
雌雄构树幼苗光合能力对干旱胁迫响应的性别差异
2021-01-05 02:53苟蓉游明鸿白史且刘金平曾晓琳
四川林业科技 2020年6期
苟蓉, 游明鸿, 白史且, 刘金平*, 曾晓琳
1. 西华师范大学西南野生动植物资源保护教育部重点实验室,四川 南充 637009;
2. 四川省草原科学研究院,四川 成都 611731
雌雄异株植物受性别分株影响,维持种群稳定性的能力较弱[1],雌雄个体对生境胁迫的适应和抵抗能力,决定着个体存活、种群特征、物种延续及生态系统稳定性。诸多研究说明,雌雄个体对干旱、养分缺乏、重金属及盐胁迫等环境胁迫,表现出显著的抗性性别差异[2-5]。多数雌雄异株植物开花之前很难分辨性别,依株高、叶型、叶色等外部形态辨别的准确度不高,依易受环境因素及植物发育状况影响的生理指标鉴定的可靠性较差[6,7],依染色体组型和同工酶鉴定的局限性明显[8,9],依特异蛋白质和分子标记鉴定的可信度高,但鉴定程序复杂、成本高、耗时长,难以大范围推广,故目前关于雌雄异株植物抗性研究多是在成年植株上完成。幼苗是生活史中最敏感、最脆弱阶段,幼苗对生境胁迫的适应性及抗性性别差异,对种群密度、年龄结构、性比、出生率和死亡率及空间格局等种群特征影响深远。故开展幼苗抗性研究,对雌雄异株植物个体发育、种群形成研究及濒危度评价有指导价值,对该类植物保护、开发和利用有重要意义。
构 树(Broussonetia papyrifera) 为 桑 科(Moraceae)构树属(Broussonetia)雌雄异株植物,具有材用、饲用、药用、食用等开发价值。关于其形态特征、群落分布、化学成分、饲用及药用价值[10-13],及胁迫下的适应性和抗性已进行了大量研究[14-18],但诸多研究未区分雌雄株进行,未关注雌雄个体生长、生理、生殖及抗性等方面的性别差异。遗传特性、生理特化、环境因子及生殖成本共同作用,导致了雌雄个体适应性和抗逆性差异[3,5]。通过叶片特征[19]和同工酶[20]分析,初步表明雌雄构树形态结构和生理代谢有差异,关于其生长、生理、生殖及抗性基础的性别差异研究极少。光合作用是决定植物生产性能和抵御能力的物质与能量根本来源,干旱是植物幼苗期最易遭受的生境胁迫,幼苗光合作用对干旱胁迫的响应速度和应对策略,对植株存活与否、生长速度及抗性强弱起决定性作用。
干旱对光合作用的影响,体现在光合叶面积、光合色素、光合参数及同化效益等方面。本研究以构树雌雄扦插幼苗为材料,经60 d 干旱处理后,通过测定叶片数、叶长、叶宽、叶厚、叶面积等叶性状,测定Chla、Chlb 和 Caro 等色素含量,及Pn、Gs、Tr、Ci等光合参数,分析干旱下雌雄株光合叶面积、光合色素组成及光合速率变化规律,研究干旱胁迫对构树幼苗光合能力影响的性别差异。以期为构树品种选育、栽培养护、开发利用及资源保护提供依据,为雌雄异株植物的开发、利用和保护提供借鉴。
1 材料与方法
1.1 试验材料及设计
于2019 年4 月,在上年标记性别的中龄雌雄构树上,剪取半木质化枝条,选取长3±0.2 cm,粗0.5±0.1 cm,含1 芽、芽距两端各约1.5 cm、重量0.65±0.01 g 的插穗,生根剂处理后,扦插于以沙土:紫色土:腐殖土1∶1∶1 为基质,高40 cm、口径30 cm 塑料花盆中。每盆同性别扦插5 株,雌雄各12 盆,置相对湿度60%、光照16 h·d−1、28 ℃的培养箱,期间保持土壤湿润,育苗培养40 d 后,移出培养箱。
在室温自然蒸发下,以1 018 mL·盆−1(饱和持水量)浇水48 h、72 h、144 h 和192 h 后,测得土壤水分(土壤含水量测定仪TDR300)为饱和含水量的62.15%、48.33%、32.62%和21.89%,为水分充足(CK)、轻度干旱、中度干旱、重度干旱4 个水分梯度。每性别随机3 盆为1 组,分为4 组。分别进行干旱处理。
1.2 测定项目与方法
叶的形状:每组随机选择9 株,测叶片数,自上而下测第3 叶片的叶长、叶宽、叶厚(YH-1 型叶厚仪)、单叶面积(YMJ-C 型叶面积测定仪)和株总叶面积。
叶绿素含量采用乙醇丙酮混合提取法[21]。用LI-6400 便携式光合仪,光照强度1300lx 下测现实光合参数,在有效辐射为850 μmol·m−2·s−1(光响应曲线表明该光照下Pn 最大)的内置光源下测潜在光合速率,于第3 叶片中部随机3 点连续采集3 个值。
1.3 数据预处理与处理
用SPSS 19.0 软件对所有数据进行方差分析,并用Duncan 法对各参数进行显著性检验(SNK 检验)。
2 结果与分析
2.1 干旱对叶性状影响的性别差异
干旱显著影响幼苗的叶数、叶长、叶宽、叶厚、单叶面积和总叶面积(P<0.05)(见表1)。与水分充足(CK)比较,轻度干旱显著降低叶长和总叶面积,中度干旱显著降低叶宽、单叶面积而增加叶厚,重度干旱则显著降低叶数(P<0.05),使重度干旱下叶数、叶长、叶宽、单叶面积与总叶面积显著低于其他处理(P<0.05),而叶厚最厚。4 个水分下,雌株叶长与叶厚大于雄株,叶数和总叶面积小于雄株(P<0.05),而叶宽和单叶面积无性别差异。
方差分析F值表明,干旱度间叶性状差异大于性别间 (见表2),干旱影响大小为叶长>总叶面积>单叶面积>叶宽>叶厚>叶数;性别影响大小为叶长>叶数>总叶面积>叶厚>叶宽>单叶面积。干旱胁迫使叶长和叶宽、单叶面积和总叶面积表现出极显著性别差异(P<0.01),使叶数和叶厚表现出显著性别差异(P<0.05)。雌株主要通过降低叶数、增加叶厚应对干旱,雄株主要通过降低叶长与单叶面积应对干旱,干旱度越严重雄株光合总叶面积越显著大于雌株。
2.2 干旱对光合色素影响的性别差异
干旱对Chla、Chlb、Caro、Chla+b 和Chla/b 均有显著影响(P<0.05)(见表3)。3 个干旱处理Chla、Chla+b 和Chla/b 显著大于水分充足(CK),轻度干旱下Chlb 达最大值,中度干旱下Chla 和Chla+b 达最大值,重度干旱下Caro 和Chla/b 达最大值。Chla、Chlb、Chla+b 随干旱度增大显著先增后降,Caro 和和Chla/b 随干旱度增大不断增加。轻、中度干旱间Chlb 无显著差异,中重度干旱间Caro 和Chla/b 无显著差异(P>0.05)。4 个水分下,雌株Chla 和Chla+b大于雄株(P<0.05),而Chlb、Caro 和Chla/b 无性别差异(P>0.05)。
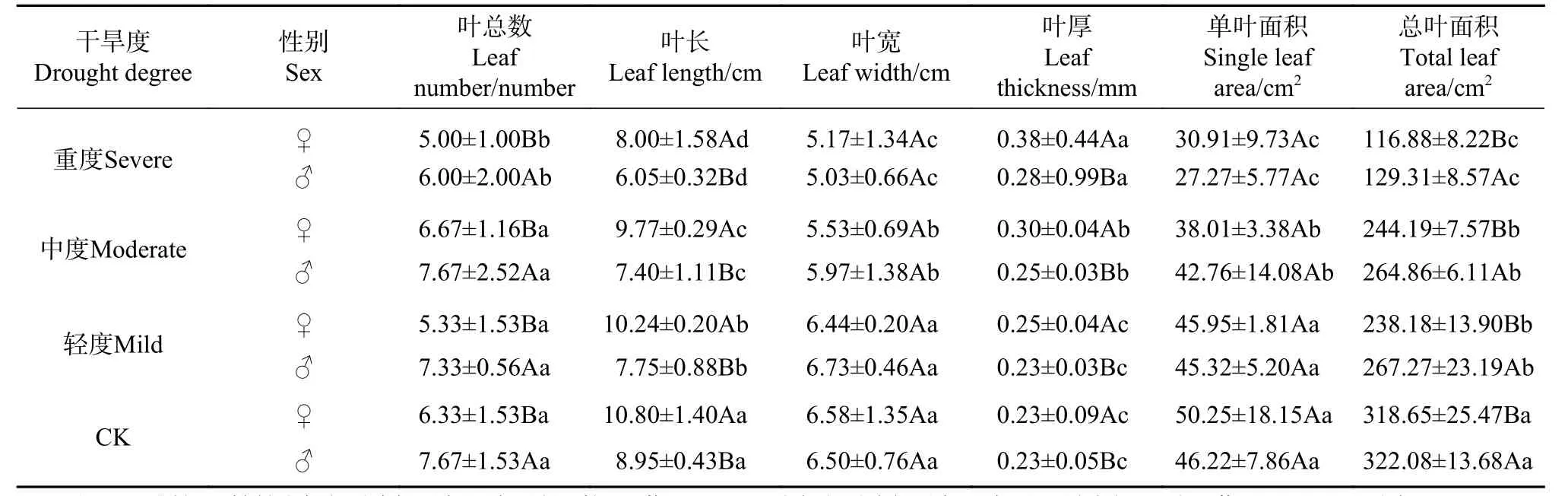
表 1 干旱对叶性状影响的SNK 检验Tab. 1 SNK test of leaf traits under different drought degrees
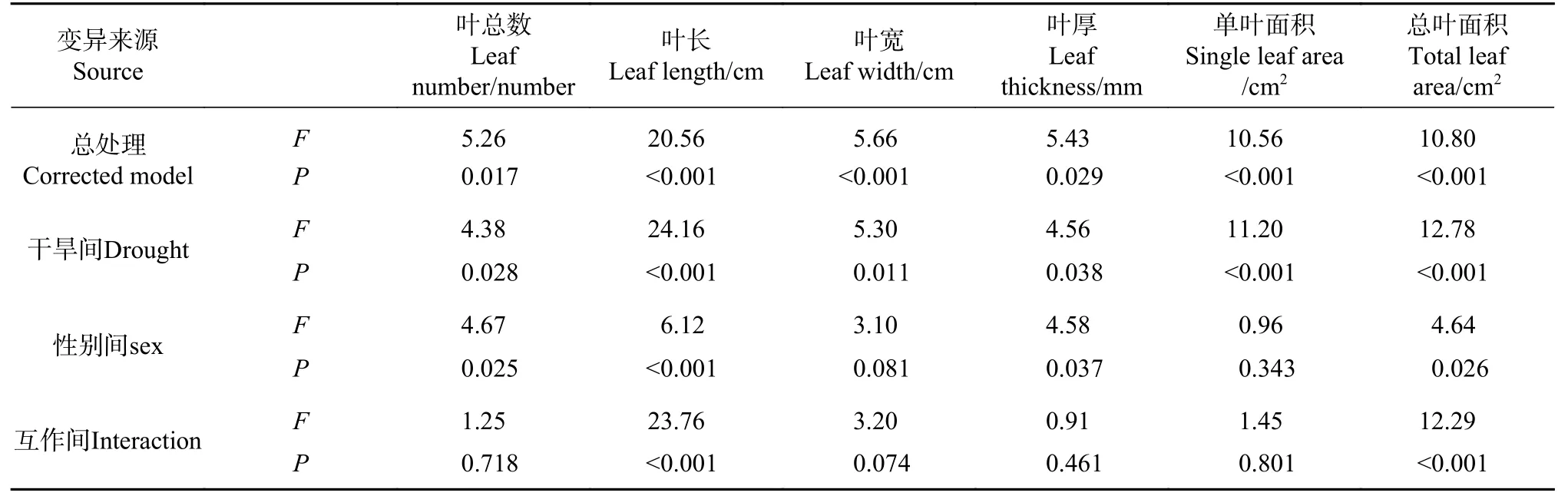
表 2 叶性状差异的双因素方差分析Tab. 2 Two-factor variance analysis about differences of leaf traits
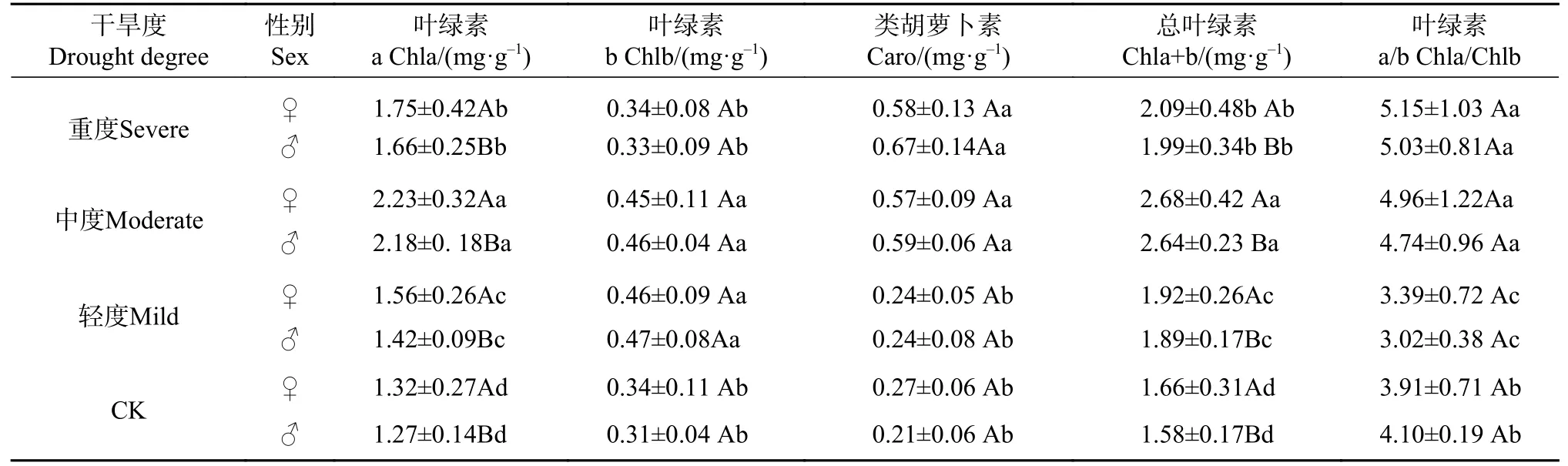
表 3 干旱对光合色素含量影响的SNK 检验Tab. 3 SNK test of photosynthetic pigment content under different drought degrees
方差分析F值表明,干旱度间光合色素差异大于性别间 (见表4),干旱对色素影响大小为Caro>Chla+b>Chla>Chlb>Chla/b,性别对色素影响为Chla>Chla+b>Chlb>Chla/b>Caro。干旱胁迫极显著改变了Chla、Caro 和Chla+b 的性别差异(P<0.01),显著改变了Chlb 和Chla/b 的性别差异(P<0.05)。干旱度越重雄株Chla 和Chla+b 越大于雌株,而Caro 越小于雌株。干旱是影响色素含量的主要因子,雌雄株均通过提高Caro 和Chla/b 应对干旱。
2.3 干旱对光合参数的性别差异
干 旱 对Pn、Gs、Tr、Ci和PPn有均显著影响(P<0.05)(见表5)。轻度干旱Pn、Gs、Tr、Ci均大于CK(P<0.05),中度干旱Tr 和Ci 达最大值 而Pn和Gs低于轻度干旱,重度干旱Pn、Gs、Tr、Ci均低于中度干旱(P<0.05)。轻中度干旱对PPn无影响,重度干旱下PPn下降(P<0.05)。4 个水分下,雄株Pn、Gs、Tr、Ci均大于雌株,而PPn小于雌株。轻度干旱利于Pn和Gs,中度干旱利于Tr和Ci,重度干旱造成光合系统破坏使Pn和PPn最低。
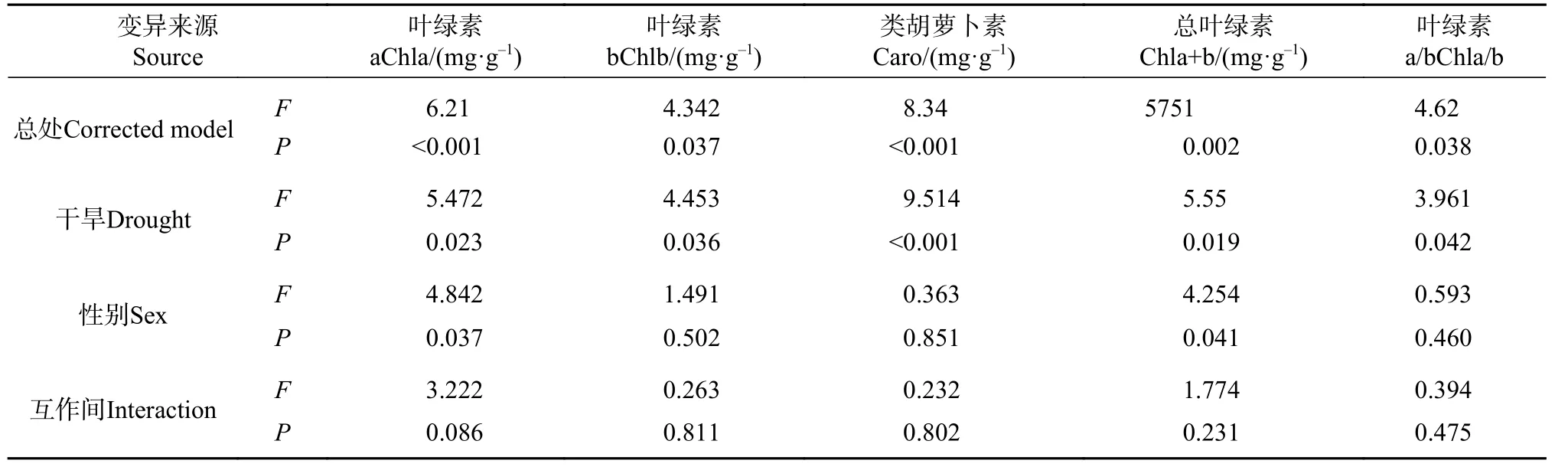
表 4 光合色素含量差异的双因子方差分析Tab. 4 Two-factor variance analysis about differences of photosynthetic pigment content
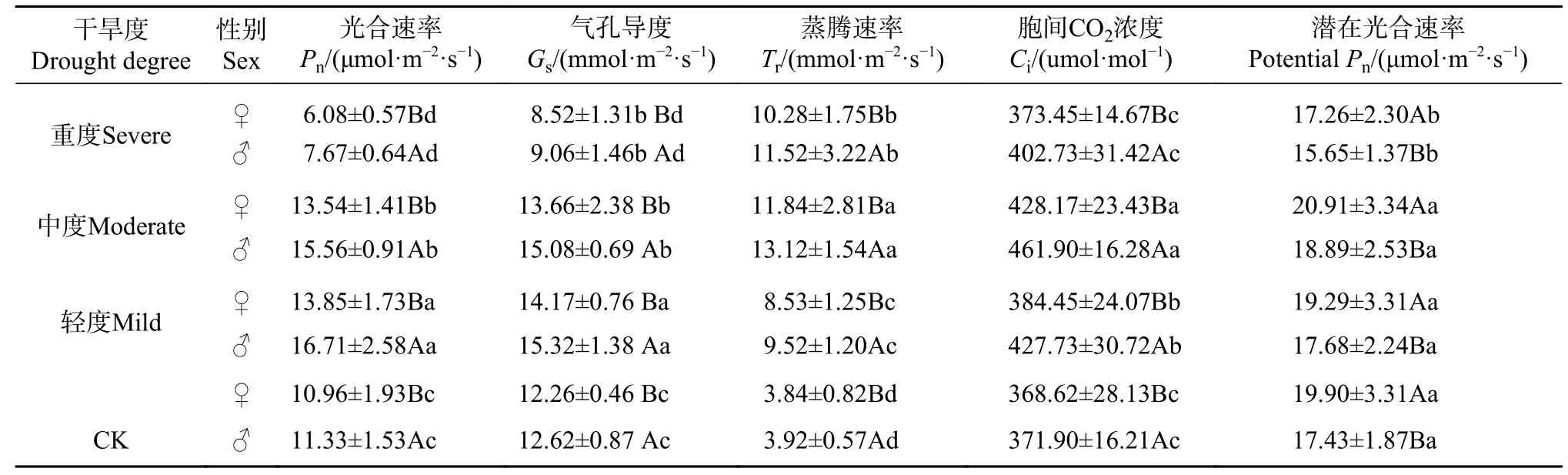
表 5 干旱对光合参数影响的SNK 检验Tab. 5 SNK test of photosynthetic parameters under different drought degrees
方差分析F值表明,干旱度间光合参数差异大于性别间 (见表6),干旱对色素参数影响大小为Tr>Pn>Gs>Ci> PPn,性别影响为Pn>Ci>PPn>Gs>Tr。干旱胁迫极显著增大或缩小了Pn、Gs、Tr、Ci的性别差异,显著缩小了PPn的性别差异。干旱与性别互作极显著影响Pn和Tr,显著影响Ci。
3 讨论与结论
诸多研究表明,植物通过降低叶片位置、减少叶片数和叶面积、停止或延缓分蘖数、缩短节间长和茎长、增加根长和根数等构件调节方式应对干旱[22-24],本试验中构树雌雄幼苗均通过降低株高、叶片数、叶长、叶宽、单叶面积和总叶面积,增加叶厚、茎粗和根长应对干旱胁迫,与其他植物研究结果相似。干旱下叶性状变化表现出显著性别差异,雌株通过降低叶片数,雄株通过缩小叶长、叶宽及叶面积应对干旱,虽雌雄株叶性状适应策略不同,结果都减少总叶面积来抑制蒸腾水分消耗作用。雄株叶片表皮厚度、栅栏组织厚度、主脉维管束木质部厚度及维管束厚度均显著大于雌株叶片[25],与本研究雄株叶片厚度大于雌株一致。雄株叶片结构使其具有更高的水分储存能力和更强的水分输送能力,故干旱下雄株比雌株保留了更多的总叶面积,使其光合叶面积优于雌株。干旱影响分生组织和幼嫩组织发育,叶面积减少是应答缺水早期的表现,同时幼叶向老叶吸水,加速老叶枯萎死亡[26]。试验发现,干旱下雌株单叶面积未减少而老叶衰老,雄株单叶面积缩小而老叶健康,表明在老叶与新叶水分整合也有性别差异。
构树常通过缺刻数、缺刻裂度和表面绒毛密度,形成与生境相适应的光合叶面积。野外调查发现,种群间、个体间、枝条间及叶片位置间,缺刻数和缺刻裂度有明显差异。缺刻不受性别影响,且与植株或枝条年龄无关,干旱生境下幼苗常有浅裂出现。本试验在光照柔和、空气湿度相对稳定的室内完成,幼苗叶片均无缺刻出现。缺刻由不完全显性基因控制[27],或叶片分化时遭遇的空气湿度和光照强度,是引起叶片缺刻性状差异的主要原因。缺刻不仅减少单叶面积,改变叶片水分运输、散失与利用效率,同时可提高树冠透光率与空气流通性,从而影响整株的光合能力。探究叶片缺刻的遗传基础及生境影响机理,对认知构树光合叶面积调节能力具有重要意义。
植物常依据生境条件,改变吸收和传递光量子的光合色素含量和比例,维持正常光合所需,同时防止过剩光能的过量积累[28]。干旱胁迫下光合色素变化可反映植物对干旱的敏感度和抵御干旱的能力[29]。诸多研究表明,干旱胁迫使叶绿素合成受阻,降解加快,导致随干旱度加重叶绿素含量迅速下降[30-31],其中Chla/b 对干旱更为敏感降低更快[32]。本试验中,轻中度干旱Chla、Chlb、Chla+b 含量显著增加,重度干旱虽降低叶绿素含量,但Chla 和Chla+b 含量大于水分充足时,其中Chla 比Chlb 更易受干旱影响。Chla/b 随干旱增加而显著增加,表明构树为抗旱能力较强的阳生树种。Caro 具有保护叶绿素及抗氧化和延缓衰老的作用,常把Caro 增加当作植物受伤或衰老的表现,干旱下Caro 与叶绿素含量并线性增减,或许Caro 具有更多未知功能。雌雄株均通过提高Caro 和Chla/b 应对干旱,但干旱越严重雌株Chla 和Chla+b 含量越大于雄株,说明雌株吸收、传递和转化光能的色素基础强于雄株。
短期或轻度干旱使植物叶片水势降低、气孔关闭、降低CO2摄取量使光合作用降低[33],构树幼苗在轻度干旱下Pn、Gs、Tr和Ci均显著大于CK,或因叶面积缩小使叶绿体密度和光合色素含量增加,或因轻微水势降低激活了光合相关酶的活性所致。植物通过关闭气孔或降低光合器官活性应对干旱[34],中度干旱显著降低了Gs,虽显著增加了叶绿素含量和Ci,因系统活性降低先抑制了Pn。严重干旱导致光合色素分解,影响光能吸收和转化效率等[35],对幼苗光合系统造成不可逆的伤害,致使光合参数显著降低。有研究认为,雌雄异株植物光合指标在水分充足时无性别差异,但也有研究证明雌雄异株植物光合指标一直存在性别差异[3]。构树幼苗光合面积、光合色素和光合参数表现出显著的性别差异,雌株Chla 与Chla+b 大于雄株,干旱使Chlb无性别差异的降低,使雌株Chla/b 增大而影响电子传递效率,故雌株Pn小于雄株。本试验4 个干旱条件下,雌株光合色素含量大于雄株,而Pn均小于雄株,但PPn则大于雄株。仅因光源不同使雌雄株Pn和PPn表现大相径庭,是雌雄株光响应能力、光饱和点和表观量子数存在性别差异,还是因水分利用率性别差异,导致Pn和PPn呈相反表现,有待于深入研究。
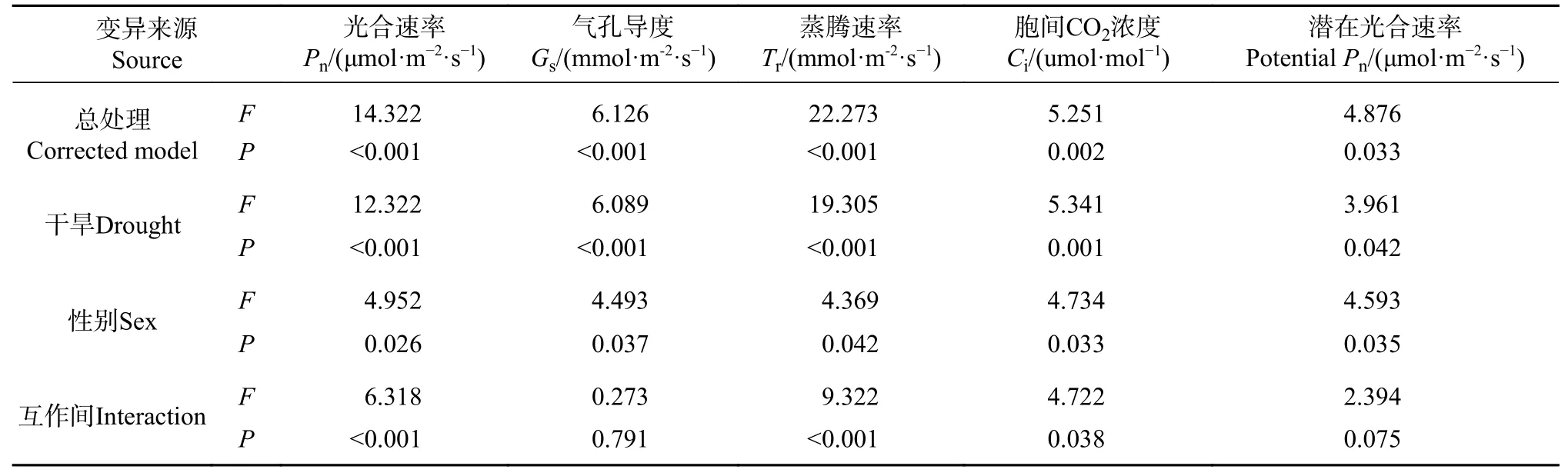
表 6 光合参数差异的双因子方差分析Tab. 6 Two-factor variance analysis about differences of photosynthetic parameters
猜你喜欢
安徽农业科学(2021年6期)2021-04-20
四川蚕业(2020年2期)2020-12-18
新疆农业科学(2020年1期)2020-02-14
绿色中国(2019年16期)2019-11-26
江苏农业学报(2018年5期)2018-09-10
现代农业研究(2017年10期)2018-01-02
中国扶贫(2017年16期)2017-08-16
中国扶贫(2017年5期)2017-03-10
江苏农业科学(2016年10期)2017-02-05
西南学林(2016年2期)2016-11-08
