
Obeticholic acid attenuates human immunodeficiency virus/alcohol metabolism-induced pro-fibrotic activation in liver cells
2021-01-13 07:58MosesNewAaronMuraliGanesanRaghubendraSinghDagurKusumKharbandaLarisaPoluektovaNataliaOsna
World Journal of Hepatology 2020年11期
Moses New-Aaron, Murali Ganesan, Raghubendra Singh Dagur, Kusum K Kharbanda, Larisa Y Poluektova, Natalia A Osna
Moses New-Aaron, Department of Environmental, Agriculture and Occupational Health, College of Public Health, University of Nebraska Medical Center, Omaha, NE 68105, United States
Moses New-Aaron, Murali Ganesan, Raghubendra Singh Dagur, Kusum K Kharbanda, Natalia A Osna, Research Service, Veterans Affairs Nebraska-Western Iowa Health Care System, Omaha, NE 68105, United States
Murali Ganesan, Raghubendra Singh Dagur, Kusum K Kharbanda, Natalia A Osna, Department of Internal Medicine, University of Nebraska Medical Center, Omaha, NE 68105, United States
Larisa Y Poluektova, Department of Pharmacology and Experimental Neuroscience, University of Nebraska Medical Center, Omaha, NE 68198, United States
Abstract
Key Words: Human immunodeficiency virus; Liver; Obeticholic acid; Alcohol; Hepatocytes; Fibrosis
INTRODUCTION
Liver disease is a second-leading cause of mortality in human immunodeficiency virus (HIV)-infected patients[1].While effective anti-retroviral treatment (ART) has dramatically decreased the onset of acquired immunodeficiency syndrome, the morbidity and mortality are often associated with liver disease, which progresses slowly into severe liver dysfunction.This progression is related not only to the ARTinduced hepatotoxicity, but to HIV properties likely associated with progression of non-alcoholic fatty liver disease/non-alcoholic steatohepatitis (NASH)[2,3].There are multiple insults, which promote HIV-related liver injury, including dysregulation of lipid metabolism and fat turnover, co-infections with hepatotropic viruses and alcohol abuse[4,5].
Recently, we have shown that alcohol metabolite, acetaldehyde promotes accumulation of HIV proteins in hepatocytes, inducing oxidative stress and cell apoptosis[6].Furthermore, the engulfment of these HIV-containing hepatocytes by nonparenchymal cells, macrophages and hepatic stellate cells (HSC) causes inflammasome activation in macrophages and pro-fibrotic activation of HSC[6].This indicates that massive apoptosis of infected hepatocytes may induce continuous activation of HSC leading to liver fibrosis development.Since acetaldehyde-triggered HIV replication in hepatocytes is abortive[6], HIV accumulation cannot be fully controlled by ART and requires the treatment with additional anti-fibrotic drugs.
One of the promising drugs with anti-fibrotic activity is obeticholic acid (OCA).In 2016, OCA became the United States Food and Drug Administration-approved to treat primary biliary cholangitis and currently is at phase 3 trial (Regenerate) to treat fibrosis caused by NASH[7].OCA is a farnesoid-X receptor (FXR) agonist, which binds to the FXR in the nucleus of liver and intestinal cells.Multiple genes are activated by this pathway, including the control of metabolism of bile acids, lipids, glucose, and amino acids.FXR is highly expressed on hepatocytes and immune cells, and is involved in pathogenesis of viral hepatitis, alcohol-and non-alcohol-induced liver disease[8].In NASH-fibrosis, OCA regulates liver injury progressionviatargeting of gut microbiota[9].Currently, it is not quite clear whether OCA directly modulates hepatocyte apoptosis: While some studies reported the lack of anti-apoptotic effects of OCA on these cells[10], other studies demonstrated the reduction of hepatocyte apoptosis by OCA-mediated suppression of metabolic stress and prevention of subsequentp53activation, with further anti-fibrotic and anti-inflammatory downstream effects[11].These studies were mainly performed on experimentalin vivomodels, which makes difficult to exclude the effects of OCA on gut microbiota, narrowing down the mechanisms to only OCA-regulated hepatocyte apoptosis.None of publishedin vivoorin vitrostudies have been performed in the context of the effects of OCA on HIV- and alcohol-induced liver injury.However, activation of HSC by engulfment of HIV-expressing apoptotic hepatocytes generated as a downstream event in HIV and ethanol metabolism-induced oxidative stress is one of the reasons for liver fibrosis progression, which serves as an important target to prevent end-stage liver disease development.Thus, based on already characterized mechanisms of liver injury progression triggered by the combination of HIV with acetaldehyde[6], we aimed to investigate whether OCA protects from apoptotic hepatocyte death and from activation of HSC by engulfment of apoptotic HIV-infected hepatocytes.
MATERIALS AND METHODS
This is the original (basic) study performed at University of Nebraska Medical Center, Omaha, NE, United States.Here, for the first time,in vitroapproach is used to characterize the ability of OCA to reverse the pathology induced by HIV and ethanol metabolism in liver parenchymal and non-parenchymal cells.In our study, OCA has been tested as an anti-fibrotic drug, which affects the pathogenesis of HIV-alcoholinduced liver fibrosis development.
Reagents and media
High glucose Dulbecco’s modified eagle medium and fetal bovine serum were purchased from Invitrogen (Carlsbad, CA, United States), Trizol was from Life Technologies, primer probes and real-time polymerase chain reaction (RT-PCR) reagents were from Applied Biosystems by Thermo Fisher Scientific, CA , United States.Other reagents, all analytical grade quality, were from Sigma (St.Louis, MO, United States).
Cells and treatments
As experimental prototype of human primary hepatocytes, we used Huh7.5-CYP (RLW) cells.These cells have reduced innate immunity and can be infected with HIV.To metabolize ethanol, they were stably transfected by CYP2E1, but do not express alcohol dehydrogenase (ADH).To overcome this limitation and mimic natural ethanol metabolism in primary hepatocytes, we treated RLW cells with acetaldehydegenerating system (AGS), which contains yeast ADH as a source of enzyme, NAD as a co-factor, and 50 mmol/L ethanol as a substrate for ADH and continuously produce physiologically relevant amount of acetaldehyde (Ach) without toxic effects.We have characterized and successfully used these cells and AGS for HCV-based ethanol liver studies[12-14].Cells were pre-treated for 24 h with AGS and then exposed to HIVADA(MOI 0.1) for 48 h.To investigate the effects of OCA, cells were pre-treated with OCA (50 μmol/L) for 4 h before experiment.
HIV RNA, HIV DNA, integrated HIV DNA, western blot and reactive oxygen species
HIV RNA was detected by RT-PCR; HIV DNA was detected by semi-nested PCR;integrated HIV DNA was measured by digital droplet PCR; reactive oxygen species (ROS) were quantified by DCF (2’,7’–dichlorofluorescein fluorescence method); western blot was performed as described.The details of all these methods were already published[6].
Apoptosis measurements:
Apoptosis in RLW cells was measured by caspase 3 and PARP cleavage in cell lysates (western blot) and M30 Apoptosense ELISA (Duapharma group, Inc.West Chester, OH, United States) in cell supernatants.
Activities of proteasome and cathepsins
Proteasome activities and cathepsin B and L activities were assayed fluorometrically as described previously[15,16].
Apoptotic bodies
Apoptotic bodies (AB) were generated from uninfected and HIV-infected RLW cells by exposure to UV light and characterized as shown previously[13].
Hepatic stellate cells and treatments with AB
As the source of human HSC, we used commercially available human cell line, LX2 (EMD Millipore, cat SCC064) grown based on instructions from the manufacturer.AB from RLW cells (AB Hep), both uninfected (control) and HIV-infected (ABHIV), were incubated with LX2 cells for 2-8 h at 1: 3 ratio and then pro-fibrotic markers [Col1A1, transforming growth factor (TGF)-β] and inflammasome-related parameters [NLRP3, caspase 1, interleukin (IL)-6, IL-1β] were quantified by RT-PCR.
Statistical analyses
Data were analyzed using GraphPad Prism v7.03 software (GraphPad, La Jolla, CA, United States).Data from at least three duplicate independent experiments were expressed as mean ± SEM. Comparisons among multiple groups were performed by one-way ANOVA, using a Tukey post-hoc test.For comparisons between two groups, we used Student’st-test.APvalue of 0.05 or less was considered significant.
RESULTS
Our previous studies on primary human hepatocytes exposed to ethanol and RLW cells treated with AGS demonstrated, first, that incubation of ADH-non-expressing RLW cells with AGS recapitulated the effects of ethanol on ethanol-metabolizing hepatocytes and second, that the highest levels of apoptotic hepatocyte death were observed when cells were exposed to both insults (ethanol/AGS and HIV)[6].That is why in this paper, we presented the data on ability of OCA to protect cells from the harmful effects of AGS on HIV-infected hepatocyte-like RLW cells to mimic ethanol metabolism observed in primary human hepatocytes.For OCA screening, we did not use primary human hepatocytes since their supply was limited and they quickly (in 24 h) de-differentiate to loose ethanol-metabolizing capacity.
OCA attenuates AGS-HIV-induced apoptotic cell death and oxidative stress
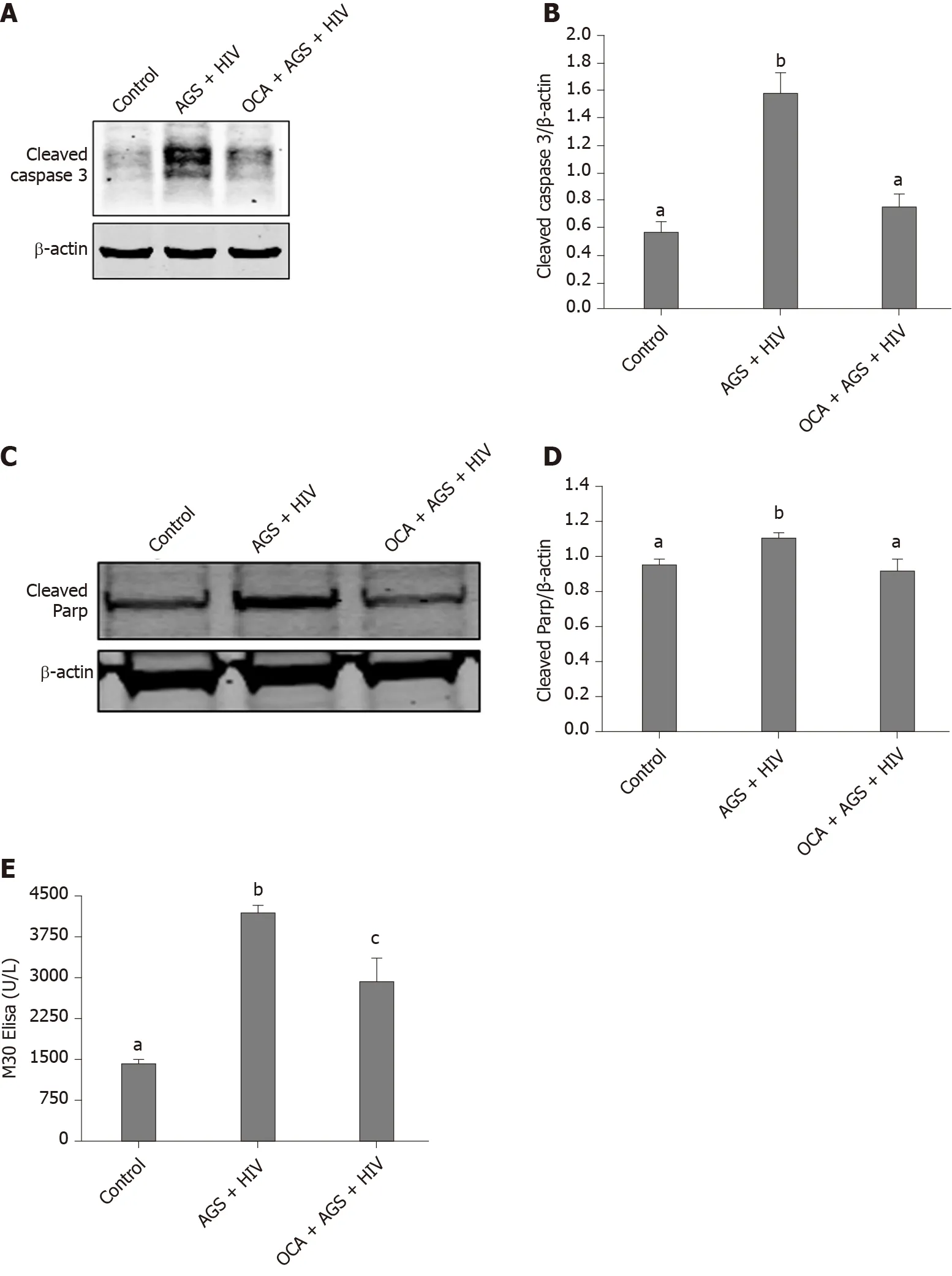
Figure 1 Obeticholic acid attenuates apoptosis in acetaldehyde-generating system-human immunodeficiency virus-exposed Huh7.5-CYP cells.
We found that while AGS + HIV induced caspase 3 cleavage almost three-fold, pretreatment with OCA suppresses these effects (Figure 1A and B).The same trend (but with lower magnitude of response to AGS) was observed on cleaved PARP, and OCA also suppressed it (Figure 1C and D).Measuring cleaved cytokeratin 18 by M30 ELISA in cell supernatants, we found that OCA attenuated AGS-HIV-induced apoptosis in hepatocytes (Figure 1E).Since apoptosis may be induced by elevatedp53, we measured the induction ofp53mRNA by AGS+HIV, in the presence or absence of pancaspase inhibitor or OCA.As shown on Figure 2A, while AGS and HIV inducedp53mRNA level two-fold, both OCA and pan-caspase inhibitor reducedp53expression only by 14%.In addition, OCA attenuated AGS-HIV-induced ROS production (Figure 2B), thereby protecting liver cells from oxidative stress.
OCA suppresses accumulation of HIV in hepatocytes
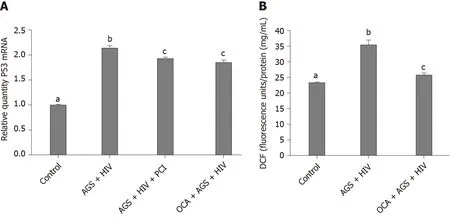
Figure 2 Effects of obeticholic acid on p53 mRNA and reactive oxygen species production in human immunodeficiency virus-infected Huh7.5-CYP cells exposed to acetaldehyde-generating system.
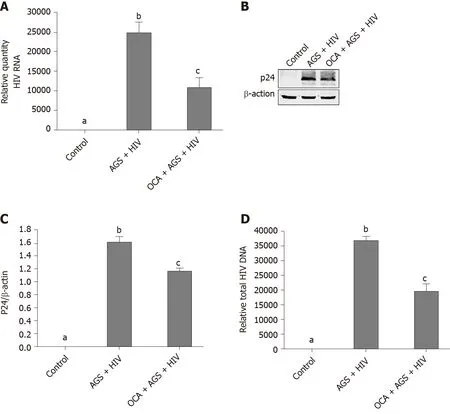
Figure 3 Obeticholic acid suppresses human immunodeficiency virus expression in Huh7.5-CYP cells exposed to acetaldehydegenerating system + human immunodeficiency virus.
We measured the effects of OCA on accumulation of HIVgag RNA (RT-PCR) and p24 (western blot) induced by AGS + HIV exposure to RLW cells.In these experiments, OCA suppressed the treatment-induced HIVgag RNA levels by 50% (Figure 3A) and p24 –by 26% (Figure 3B and C).OCA also decreased the level of HIV DNA in RLW cells by about 60% (Figure 3D), with no integration of HIV DNA in cell genome.As has been shown by us before, the accumulation of HIV components in RLW cells is controlled by HIV-AGS-decreased lysosomal and proteasomal activities[6].In fact, we observed partial restoration of chymotrypsin-like proteasome and cathepsin B and L lysosomal activities by OCA; however, OCA did not restore AGS-HIV-affected trypsin-like proteasome activity (Figure 4A-D).
OCA reduces pro-fibrotic activation of HSC by engulfment of HIV-containing apoptotic hepatocytes
AB generated from uninfected (control) or HIV-infected RLW cells were exposed to LX2 cells, followed by measurement of Col1A1 and TGF-β mRNAs by RT-PCR.We found that HIV-containing apoptotic cells induced more prominent pro-fibrotic activation of HSC than uninfected apoptotic cells; OCA suppressed this pro-fibrotic HSC activation by uninfected AB and to most extend, by ABHIVengulfment (Figure 5A and B).
To characterize the effects of OCA on inflammasome activation in HSC, we measured NRLP3, caspase 1 and pro-inflammatory cytokines, IL-6 and IL-1β mRNA expression induced by engulfment of uninfected AB and HIV-containing AB, Upregulation of NRLP3, caspase 1, IL-6 and IL-1 β levels by HIV-containing AB was successfully prevented by OCA treatment (Figure 6).
DISCUSSION
As previously shown, HIV accumulation induced by pre-exposure of cells to ethanol metabolites, mainly acetaldehyde produced by AGS, induces oxidative stress and apoptosis in hepatocytes.This is beneficial due to infected hepatocytes clearance before the integration of HIV DNA into human genome occurs.However, intensive hepatocyte apoptosis may have detrimental outcomes since HIV-containing apoptotic hepatocytes induce pro-fibrotic activation of HSC, thereby promoting fibrosis development[6].Here, we investigated whether OCA protects from AGS-HIV-induced hepatocyte apoptosis, which causes HSC activation by ABHIVengulfment to drive liver fibrosis development.
In our model, OCA pre-treatment attenuated apoptosis (caspase 3 and PARP cleavage as well as cleaved cytokeratin 18 expression) in AGS-HIV-exposed liver cells.Unlike suppression ofp53expression on liver cells by OCA reported in HIV-noninfected cells by others[11], here, OCA mildly suppressed AGS-HIV-inducedp53mRNA.Apoptotic hepatocyte death was triggered by oxidative stress induced by AGS and HIV in CYP2E1-expressing RLW cells.In our hands, OCA indeed suppressed ROS production, thereby attenuating oxidative stress, which corroborated the data obtained on different models[17,18].Here, the suppression of oxidative stress by OCA restores proteasome and lysosome functions, which increases the degradation of HIV proteins[19,20]and thus, diminishes the expression of HIV gag RNA and p24 gag protein in infected RLW cells.However, as we established before, the prevention of HIV and AGS- induced apoptotic hepatocyte death by exposure to pan-caspase inhibitor causes accumulation of cells with integrated HIV DNA[6], which is an unwanted event.Importantly, while OCA pre-treatment suppresses apoptosis in HIV-infected hepatocytes, there was no increase in hepatocytes expressing integrated HIV DNA.
In addition to beneficial effects of OCA on attenuation of HIV-AGS-induced hepatocyte death and reduction of HIV markers expression, OCA also reverses profibrotic activation (based on Col1A1 and TGF-β mRNA levels) of HSC by engulfment of HIV-containing apoptotic hepatocytes.Similar event was observed by[10]on CCl4-injured mice, indicating that OCA controls HSC activation triggeredviamultiple mechanisms.One of this mechanisms contributing to liver fibrosis progression is an activation of inflammasome pathway in HSC[21].In fact, we observed the reversing effect of OCA on this pathway in HSC, which supports anti-fibrotic activity of OCA.Thus, the suppression of pro-fibrotic activation by OCA in HSC is crucial for liver fibrosis development.
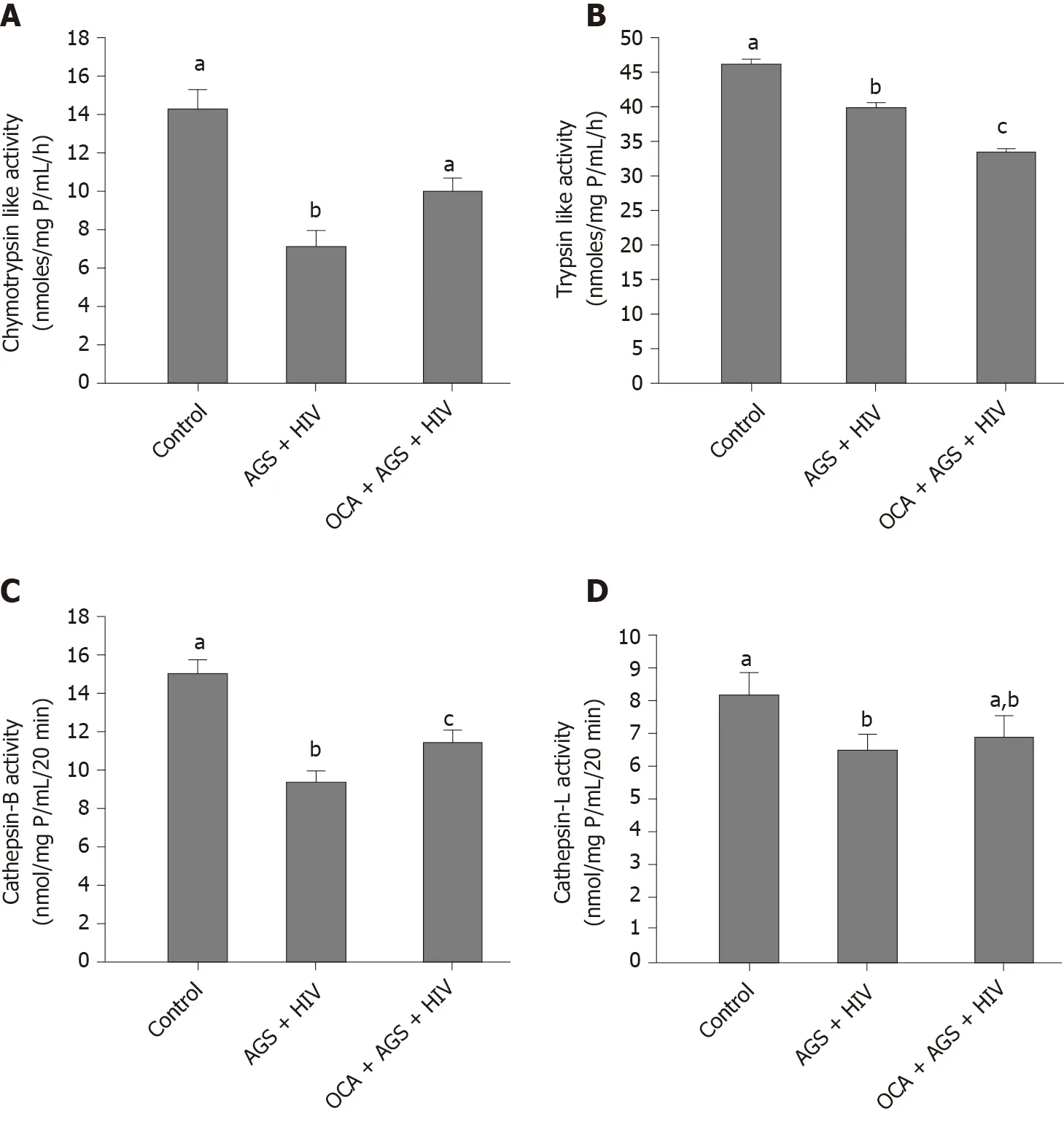
Figure 4 Restorative effects of obeticholic acid on proteasomal and lysosomal activities in acetaldehyde-generating system-human immunodeficiency virus-exposed Huh7.5-CYP cells.
In fact,in vivoprotection from early alcohol-induced liver damage by OCA has been already demonstrated on alcohol-fed uninfected mice without disclosing the mechanisms, by which this happened[22].Furthermore, in monocytes from peripheral blood of HIV-infected patients, the expression of nuclear receptors, including FXR, was reported to be low[23], and the treatments with FXR agonists like OCA might play a beneficial role.Importantly, in HIV-infected alcohol-abused patients, the efficacy of OCA treatment has never been tested.Furthermore, these studies have not been done in the context of HIV-affected liver function.Our experiments providesin vitroevidence for protective effects of OCA from liver fibrosis progression induced by HIV and ethanol metabolism.The major limitation of this innovative study is that the results are currently based only on in vitro, but notin vivoexperiments.Nevertheless, thesein vitroexperiments are necessary to characterize the exact mechanisms, by which OCA prevents HIV/ethanol metabolism-induced liver injury.These mechanisms are difficult to identify byin vivostudies, due to multiple triggers of liver fibrosis progression.We plan to confirm thein vitroeffects of OCA in future byin vivostudies on humanized mice model since only these mice can be infected with human live HIV and fed the liquid ethanol-containing diet.Thus, our currentin vitrostudy pioneers in justifying OCA inclusion to the treatment scheme of HIV-infected alcohol abusers with high risk of liver fibrosis development.
CONCLUSION
In conclusion, in HIV-infected hepatocytes exposed to continuously released acetaldehyde, OCA attenuates apoptotic death of infected cells and pro-fibrotic activation of HSC by engulfment of apoptotic HIV+hepatocytes, thereby protecting from liver fibrosis development.
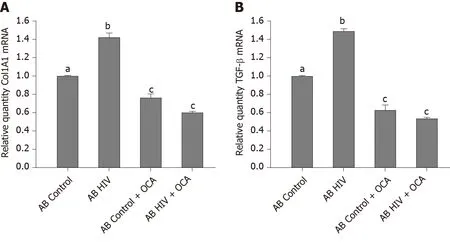
Figure 5 Anti-fibrotic effects of obeticholic acid on hepatic stellate cells activated by engulfment of apoptotic Huh7.5-CYP cells.
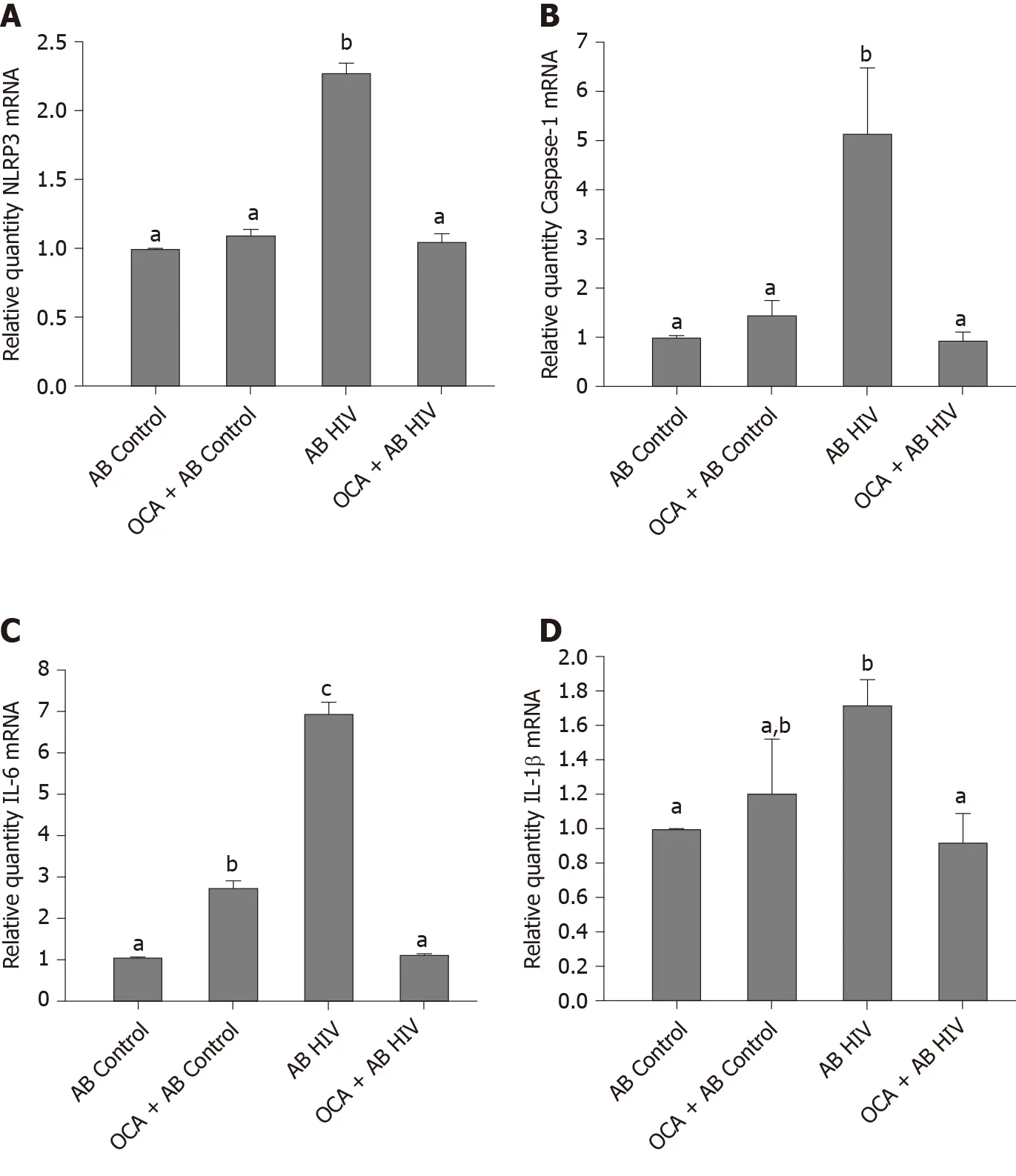
Figure 6 Obeticholic acid suppresses acetaldehyde-generating system-human immunodeficiency virus-induced activation of inflammasome, mRNAs.
ARTICLE HIGHLIGHTS
Research background
Due to frequent association of morbidity and mortality of human immunodeficiency virus (HIV)-infection with liver injury, the inclusion of drugs with anti-fibrotic activity to the treatment of people living with HIV (PLWH) is pathogenically important.Alcohol consumption is known to speed up liver fibrosis development.Previously, we have shown that the exposure of hepatocytes to HIV and ethanol metabolites causes high oxidative stress, impairs proteasomal and lysosomal functions leading to accumulation of HIV in these cells, which end-ups with apoptotic cell death and finally, promotes progression to liver fibrosis.
Research motivation
The combined exposure of hepatocytes to HIV and alcohol induces hepatotoxicity and pro-fibrotic activation of hepatic stellate cells (HSC).Since HIV replication in hepatocytes is abortive, it cannot be fully controlled by antiretroviral therapy (ART) and thus, to prevent liver fibrosis progression in alcohol-abused PLWH, ART should be combined with the drugs suppressing apoptosis without enhancing HIV DNA integration in hepatocytes and decreasing pro-fibrotic activation of liver nonparenchymal cells.
Research objectives
The objective of this study was to investigate whether obeticholic acid (OCA) prevents HIV/ethanol metabolism-induced activation of HSC by HIV+apoptotic hepatocyte engulfment, thereby diminishing liver fibrosis.
Research methods
The study was performed on hepatocyte-like Huh7.5-CYP (RLW) cells infected with HIV ADA and exposed to acetaldehyde-generating system (AGS) in the presence or absence of OCA.As an end-point, we have measured expression of HIV-related markers (HIVgagRNA-by real-time polymerase chain reaction (PCR), p24- by western blot, HIV DNA-by semi-nested PCR, integrated HIV DNA-by ddPCR) and non-HIVrelated parameters (lysosomal and proteasomal activities, hepatocytes apoptosis).We also characterized pro-fibrotic activation and inflammasome induction in HSC (LX2 cells) by HIV-containing apoptotic hepatocytes internalization.
Research results
We found that OCA attenuated HIV-AGS-induced accumulation of HIVgagRNA, HIV DNA and p24.It suppressed ROS production, restored chymotrypsin-like proteasome activity as well as cathepsin B lysosome activity and decreased apoptosis in RLW cells.Exposure of HIV-containing apoptotic hepatocytes to OCA prevented activation of inflammasome and pro-fibrotic activation of HSC.
Research conclusions
By suppressing oxidative stress and restoring proteasomal and lysosomal functions impaired by HIV and ethanol metabolism, OCA decreases accumulation of HIV in hepatocytes, leading to down-regulation of apoptosis in these cells.OCA also reverses pro-fibrotic and inflammasome-related activation of HSC triggered by engulfment of HIV-containing apoptotic hepatocytes, potentially contributing to suppression of liver fibrosis development.
Research perspectives
Ourin vitrostudies are in the frame of pre-clinical characterization of anti-fibrotic effects of OCA in alcohol-exposed PLWH with a high risk of liver fibrosis development.At the next step, thesein vitroeffects will be confirmed byin vivoexperiments on humanized mice infected with HIV and fed ethanol diet.
 World Journal of Hepatology2020年11期
World Journal of Hepatology2020年11期
- World Journal of Hepatology的其它文章
- Malnutrition in cirrhosis: More food for thought
- Paraneoplastic syndromes in cholangiocarcinoma
- Noninvasive scores for the prediction of esophageal varices and risk stratification in patients with cirrhosis
- Natremia and liver transplantation: The right amount of salt for a good recipe
- Inhibition of vascular adhesion protein-1 modifies hepatic steatosis in vitro and in vivo
- Aceclofenac-induced hepatotoxicity: An ameliorative effect of Terminalia bellirica fruit and ellagic acid