
198份小麦种质资源赤霉病综合抗性鉴定及其FHB1抗性基因检测
2021-01-02 17:05贾宝森徐锐熊泽浩高德荣王书平王晓玲方正武
江苏农业科学 2021年23期
贾宝森 徐锐 熊泽浩 高德荣 王书平 王晓玲 方正武
摘要:选取26份小麦地方品种,75份黄淮麦区小麦改良品种,97份长江中下游麦区小麦改良品种,采用单花滴注法进行赤霉病抗性鉴定并调查供试品种的穗长、株高、千粒质量和小穗密度等农艺性状。结果表明,鉴定筛选到抗病品种2个,中抗品种(系)35个,中感品种(系)77个,高感赤霉病的品种(系)84个。利用FHB1分子标记对供试小麦进行检测。其中在Fhb1位点上表现为抗病性基因型(Fhb1+)的小麦品种(系)33个,在Fhb1位点上表现为感病性基因型(Fhb1-)小麦品种(系)165个,且呈抗病基因型品种与感病基因型品种的平均病小穗率差异显著(P<0.05)。FHB1分子标记检测结果为阳性的小麦品种赤霉病抗性显著高于检查结果为阴性的小麦品种。此外,筛选得到抗性达到中感及以上水平的小麦品种(系) 114个,考察农艺性状发现扬麦18、宁麦19、宁麦8号、扬麦12、等18个品种(系)农艺性状优良,可作为小麦赤霉病育种的亲本选用。
关键词:小麦;赤霉病;农艺性状;Fhb1抗病基因
中图分类号: S512.102.4 文献标志码: A
文章编号:1002-1302(2021)23-0104-05
收稿日期:2021-04-01
基金项目:湖北省科学技术重大创新专项(编号:2018ABA085)。
作者简介:贾宝森(1997—),男,山东济宁人,硕士研究生,主要从事麦类种质资源创新与利用研究。E-mail:jbs1010813546@126.com。
通信作者:方正武,博士,教授,主要从事麦类种质资源创新与利用研究。E-mail:fangzhengwu88@163.com。
小麦赤霉病(Fusarium head blight,简称FHB)是一种由禾谷镰刀菌侵染导致的真菌型病害,是目前我国小麦生产上最严重、最普遍、危害最大的小麦穗部病害之一。禾谷镰刀菌产生的子囊孢子在小麦开花期侵染穗部颖壳,病菌侵染后在小麦籽粒生长发育过程中不断增殖生长,并在小麦籽粒中产生和积聚脱氧雪腐镰刀菌烯醇(DON)、玉米赤霉烯酮(ZEN)等多种危害人畜健康的毒素[1-2],严重威胁粮食安全和小麦生产质量水平的进一步提高。长江中下游麦区、黄淮麦区的小麦种植面积约占全国小麦种植总面积的2/3,是我国重要的小麦生产区。因气候原因长江中下游麦区长久以来都是小麦赤霉病的高发区、重发区。近年来,江苏省年均赤霉病发生面积已超该省小麦种植总面积的50%,高达120万hm2[3]。随着农作制度改变和气候变化等因素的影响,小麦赤霉病迅速由长江中下游麦区向黄淮麦区蔓延,病害面积日渐扩大并有加重的趋势。培育并推广农艺性状优良且兼顾抗赤性的小麦品种是减轻小麦赤霉病危害最经济、安全、有效的途径[4]。长江中下游麦区为赤霉病抗病性育种提供了丰富的种质资源,其中苏麦3号、部分扬麦系列和宁麦系列等都是小麦抗赤霉病育种中常用的抗源材料[5-6]。此外,望水白、海盐种、黄方柱和白三月黄等抗性优良的地方品种同样源于长江中下游麦区[7]。黄淮麦区历史上不是赤霉病的常发区,对抗赤霉病品种的关注较少,迄今为止黄淮麦区优质抗源稀缺,该地区尚未发现有效稳定的赤霉病抗源[8]。发现和利用抗赤霉病小麦种质资源,提高赤霉病抗性已成为黄淮麦区主要育种目标之一。
小麦赤霉病抗性可分为抗初侵染(TypeⅠ)、抗拓展(TypeⅡ)、抗籽粒侵染(TypeⅢ)、耐病性(TypeⅣ)、抗DON积累(TypeⅤ)5种抗病类型[9]。小麦赤霉病抗性是由多基因控制的数量性状,目前已定位上百个与赤霉病抗性相关的数量性状基因座(QTL)[10],但仅有7个被正式命名,迄今为止只有Fhb1和Fhb7相继被克隆[11-12],研究表明来源于望水白的FHB1是目前抗性最强、最稳定且应用也最为广泛的抗病基因。FHB1不仅可以显著降低赤霉病严重度还能将由禾谷镰刀菌代谢产生的DON转化为低毒的脱氧雪腐镰刀菌烯醇-3-葡萄糖苷(DON-3G),减轻毒素积聚的危害[13-14]。随着我国小麦赤霉病的发生日渐严重,近年来有关赤霉病抗性鉴定的研究日益增多。徐乔乔等通过单花滴注接种法对45份小麦品种(系)进行了抗性鉴定,获得了13份抗病材料[15]。靳凤从363份美国冬小麦核心种质材料中,发现22份材料表现出较好的赤霉病抗性并且其中一部分材料在Fhb1位点上表现为感病性基因型[16]。周淼平等通过对黄淮麦区的500多份小麦材料进行赤霉病抗性鉴定,获得抗性水平达中感以上的小麦品种(系)共18份[17]。这些鉴定结果极大地丰富了抗赤霉病品种选育的种质信息。本研究采用单花滴注法对198份小麦材料进行赤霉病抗性鉴定并进行农艺性状的调查。按NY/T 2954—2016《小麦区域试验品种抗赤霉病鉴定技术规程》标准评价供试小麦抗性水平。采用Fhb1 STS功能标记,对供试材料进行Fhb1等位基因检测。筛选获得抗赤性较好且兼顾丰产性的小麦种质资源,为小麦抗赤霉病育种的亲本选择提供依据。
1 材料与方法
1.1 材料与试验设计
供试材料来源于黄淮麦区、长江中下游麦区的198个小麦品种(系)。其中地方品种26个,黄淮麦区改良品种75个,长江中下游麦区改良品种97个。试验设苏麦3号、扬麦158、淮麦20、周麦18分别为赤霉病高抗、中抗、中感、高感对照品种。
1.2 抗病鉴定圃设置
试验于2018—2019年小麦生长期,在湖北省荆州市荆州区长江大学农学院试验基地进行。每个材料种植2行,行长1.5 m,行距0.2 m,株距0.1 m,2次重复,随机区组排列。试验田的田间管理同常规育种田。
1.3 赤霉病抗性鉴定
于2019年4月上旬,在田间进行接种鉴定。参照Yu等方法制备禾谷镰刀菌孢子悬浮液,悬浮液浓度为5×105个/mL[18]。于小麦扬花始期使用微量移液器进行单花滴注接种:每个品种选取20个开花期基本相同的单穗,每穗选中部1个小花注入 10 μL 孢子悬浮液,用彩色胶带对接种穗进行标记,并记录接种日期,接种后进行弥雾保湿。接种21 d后调查病小穗数,计算病小穗率。按NY/T 2954—2016《小麦区域试验品种抗赤霉病鉴定技术规程》标准划分抗性水平和小麥赤霉病严重度。
1.4 Fhb1基因的检测
每个品种选取20粒健康饱满的种子种植于发芽盒中,在温室中催芽出苗,每个品种(系)分别取 5~10张幼嫩叶片,在液氮中速冻并研磨至粉末,采用改良十六烷基三甲基溴化铵(CTAB)法[19]提取小麦基因组DNA。并用NanoDrop 2000微量分光光度计和琼脂糖凝胶电泳测定DNA浓度和纯度,于 -20 ℃ 条件下保存待用。
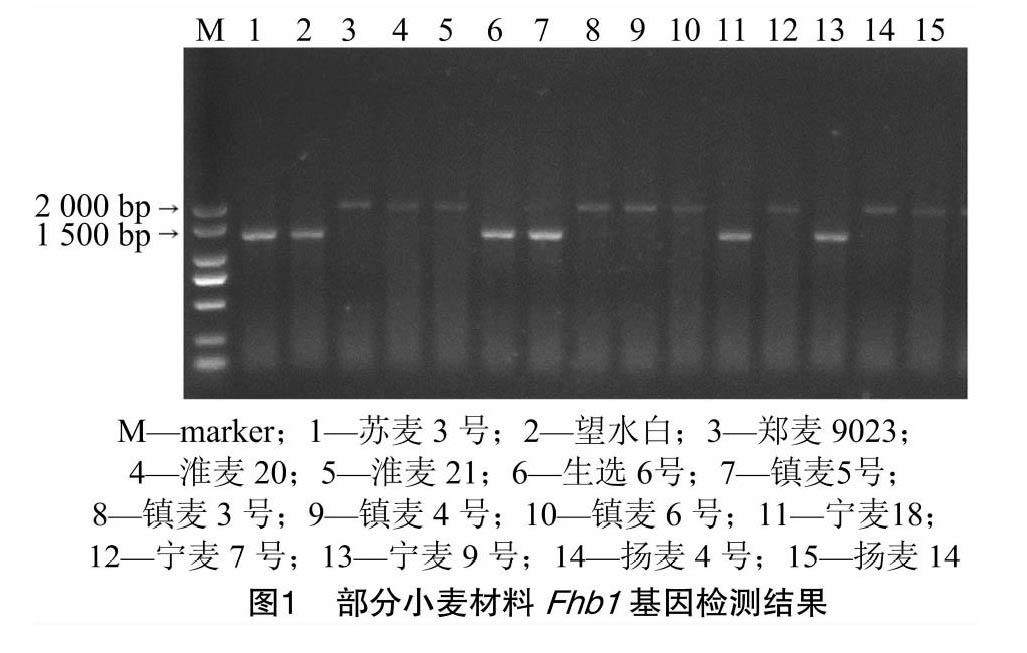
参照李腾公布的引物序列[20](TaHRC-STS-F:ATTCCTACTAGCCGCCTGGT,TaHRC-STS-R:ACTGGGCAAGCAAACATTG)进行扩增,PCR体系和扩增程序参考李腾的方法[20]。用1.5%琼脂糖凝胶电泳检测PCR产物。Fhb1抗病等位基因(记为Fhb1+)、Fhb1感病等位基因(Fhb1-)扩增片段分别为1.4 kb和 2.0 kb。
1.5 农艺性状的测定
2019年,待各品种成熟期后,每份材料每次重复取10株主茎穗,测量记录穗长、小穗数并计算小穗密度(每穗小穗数/穗长)。株高:每个品种随机选取10个单株,测量主茎高度取平均值;晾干脱粒后使用浙江云普智能考种仪测量小麦籽粒粒长、粒宽、穗粒数和千粒质量等经济性状。
1.6 统计分析
利用Microsoft Excel 2019和Sigmaplot 10.0对试验获得的8个农艺性状表型值、病小穗率、赤霉病严重度等数据进行描述性统计分析。
2 结果与分析
2.1 供试小麦品种(系)的赤霉病抗性
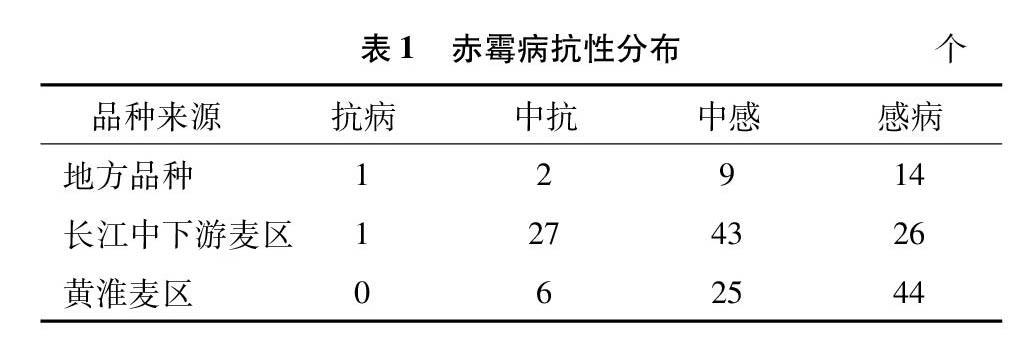
采用单花滴注接种法对198份供试小麦品种(系)进行抗性鉴定,赤霉病抗性鉴定结果见表1。单花滴注鉴定法共筛选到赤霉病抗性达高抗的品种(系)2个,分别是苏麦3号、望水白,占1.01%;中抗35个,占17.68%,其中长江中下游麦区27个,黄淮麦区6个,地方品种2个;中感赤霉病品种(系)77个,占38.9%,其中长江中下游麦区43个,黄淮麦区25个,地方品种9个。高感品种(系)84个,占42.4%,其中黄淮麦区44个,长江中下游麦区26个,地方品种14个。苏麦3号、扬麦158、淮麦20、周麦18平均病小穗率分别为7.4%、22.4%、29.0%、54.9%。
2.2 FHB1基因的分子标记检测结果
利用FHB1的分子标记检测198份供试小麦。
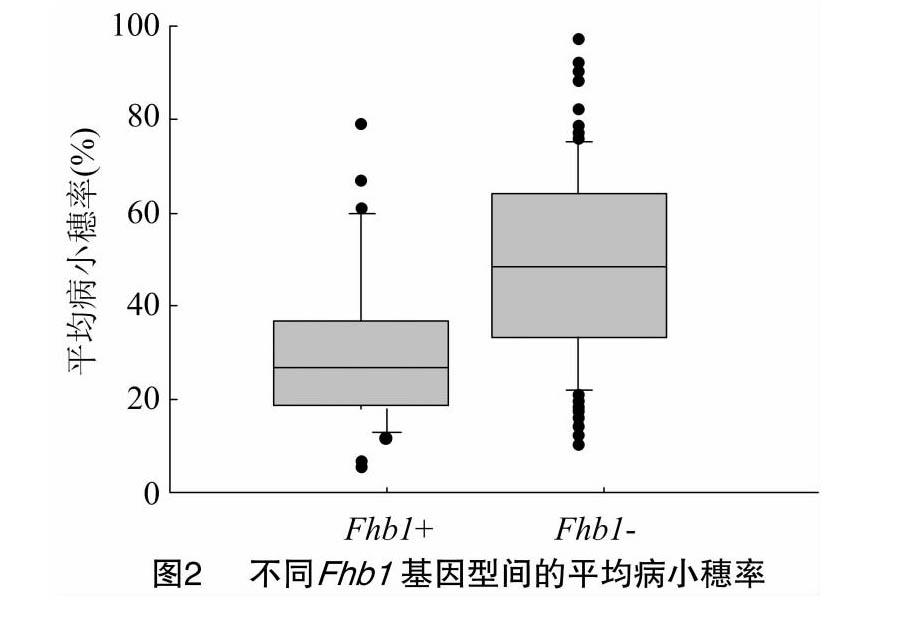
结果表明,33个材料在Fhb1位点呈抗病基因型Fhb1+。其中地方品种9个(白慈麦、白蒲、赤小麦、和蒲头、胡须麦、糯麦、望水白、紫皮、翻山小麦),黄淮麦区1个(淮麦40),长江中下游麦区23个(浙麦1号、宁麦9号、苏麦5号、扬麦18、镇麦5号、扬麦28、镇12096、宁麦13、宁麦14、宁麦15、扬辐3046、扬17G83、苏麦3号、扬16-157、鄂麦178、宁麦16、宁麦18、宁麦19、宁麦24、宁麦26、生选6号、镇麦8号、镇麦12号)。其他材料在该位点呈感病基因型Fhb1-。统计结果表明,呈抗病基因型Fhb1+与感病基因型Fhb1-品种的平均病小穗率差异显著(P<0.05)。FHB1分子标记检测结果为阳性的小麦品种赤霉病抗性显著高于检查结果为阴性的小麦品种,但有少数检测结果为阳性的品种表型为感病(图1、图2)。
2.3 抗赤霉病小麦品种(系)农艺性状与经济性状
经田间调查和统计分析供试品种(系)的相关农艺性状,农家种的千粒质量指标普遍偏低,除农家种之外的中抗品种(系)的千粒质量指标高于中感和高感品种;农家种的株高性状普遍偏高,黄淮麦区中抗品种(系)的株高高于中感和高感品种,而长江中下游麦区材料则表现相反; 所有品种(系)小穗密度与抗性呈负相关,且黄淮麦区品种(系)的小穗密度高于长江中下游麦区品种(系);其次,农家种的穗长性状高于改良品种,中抗和中感品种(系)的穗长明显高于高感品种;中抗品种的穗粒数低于中感和高感品种的穗粒数,同时黄淮麦区品种(系)穗粒数明显低于长江中下游麦区品种;其他各抗性等级、麦区小麦品种(系)间粒长、粒宽、小穗数等农艺性状均没有明显差异(图3)。
在抗性综合评价为中感及以上的品种(系)中,以农艺性状株高<90 cm,穗粒数>40粒/穗,小穗数>20个,小穗密度>1.7个/cm,千粒质量>35 g为优良的评测标准,结果筛选出扬麦19、扬麦22、扬16初74、扬糯麦1号、扬麦18、宁麦19、扬辐麦2号、宁麦8号、扬麦12、宁麦7号、扬14-214、宁麦18、鄂麦178、扬麦6号、镇麦168、扬麦17、扬麦20、扬辐麦9311、宁麦16、扬麦11号、扬麦16和皖麦32共22个品种(系),这些品种除皖麦32来自黄淮麦区外,其他品种均来自于长江中下游麦区(表2)。
3 讨论与结论
本研究采用单花滴注接种鉴定的方法,对198份供试小麦材料进行了赤霉病抗性的表型鉴定,仅有苏麦3号、望水白2个品种达到R级抗性,苏麦3号为改良品种,望水白则为农家种,且2个品种均来自长江中下游麦区;黄淮麦区未鉴定到赤霉病抗性水平达到R级的品种(系),说明R级赤霉病抗性的小麦种质资源十分稀少。同时,经过单花滴注接种鉴定到赤霉病抗性达到中抗的品种有35个,中感赤霉病的品种(系)为77个,其中大部分材料来源于长江中下游麦区,黄淮麦区供试材料赤霉病抗性普遍低于长江中下游麦区供试材料,说明黄淮麦区的赤霉病抗性品种匮乏,与相关研究结果[9,17]一致。农家种白慈麦、红壳酱以及黄淮麦区的郑麦9023、皖麦31、淮麦30、新乡289、皖麦32、淮麦40和长江中下游麦区的镇麦4号、扬麦11、宁麦24等小麦品种(系)赤霉病抗性鉴定为中抗赤霉,表明这些品种(系)的赤霉病抗性优良,可在赤霉病重发年份减轻赤霉病危害。程顺和等提出选用农艺性状优秀、抗病性优良的品种(系)杂交,后代兼顾产量与抗病性等性状选择,可达到选育出大面积推广种植中抗品种的目的[21]。但抗性优良的小麦品种(系)及农家种大多农艺性状较差,仅有2个R级抗性品种(苏麦3号、望水白)的株高在1.2 m以上。通过相关农艺性状分析,在单花滴注鉴定条件下,高抗和中抗小麦品种(系)的小穗数、穗长、小穗密度和株高等性状明显差于中感和感病品种(系)。改良品种中长江中下游麦区供试品种(系)的千粒质量、小穗密度、穗粒数等性状明显优于黄淮麦区材料,但株高较高。本研究在赤霉病抗性水平介于中感和中抗之间的品种(系)中筛选出性状优良的扬麦18、宁麦19、宁麦8号、扬麦12、宁麦7号、扬14-214、宁麦18、鄂麦178、镇麦168、扬麦6号、扬麦17、扬辐麦2号、扬麦20、宁麦16、扬辐麦9311、扬麦11号、扬麦16、皖麦32等共22个品种(系)可為赤霉病抗性育种的亲本选择。
Fhb1是目前抗性最强最稳定且应用最为广泛的抗赤霉病基因。本研究在198份供试小麦品种(系)中检测了Fhb1情况,共33个小麦品种(系)携带Fhb1基因,其中绝大部分材料来源自长江中下游麦区,而在黄淮麦区仅有淮麦40品种检测到携带有Fhb1基因,这可能与黄淮麦区抗赤霉育种时间较短和Fhb1基因与感秆锈病基因、某些不利农艺性状的连锁较为紧密[6,22]有关,因此在小麦育种进程中受到人工选择压力而被去除。目前长江中下游麦区主要通过导入Fhb1基因来改良小麦品种的赤霉病抗性,并由此育成大量品种[23]。苏麦3号、宁麦9号是我国小麦品种所含Fhb1基因的主要来源。长江中下游麦区抗性较好的宁麦系列品种和扬麦系列品种[7]就是利用宁麦9号培育成的。本试验鉴定结果的中抗材料郑麦9023、皖麦31、镇麦1号、扬辐麦2号等22个品种(系)并不携带Fhb1抗病基因。这可能是因为小麦赤霉病抗性为数量性状遗传,这些材料可能含有Fhb1以外的其他抗病基因。寧麦13、宁麦15、翻山小麦、鄂麦178等18份材料,虽携带Fhb1抗病基因,但赤霉病抗性水平均未达到中抗级别,可能是由于遗传背景不同,主效抗赤霉病基因与其他基因互作致使效应发生变化而导致赤霉病抗性改变或降低。
参考文献:
[1]甄阳光,柏 凡,张克英,等. 我国主要饲料原料及产品中呕吐毒素污染分布规律研究[J]. 中国畜牧杂志,2009,45(8):21-24,28.
[2]陆 彦,王科峰,殷 茵,等. 小麦赤霉病田间病情指数、病粒率及DON毒素含量之间的关系初探[J]. 农药科学与管理,2017,38(7):54-58.
[3]吴佳文,杨荣明,朱 凤,等. 2015年江苏省小麦赤霉病发生特点与防控对策探讨[J]. 中国植保导刊,2016,36(10):31-34,49.
[4]Bai G H,Shaner G.Management and resistance in wheat and barley to Fusarium head blight[J]. Annual Review of Phytopathology,2004,42:135-161.
[5]朱展望,徐登安,程顺和,等. 中国小麦品种抗赤霉病基因Fhb1的鉴定与溯源[J]. 作物学报,2018,44(4):473-482.
[6]蒋正宁,吕国锋,王 玲,等. 扬麦品种(系)赤霉病抗扩展性基因分子检测及其抗性评价[J]. 麦类作物学报,2019,39(12):1406-1415.
[7]Yu J B,Bai G H,Cai S B,et al. New Fusarium head blight-resistant sources from Asian wheat germplasm[J]. Crop Science,2008,48(3):1090-1097.
[8]张 煜,李正玲,王 震,等. 黄淮南部麦区小麦赤霉病抗性鉴定及基因型分析[J]. 麦类作物学报,2020,40(3):270-277.
[9]Mesterházy A.Types and components of resistance to Fusarium head blight of wheat[J]. Plant Breeding,1995,114(5):377-386.
[10]Jiang Y,Schulthess A W,Rodemann B,et al. Validating the prediction accuracies of marker-assisted and genomic selection of Fusarium head blight resistance in wheat using an independent sample[J]. Theoretical and Applied Genetics,2017,130(3):471-482.
[11]Lagudah E S,Krattinger S G.A new player contributing to durable Fusarium resistance[J]. Nature Genetics,2019,51(7):1070-1071.
[12]Wang H W,Sun S L,Ge W Y,et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat[J]. Science,2020,368(6493):eaba5435.
[13]Horevaj P,Brown-Guedira G,Milus E A.Resistance in winter wheat lines to deoxynivalenol applied into florets at flowering stage and tolerance to phytotoxic effects on yield[J]. Plant Pathology,2012,61(5):925-933.
[14]Pumphrey M O,Bernardo R,Anderson J A.Validating the Fhb1 QTL for Fusarium head blight resistance in near-isogenic wheat lines developed from breeding populations[J]. Crop Science,2007,47(1):200-206.
[15]徐乔乔,李巧云,牛吉山,等. 45份小麦种质赤霉病抗性评价与农艺性状分析[J]. 麦类作物学报,2019,39(4):393-399.
[16]靳 凤. 美国冬小麦核心种质赤霉病抗性鉴定及赤霉病抗性全基因组关联分析[D]. 杨凌:西北农林科技大学,2014.
[17]周淼平,姚金保,张平平,等. 黄淮麦区小麦抗赤霉病新种质的创制和筛选[J]. 麦类作物学报,2018,38(3):268-274.
[18]Yu J B,Bai G H,Cai S B,et al. Marker-assisted characterization of Asian wheat lines for resistance to Fusarium head blight[J]. Theoretical and Applied Genetics,2006,113(2):308-320.
[19]Saghai-Maroof M A,Soliman K M,Jorgensen R A,et al. Ribosomal DNA spacer-length polymorphisms in barley:mendelian inheritance,chromosomal location,and population dynamics[J]. Proceedings of the National Academy of Sciences of the United States of America,1984,81(24):8014-8018.
[20]李 腾. Fhb1基因对黄淮冬麦區小麦品种赤霉病抗性和主要农艺性状的影响[D]. 北京:中国农业科学院,2019.
[21]程顺和,张 勇,张伯桥,等. 小麦抗赤霉病育种2条技术路线的探讨[J]. 扬州大学学报,2003,24(1):59-62.
[22]He X Y,Singh P K,Dreisigacker S,et al. Dwarfing genes Rht-B1b and Rht-D1b are associated with both type Ⅰ FHB susceptibility and low anther extrusion in two bread wheat populations[J]. PLoS One,11(9):e0162499.
[23]张宏军,宿振起,柏贵华,等. 利用Fhb1基因功能标记选择提高黄淮冬麦区小麦品种对赤霉病的抗性[J]. 作物学报,2018,44(4):505-511.
猜你喜欢
今日农业(2022年16期)2022-11-09
天津农林科技(2022年2期)2022-04-19
今日农业(2021年9期)2021-11-26
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
作文小学中年级(2020年4期)2020-06-11
农药科学与管理(2019年5期)2019-08-13
中国中药杂志(2016年20期)2016-11-19
