量
2.2 测序数据的合理性分析
通过Illumina HiSeq测序平台对土壤样品进行真菌多样性检测分析,获得真菌有效序列46 782条,序列的长度主要分布在191 bp-280 bp的区段内,其中221 bp-250 bp区段内分布最多。由于序列长度是将序列匹配到数据库从而产生基因注释的关键,因此根据这些优质序列分析得到的物种多样性结果科学、真实,具有一定的参考价值[9]。图1显示随着随机抽取的样本真菌测序量不断增大,各样品OTU数目的增加趋于平缓,说明每个样品测序数量已经满足数据分析需求,因此测序数据充足、合理,可以用于后续的生物信息学分析。
2.3 根际土壤真菌的数量分析
选取属水平的OTU列表,在相似度为97%的水平下对OTU的聚类结果进行分析。由图2可知,在属水平上,随着种植年限的增加,土壤真菌的OTU数目呈现一个增加的趋势。3个样本中真菌的物种总数为962种,共有物种数量为389种,CK特有的物种数为55种,Y3特有的物种数为135种,Y6特有的物种数为70种。
2.4 华中五味子根际土壤真菌生物多样性
通过微生物多样性指数比较不同年限华中五味子根际土壤真菌的α多样性(表2),CK、Y3与Y6的根际土壤真菌的Ace指数和Chao1指数大小均呈现为Y3>Y6>CK,表明Y3的土壤真菌丰度最高,Y6次之,CK最低。CK、Y3与Y6的Shannon指数分别为4.48、4.16和4.41,呈现为:CK>Y6>Y3,表明CK的土壤真菌群落多样性最高,Y6次之,Y3多样性最低。
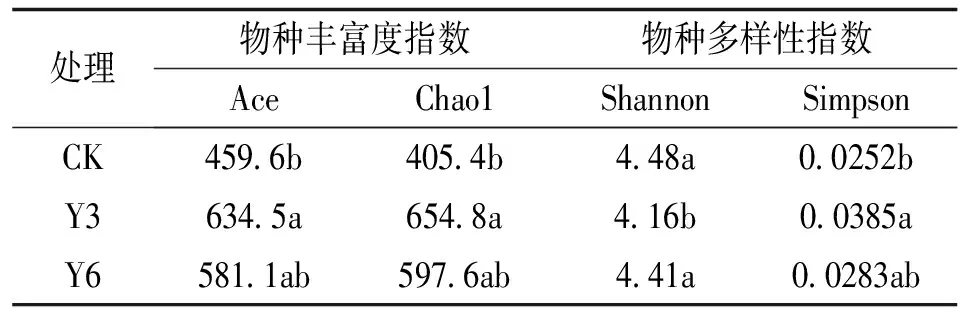
表2 不同年限华中五味子根际土壤真菌α多样性指数
2.5 华中五味子根际土壤真菌群落组成的比较分析
使用QIIME软件进行β多样性(Beta diversity)分析,比较不同样品在物种多样性方面存在的相似程度。根际土壤样本中真菌群落的主成分1(PC1)和主成分2(PC2)的样品差异性贡献率分别达到61.29%和26.07%(见图3)。CK、Y3与Y6均分布在PC1和PC2的区域内,并且每个种植年限根际土壤样品中的真菌群落相对集中。图3表明CK和Y62组根际土壤样品内真菌群落组成相似,与Y3的土壤样品真菌群落组成有差异。
2.6 华中五味子根际土壤真菌群落结构组成分析
由各个样品在科水平和属水平上的真菌群落物种丰度热图(图4)可知,在供试的3个土样中共注释出18个相对丰度较大真菌科类(图4a):其中CK中毛壳菌科Chaetomiaceae(12.54%)、赤壳科Nectriaceae(19.27%)、被孢霉科Mortierellaceae(7.93%)、毛球壳科Lasiosphaeriaceae(3.03%)、枝孢霉科Cladosporiaceae(4.62%)的相对丰度较大。Y3中Plectosphaerellaceae(11.20%)、亚隔孢壳科Didymellaceae(14.39%)、Mrakiaceae(12.03%)、柔膜菌科Helotiaceae(5.77%)的相对丰度较大。Y6中毛壳菌科Chaetomiaceae(12.22%)、Piskurozymaceae(6.63%)、球腔菌科Mycosphaerellaceae(3.32%)、赤壳科Nectriaceae(19.63%)和Piskurozymaceae(6.63%)的相对丰度较大。由图4(b)可知,CK的优势真菌属为枝鼻菌属Cladorrhinum(2.56%)、分子孢子菌属Cladosporium(4.62%)、腐质霉属Humicola(4.13%)、被孢霉属Mortierella(7.86%)和尖孢镰刀菌属Fusarium(6.98%);亚隔孢壳属Didymella(10.57%)、赤霉菌属Gibberella(2.40%)、Tausonia(10.69%)和Tetracladium(5.56%)为Y3的优势真菌属;青霉菌属Penicillium(1.52%)、紧密帚枝霉属Sarocladium(4.85%)、Solicoccozyma(6.63%)为Y6的优势真菌属。
真菌群落热图中CK、Y3和Y6真菌群落结构差异性较大。CK的优势真菌属随着华中五味子种植年限的增加呈现总体降低的趋势:分子孢子菌属Cladosporium在CK、Y3及Y6中所占比例分别为4.62%、4.42%、0.71%,腐质霉属Humicola在CK、Y3及Y6中所占比例分别为4.13%、3.85%、2.89%。被孢霉属Mortierella在CK、Y3及Y6中所占比例分别为7.86%、1.23%、2.31%,尖孢镰刀菌属Fusarium在CK、Y3及Y6中所占比例分别为6.98%、1.07%、3.35%。
2.7 真菌群落结构与土壤化学性质相关性分析
从冗余分析(RDA)图(图5)结果发现,CK、Y3及Y6 3个样本中各类真菌与土壤化学因子分布在3个象限内。对土壤中的优势真菌菌群(属水平)和土壤化学性质进行相关分析,其中铵态氮对根际土壤样品中优势菌属如被孢霉属Mortierella、尖孢镰刀菌属Fusarium、分子孢子菌属Cladosporium、腐质霉属Humicola的相关性影响最大,Mn2+对根际土壤样品中优势真菌的相关性影响最小。被孢霉属Mortierella、尖孢镰刀菌属Fusarium与Mn2+、Zn2+呈正相关,与速效磷、铵态氮、S呈负相关;分子孢子菌属Cladosporium、腐质霉属Humicola与Mn2+、Zn2+呈正相关,与速效磷、铵态氮呈负相关。
3 讨论
本研究表明,随华中五味子种植时间延长,根际土壤的化学性质的含量发生了变化。方差分析结果显示,种植华中五味子3年、6年后,土壤中铵态氮、速效磷、S的含量显著增加(P<0.05),这与在怀牛膝[10]、牡丹[11]和藤本植物[12]上的研究结果一致。其原因可能是华中五味子生长期内对N、P和S的需求量由高变低,同时,果实成熟末期,营养物质积累运输至根,根部储存养分并随着根系分泌物及地上的枯落物将养分输送到土壤。李爱民[13]和丛毓[14]对北五味子的研究结果亦得出了这样的结论。卜海东[5]对华中五味子地上部分生长发育动态研究,结果显示,在整个生长期内华中五味子对微量元素Fe3+、Mn2+、Zn2+均有较大的需求量,需要从土壤中大量吸收这些元素来满足生长所需。本研究的结果中华中五味子种植年限增加,土壤微量元素Fe3+、Mn2+、Zn2+、Pb的含量下降也侧面验证了这一结果。可见,根际土壤理化性质能够体现土壤的健康状况[15],同时也反映出华中五味子植株生长与发育所需要的营养元素状况。
α多样性指数显示,华中五味子随种植年限延长,根际土壤真菌群落丰富度增加,物种多样性降低。这与根系的作用有关。研究表明,根系分泌物可增强根际土壤真菌的活性[16],改变其多样性和群落结构。土壤理化特性的改变可间接影响土壤微生物的分布与活性。对崂山茶区茶树根围AM真菌多样性的研究证明,随着栽培时间增长,AM真菌优势菌种不断积累,非优势菌种逐渐消失,物种多样性降低[17],这与本研究具有相似之处。其可能原因为华中五味子作为多年生植物,随着栽培时间增加,根系形态和根系分泌物的变化,导致优势真菌数量增加,非优势真菌种逐渐减少直至消失,使真菌在根际间的种类和分布产生了显著差异,进而影响真菌的种群结构和代谢功能多样性,导致土壤真菌群落多样性降低。华中五味子植物须根发达,枯落物丰富,随着种植年限延长,在通过根系分泌物以及枯落物对土壤化学性质的影响较大,使土壤养分提高,进而使得土壤真菌群落分布不均,通过长期生长的过程中一定程度上改变了根际土壤环境,变动了土壤稳定结构,增大土壤微生物丰度和均匀度,从而有利于土壤微生物的代谢,使得土壤真菌群落丰富度增大。
β多样性结果表明,CK与Y6华中五味子的根际土壤真菌群落结构在科水平和属水平相似。赤壳科Nectriaceae属肉座菌目,已有研究发现归属于子囊菌门的肉座菌目具有降解腐烂有机质,降解纤维素的作用[8]。尖孢镰刀菌属Fusarium是引起枯萎病的主要病原菌属[18-20]。分子孢子菌属Cladosporium真菌是一种包括室内和室外都常见的霉菌,为腐生真菌,广泛存在于自然界的土壤、某些动物的粪便、蔬菜、腐木、鸟巢、腐烂水果中,会导致果实腐烂[21]。腐质霉属Humicola真菌是土壤中常见的腐生菌,广泛分布于世界各地的各种土壤,尤其是有机质丰富的土壤,部分种类也分布于植物残体、木材或人体等[22]。被孢霉属Mortierella是土壤中有益微生物,具有溶解土壤中磷的功能[23],并且能够通过提供氮营养和提高植物的抗病性来促进植物的生长,可能与植物的糖代谢有关[24]。从属水平分析结果来看,不同年限华中五味子根际土壤的优势真菌属存在差异性,说明随着华中五味子种植年限增加,优势真菌属一直在变动,其中尖孢镰刀菌属Fusarium、分子孢子菌属Cladosporium、腐质霉属Humicola等病原菌的生长可能会受到抑制,一定程度上改善了华中五味子根际土壤环境,提高了土壤的健康状况,促进华中五味子植株的生长发育。
土壤化学因子与土壤微生物群落显著相关,土壤环境因子的改变会影响土壤中微生物群落结构[25,26]。有研究发现,土壤化学性质不仅能够反映土壤的健康状况,还能够影响土壤微生物种群数量及分布,为土壤根际微生物活动提供营养和能源[27]。本研究对土壤真菌群落与土壤化学性质的相关分析表明,随着华中五味子种植时间延长,土壤化学性质与各真菌群落间具有一定的相关性,其中:速效磷、铵态氮、S与尖孢镰刀菌属Fusarium、被孢霉属Mortierella呈现负相关,与微量元素Mn2+、Zn2+呈正相关;分子孢子菌属Cladosporium、腐质霉属Humicola与速效磷、铵态氮呈负相关,与Mn2+、Zn2+呈正相关。土壤中速效磷、铵态氮、S、Mn2+、Zn2+是影响华中五味子根际土壤真菌群落的主要肥力因子,说明这些土壤化学因子对于这些根际土壤真菌群落遗传多样性起着重要作用。这与王玉等[17]和杨俊博[28]的研究结果相似,土壤中的氮元素等对真菌多样性有显著影响,各个因子会显著影响相关真菌群落。表明可以通过调节土壤养分水平,来改变华中五味子根际土壤真菌群落组成和遗传多样性水平。
综上所述,种植华中五味子后,植株生长发育对微量Fe3+、Mn2+、Zn2+、Pb营养元素的需求量较大,导致根际土壤中Fe3+、Mn2+、Zn2+、Pb等元素的含量降低;随华中五味子生长期的延长,植株根系分泌物以及枯落物导致根际土壤中铵态氮、速效磷、S等的含量显著增加,提高了土壤的保肥能力和补给能力。又因土壤化学性质的改变也会影响土壤真菌区系的变化,土壤中真菌群落的生理活动与土壤环境中的化学性质的含量密切相关,最终导致尖孢镰刀菌属Fusarium、分子孢子菌属Cladosporium、腐质霉属Humicola等根际土壤真菌群落改变。
猜你喜欢
当代水产(2022年7期)2022-09-20
土壤学报(2022年2期)2022-06-09
凤凰动漫(军事大王)(2022年1期)2022-04-19
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
湘潮(上半月)(2021年10期)2021-12-02
当代水产(2021年1期)2021-03-19
家庭科学·新健康(2017年6期)2017-06-16
江苏农业科学(2016年10期)2017-02-05
江苏农业科学(2016年5期)2016-07-23
药学研究(2015年11期)2015-12-19
