
LED单色光对兔行为及同期发情影响的机理研究
2020-12-29 05:12潘孝青王杏龙
浙江农业学报 2020年12期
潘孝青,王杏龙,杨 杰,邵 乐,秦 枫,李 健,张 霞,翟 频
(1.扬州大学 动物科学与技术学院,江苏 扬州 225009; 2.江苏省农业科学院 畜牧研究所,江苏 南京 210014)
随着规模化养兔业的兴起,兔舍小气候的控制愈加显得重要。光照是影响家畜生长、繁殖的主要环境因素之一,合理的光照对提高家兔生产力、控制母兔同期发情具有重要作用。不同光周期时,光信号首先作用于哺乳动物视网膜上的光受体细胞,诱导黑视蛋白(melanopsin)及其编码基因Opn4表达量增加,导致视网膜节细胞(intrinsically photosensitive retinal ganglion cells,ipRGCs)兴奋,并由ipRGCs轴突构成的视网膜下丘脑束(retinohypothalamic tract,RHT)传导到哺乳动物视交叉上核(suprachiasmatic nucleus,SCN)[1-3],调节松果体褪黑激素(melatonin,MLT)分泌。光信号通过MLT转化为动物体内的化学信号,从而调控动物的多种生理功能[4]。MT主要通过结合其G蛋白偶联膜受体MLT1(Mel1a)、MLT2(Mel1b)实现其功能[5-6]。
已有研究发现,人类的情绪会受到光色的强烈影响[7-8]。Bourgin等[9]研究指出,光色对哺乳动物行为的影响重要且复杂。母兔周期化繁殖效率显著影响兔场经济效益。短日照季节母兔乏情现象明显,群体同期发情率变异大,目前只能依靠外源性激素实现群体同期发情。本研究通过不同LED光色对兔选择性停留行为及其体内激素变化角度开展研究,探讨光色调控母兔发情的分子机制,旨在为提高家畜繁殖生产中光需求与利用的针对性提供研究基础。
1 材料与方法
1.1 试验动物
选用江苏省农业科学院六合动物科学基地实验兔场提供的5月龄新西兰后备母兔,生产许可证号为SCXK(苏)2017-0008。
1.2 行为学观察试验
选用40只体质量相近的母兔进行行为学观察试验,试验在江苏省农业科学院六合动物科学基地人工光照控制室内进行。人工光照控制室内正方形漏缝木地板平台(6 m×6 m)上,四周用敞门兔笼合围,呈回字形,每边6个兔笼,笼内均匀放置常规乳头式饮水器及料槽,母兔可自由出入平台上任一兔笼。兔笼上方60 cm处安装LED灯带,向下投射红光(660 nm)、绿光(540 nm)、蓝光(480 nm)和白光(400~760 nm),采用动态光照控制系统(软著登字第1551136号),将不同颜色LED灯带的光照强度与光照时间均设置为100 lx,光照制度为12 D∶12 L。每周随机调整1次LED灯带位置,避免固定位置的光色可能成为条件反射的因素,并排除温度、声音、气味刺激等干扰。动态光照控制系统在光照试验开始后0.5 h及灯光关闭前0.5 h通过控制系统逐步调节光照强度,避免突然开关灯导致的应激反应。人工光照控制室内温度控制在24 ℃,并通过机械通风方式,保证空气质量。平台正上方选用夜视功能广角高清摄像头记录并统计母兔选择性停留行为。预试验期7 d,试验期100 d。
1.3 光色试验
1.3.1 试验分组
选择体质量接近的300只纯种新西兰后备母兔,随机分成5组(白炽灯对照组、白光组、红光组、绿光组、蓝光组),每组3个重复,每个重复20只。在秋季日照时长变短后开始试验,每周进行3次补光,观察不同光色对母兔同期发情的影响。预试验期7 d,试验期100 d。试验期间,室内温度控制在24 ℃,并通过机械通风方式,保证空气质量,排除温度、声音、气味刺激等干扰。
1.3.2 光照方案
每个处理组均采用双层遮阳窗帘进行分隔,单独隔间。母兔采取不锈钢单笼饲养,兔笼规格为60 cm×60 cm×60 cm。兔笼上方60 cm处分别安装LED灯带,向下投射红光(660 nm,R)、绿光(540 nm,G)、蓝光(480 nm,B)和白光(400~760 nm,W);对照组选用白炽灯(400~1 050 nm,C)照明,灯泡每隔2 m安装1个,安装高度为兔笼上方2.5 m处(通过testo-540照度仪监测,此高度下光照强度与LED灯带下的光照强度一致)。正式试验中,每周五至周日18:00至次日06:00,光照周期为12 D∶12 L,其他时间打开遮阳窗帘,不进行任何补光处理,每组内光线随自然环境改变同步变化并保持一致。LED灯带与白炽灯对照组采用动态光照控制系统(软著登字第1551136号)控制,光照强度为100 lx。光照试验开始时,每周通过照度仪监测光照强度,保证各组间光照强度一致。开始后0.5 h及灯光关闭前0.5 h,通过控制系统实现光强度0~100 lx和100~0 lx过渡期,以避免突然开关灯产生应激。
1.3.3 样本采集
正式试验期内,每个处理共分6次,每组分别选择5只发情母兔,于14、28、42、56、78、90 d的23:30至次日00:30进行耳缘静脉采血,进行MLT浓度及血清类激素测定。在LED光处理后的第91天早晨,在每组试验兔中选择体质量接近、处于同一生理阶段(处于发情中后期,外阴颜色为深红或紫红)母兔各5只进行屠宰。屠宰时每只母兔右侧卵巢取出,放入4%多聚甲醛磷酸缓冲液(0.1 mol·L-1,pH 7.4)中固定48 h后制成常规石蜡切片进行观察。同时迅速取下每组母兔视网膜、SCN、松果体、下丘脑、垂体、卵巢分别进行液氮保存,以待后期试验。试验兔屠宰操作在江苏省农业科学院动物福利和伦理委员会的规定下执行。
1.3.4 饲养管理与营养水平
试验期内母兔均为自由采食和饮水,其他饲养管理按常规进行。水、饲料、饲养人员、饲养管理各组均一致。按照NRC兔营养标准进行饲料配置并制成全价颗粒饲料,饲料配方见表1。
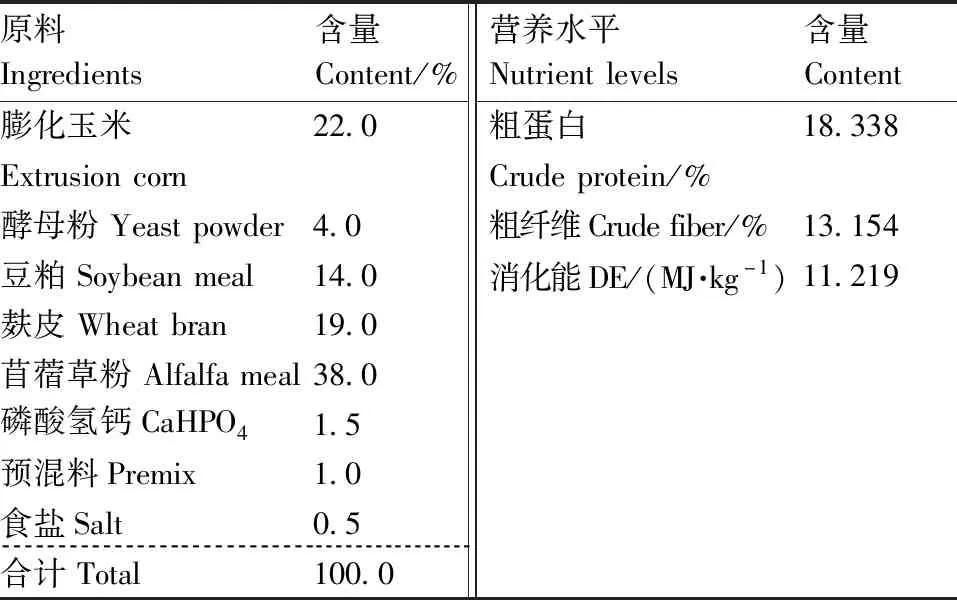
表1 基础饲粮组成及营养水平(风干基础)
预混料为每1 kg饲粮提供VA 300 000 IU, VD3200 000 IU, VE 4 000 mg, VK350 mg, VB1400 mg, VB2500 mg, VB6200 mg, VB124.2 mg,叶酸 50 mg,泛酸 2 200 mg,生物素 1 600 mg, 氯化胆碱 120 mg, VC 12 000 mg, Fe 4 000 mg, Zn 3 200 mg,Mn 1 600 mg, I 80 mg, Se 12 mg, Cu 500 mg。粗蛋白质、粗纤维、消化能均为测定值。
The premix provided the following component per kg of diets: VA 300 000 IU, VD3200 000 IU, VE 4 000 mg, VK350 mg, VB1400 mg, VB2500 mg, VB6200 mg, VB124.2 mg ,folic acid 50 mg, pantothenic acid 2 200 mg, biotin 1 600 mg, choline chloride 120 mg, VC 12 000 mg, Fe 4 000 mg, Zn 3 200 mg, Mn 1 600 mg, I 80 mg, Se 12 mg, Cu 500 mg. Crude protein, crude fiber and DE were measured values.
1.3.5 基因相对表达量测定
使用Trizol法提取样品组织总RNA,并用琼脂糖电泳检测。将提取的总RNA进行反转录。取0.2 mL PCR管,加入含2 μg RNA的溶液,加入1 μL oligo (dT) 18,用去离子水补足至12 μL,于PCR仪上70 ℃保温5 min后迅速置于冰上冷却。依次加入4 μL 5×buffer、2 μL 10 mmol·L-1dNTPs、1 μL RNA抑制剂和1 μL反转录酶,混匀后于PCR仪上42 ℃保温60 min,结束后80 ℃保温5 min灭活反转录酶。
用于基因表达检测的引物及其序列见表2。取0.2 mL PCR管,配制如下反应体系(每个反转录产物配制3管):12.5 μL 2×qPCR Mix、2 μL 7.5μmol·L-1基因引物、2.5 μL反转录产物、8.0 μL ddH2O。将cDNA原液稀释20倍,实时荧光定量PCR检测目的基因的mRNA表达。荧光定量PCR反应体系为:SYBR®Green Real-time PCR Master Mix 5 μL,上、下游引物各0.5 μL,cDNA模板2 μL,补足ddH2O至10 μL。反应条件为:95 ℃预变性3 min;95 ℃变性30 s,62 ℃退火22 s,72 ℃延伸20 s,共24个循环。每个样品重复1次,每板做1个用ddH2O和不加cDNA阴性模版的扩增,以检测水和试剂是否被污染。
1.4 血清类激素及MLT测定
血清类激素β-内啡肽(β-endorphin,β-EP)、5-羟色胺(5-hydroxytryptamine,5-HT)、乙酰胆碱(acetyl choline,ACH)、皮质醇(cortisol)及MLT浓度均按照试剂盒(南京建成)规定的方法进行检测。
1.5 统计分析
选择性停留率:统计母兔选择性停留在不同光色下的数量,统计时间段为光照周期的亮灯时间段,暗灯时段不参与统计。总共统计30次,每次统计时间为24 h,期间采用监控方式,每隔10 min观察记录1次母兔停留在不同光色下的数量。同期发情率统计:在LED光处理后的第14、28、42、56、78、90天上午,采用外阴检查法对母兔进行同期发情效果鉴定,当外阴出现大红或深红色则统计为发情,登记并统计发情母兔占该组试验兔的比率即为同期发情率。选择性停留率和同期发情率均采用SPSS 18.0软件One-Way ANOVA法进行方差分析,结果以“平均值±标准误”表示。
实时荧光定量数据的统计分析用2-ΔΔCt法进行,将目的基因的Ct值用内参基因mRNA的Ct值进行校对,计算目的基因的相对量。计算公式如下:

表2 基因表达检测用引物信息
ΔΔCt=(CtTarget-CtTBP)X-(CtTarget-CtTBP)control。
(1)
式(1)中:以对照组的目的基因CtTarge和内标基因CtTBP差值的平均值为参照进行计算;X表示任意1个样本。通过式(1)计算出每1个样本目标基因的表达,通过内标基因校正后相对于目的基因表达的倍数。
2 结果与分析
2.1 不同LED光色对母兔选择性停留行为的影响
通过连续摄像的方式,获得不同LED光色环境下母兔对活动空间选择性停留的观察数据。LED红光组、LED绿光组、LED蓝光组和LED白光组选择性停留率分别为(15.70±1.41)%、(2.60±1.34)%、(4.10±0.56)%、(8.90±1.19)%。不同LED光色能够影响母兔活动空间的选择,LED红光环境更易被母兔接受并停留,其停留频率分别比LED绿光、LED蓝光高13.1%和11.6%,差异显著(P<0.05);与LED白光组相比,LED红光环境中母兔的选择性停留率高6.8%,差异不显著(P>0.05)。
2.2 不同LED光色对后备母兔血清类激素含量的影响
如表3所示,LED红色光及白炽灯组后备母兔血清中的β-内啡肽、5-羟色胺浓度显著(P<0.05)高于LED绿光组和LED蓝光组,但与LED白光组差异不显著(P>0.05);血清乙酰胆碱在各处理间差异不显著;LED红光环境中皮质醇浓度显著(P<0.05)高于LED绿光组。

表3 不同LED光色环境下后备母兔血清类激素差异(n=30)
2.3 不同LED光色对母兔同期发情的影响
母兔同期发情率统计结果显示,LED红光组、LED绿光组、LED蓝光组、LED白光组和白炽灯对照组同期发情率分别为(85.64±6.59)%、(80.98±8.34)%、(78.31±7.23)%、(75.13±9.66)%、(64.75±5.33)%,LED红光组母兔同期发情率显著(P<0.05)高于白炽灯对照组和LED白光组,其他处理差异不显著。
2.4 不同LED光色对母兔视网膜黑视蛋白编码基因Opn4 mRNA表达的影响
如图1所示,母兔视网膜内Opn4基因的转录受到LED不同光色的影响,LED红光组母兔视网膜内Opn4基因表达量显著(P<0.05)高于白炽灯组和LED绿光组,但与LED白光组及蓝光组相比差异不显著。
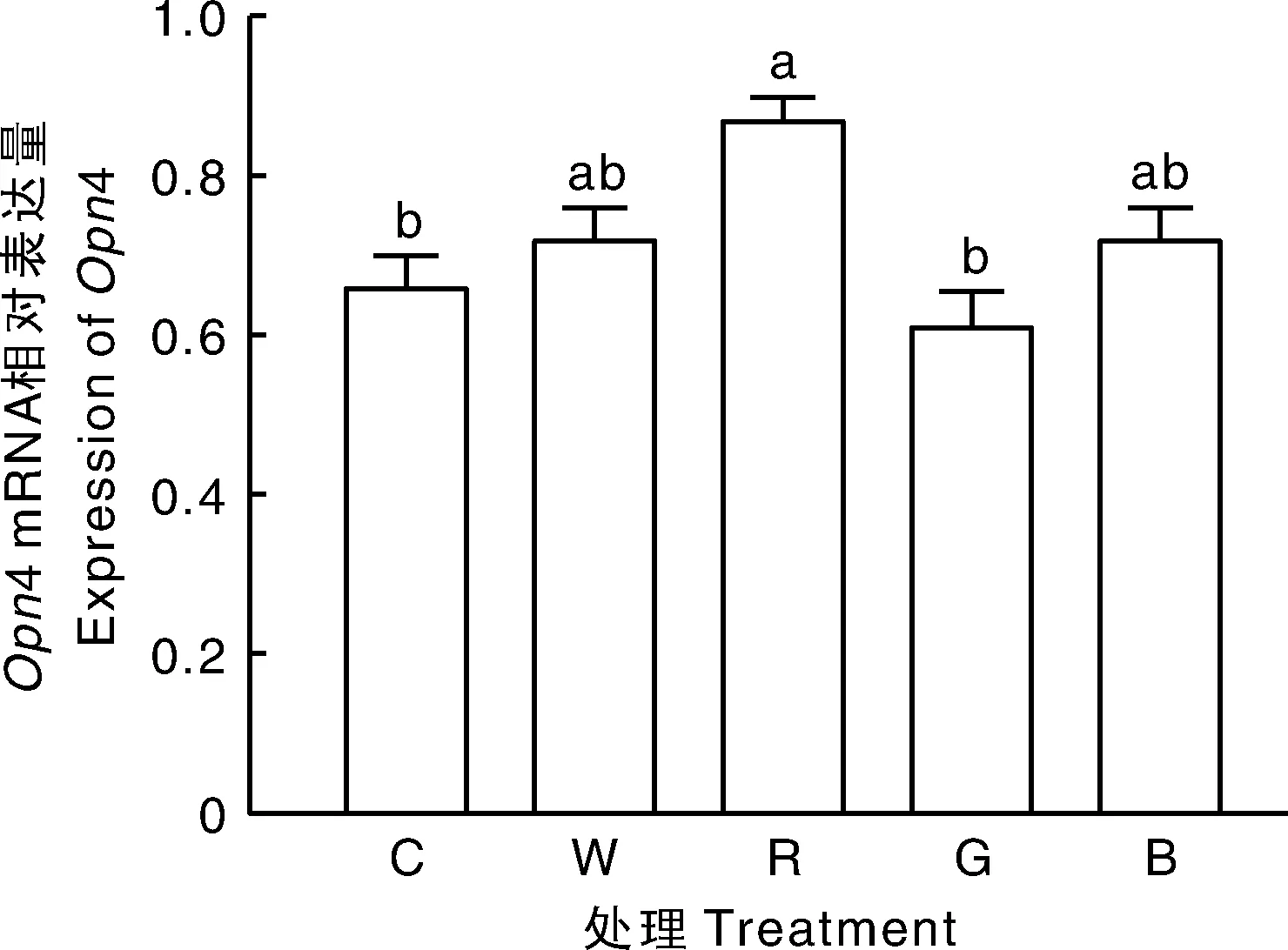
不同柱上无相同字母表示处理间差异显著(P<0.05)。下图同。Bars marked without the same letters indicated significant difference within treatment at P<0.05. The same as below.图1 不同LED光色对母兔视网膜黑视蛋白Opn4 mRNA表达的影响Fig.1 Effect of different light colors of LED on expression of Opn4 mRNA in retina of female rabbit
2.5 不同LED光色对母兔MLT分泌及其合成限速酶Aanat mRNA表达的影响
如图2所示,不同LED光色对特定时间段(23:30至00:30)母兔血清MLT含量有影响。与LED绿光组相比,LED红光组母兔血清MLT含量显著(P<0.05)降低,与其他处理差异不显著。如图3所示,松果体内MLT合成限速酶AanatmRNA的表达受到LED不同光色的影响。与LED白光相比,LED红光能显著(P<0.05)抑制松果体内AanatmRNA表达。
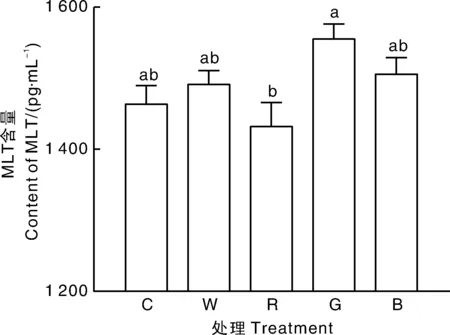
图2 不同LED光色对母兔血清MLT含量的影响Fig.2 Effect of different light colors of LED on serum MLT content of female rabbits
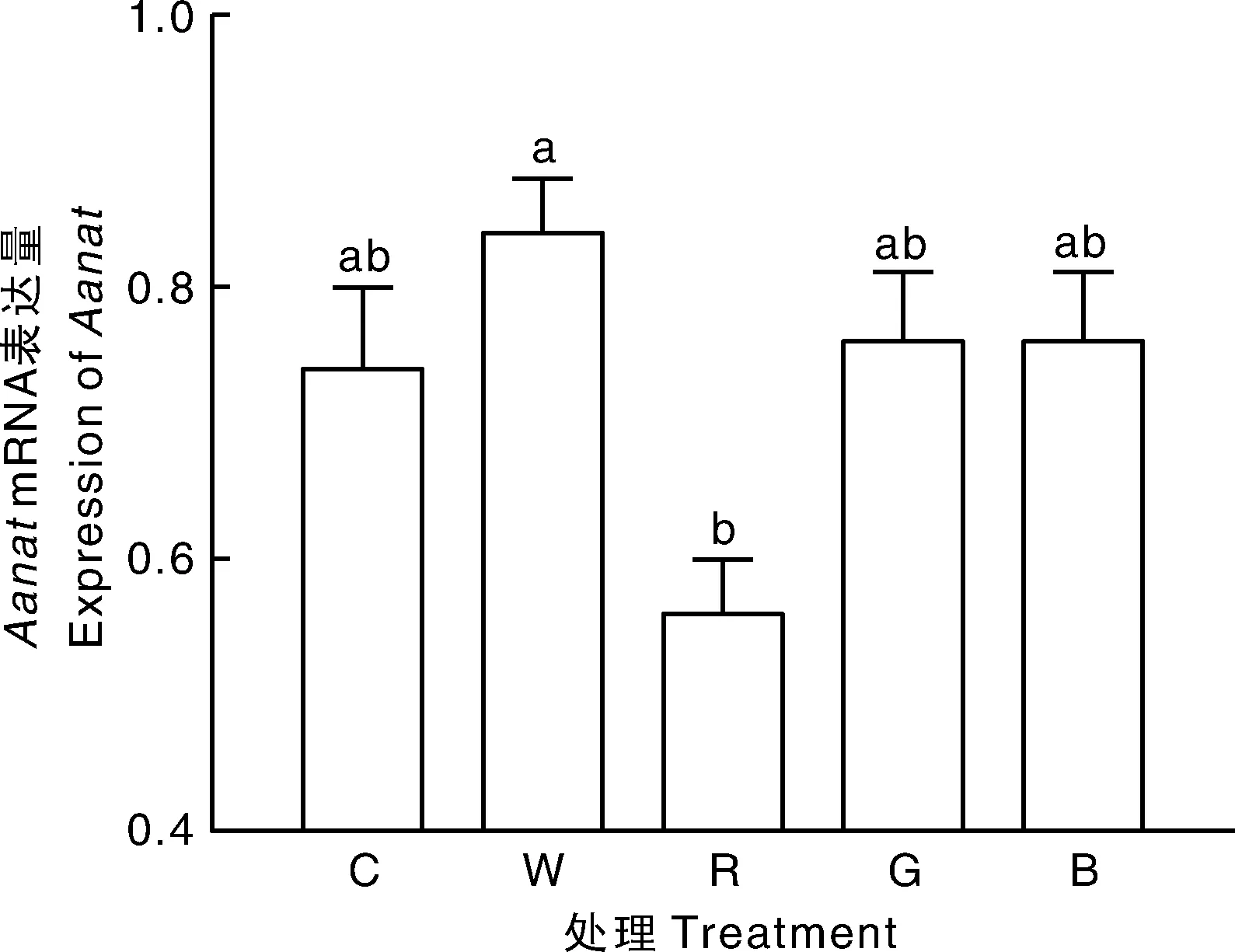
图3 不同LED光色对母兔松果体MLT合成限速酶Aanat mRNA表达影响Fig.3 Effect of different light colors of LED on expression of Aanat mRNA in female pineal gland
2.6 母兔MLT受体mRNA表达谱
如图4所示,母兔MLT受体Mel1amRNA在母兔视网膜、SCN、松果体、下丘脑、垂体、卵巢均有表达,并且与其他组织相比,在松果体中的表达量差异极显著(P<0.01);MLT受体Mel1bmRNA在母兔视网膜、SCN、松果体、下丘脑、垂体、卵巢表达。
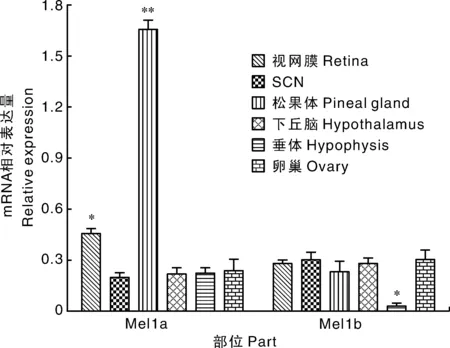
*和**分别表示显著性差异P<0.05和P<0.01。*, ** indicated significant differences at P<0.05 and P<0.01, respectively.图4 MLT受体mRNA表达谱分析Fig.4 Analysis of MLT recepto mRNA expression profile
2.7 不同LED光色对Mel1a/b mRNA表达的影响
检测MLT受体Mel1a/bmRNA在视网膜、SCN、松果体、HPG轴中的表达谱之后,选择母兔视网膜、SCN、松果体、下丘脑部位,观察不同LED光色对上述部位MLT受体Mel1a/bmRNA表达的影响。如图5所示,LED红光组母兔松果体中Mel1amRNA表达量显著(P<0.05)高于LED绿光组和蓝光组,但与白炽灯组和LED白光组相比差异不显著;不同光色对Mel1a基因在母兔视网膜、SCN、下丘脑部位的表达影响差异不显著;不同光色下Mel1b基因在母兔视网膜、SCN、松果体、下丘脑部位的表达差异不显著。
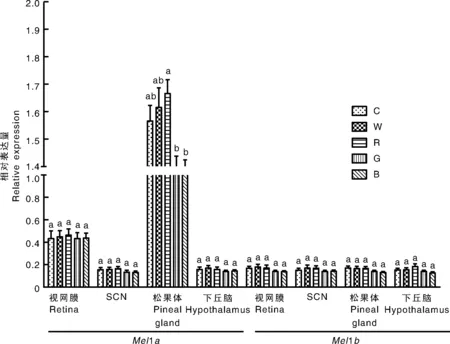
图5 LED不同光色对母兔视网膜、SCN、松果体、下丘脑部位MLT受体Mel1a/b mRNA表达影响Fig.5 Effect of different light colors on expression of MLT receptor Mel1a/b mRNA in retina, SCN, pineal gland and hypothalamus of female rabbits
3 讨论
3.1 不同LED光色对母兔行为的影响
不同光环境能引起哺乳动物昼夜节律、激素和行为的变化,如生物钟调整、抑制褪黑激素分泌、视网膜及瞳孔变化和生理参数的改变等[10]。这一系列光照的影响被称为非视觉生物效应,主要由被称为感光视网膜神经节细胞(ipRGCs)来调节。这类感光细胞不同于传统的视锥细胞和视杆细胞,在未发现视网膜神经节细胞(ipRGCs)之前,不同学者对于兔在不同颜色环境中的行为差异有不同的研究结论,研究观点普遍认为哺乳动物视网膜中的视锥细胞和视杆细胞数量差异是引起哺乳动物颜色视觉行为差异的主要因素。唐建荣等[11]对11只家兔进行色觉行为学实验研究,通过限制方位、光强等条件刺激因素,并排除声音、气味等干扰,结果显示,家兔具有红绿色觉分辨条件反射。与其他哺乳动物不同,兔具有夜行性行为特征,因此不能以其他哺乳动物的视觉特点对家兔进行颜色视觉的判定,而是需要根据对家兔行为特点进行研究。本研究结果表明,不同LED光色显著影响母兔活动空间的选择,LED红光环境更易被母兔接受并停留。此外,考虑到光照的非视觉生物效应取决于不止一种感光细胞,还取决于不同颜色LED光所携带的能量,光波长越短,光子能量越大。白炽灯波长为400~1 050 nm,LED红光波长为660 nm,推测对于兔这种夜行性敏感动物而言,与LED红光组相比,其他组较高能量的LED光可能会引起兔的应激反应,从而导致其选择性停留行为向光子能量较低的LED红光区域靠近,以缓解由于光色不同造成的警觉程度。
3.2 不同LED光色对母兔血清类激素的影响
Bourgin等[12]研究认为,光对哺乳动物生理和行为的影响取决于波长、辐照度、持续时间和光照时间。特定颜色的光环境对人类和其他动物的生理和新陈代谢具有影响。如在人体实验中,早上和晚上蓝光暴露均会严重改变健康成年人的葡萄糖代谢,而这些观察结果可能是由交感神经活性的增加和/或血浆褪黑激素的变化介导的[13]。此外,Opperhuizen等[14]在大鼠上研究表明,夜间急性暴露于绿光环境下能够诱发大鼠葡萄糖稳态下降,除了营养信号外,光的颜色在机体外周代谢的调节中起着重要作用。而关于不同LED光色对血清类激素分泌的影响主要围绕人的研究展开,Figueiro等[15]研究发现,生理系统对短波长光(蓝色)最为敏感;相比于含短波长光比较少的光源(黄白色光),在白天暴露于合适剂量的强短波长光下(青白色光)能更积极地、客观地影响阿尔茨海默病痴呆患者的睡眠和行为。此外,Yang等[16]研究发现,LED光能通过抑制人体褪黑素分泌刺激肾上腺分泌皮质醇进而改变生理节律,调节人体警觉度和生物钟。
本研究结果表明,不同LED光色环境对母兔血清类激素浓度影响差异显著,在LED红色光及白炽灯环境中,母兔血清中的β-EP、5-HT浓度显著高于LED绿光组及LED蓝光组。β-EP致欣快与满足感,可以调节焦虑情绪的发生。5-HT能改善睡眠,减少急躁情绪,增强免疫机能,红光环境减少了母兔的警觉程度,更易让母兔接受并停留。皮质醇在操纵情绪等方面具有重要作用,母兔在LED红色光环境中血清皮质醇浓度显著高于LED绿光组及LED蓝光组。红色环境氛围下,母兔表现出显著愉悦感,并且在处理一段时间后,由于发情促动,母兔体内皮质醇含量显著提高,表现出躁动不安的情况。
3.3 不同LED光色对母兔同期发情影响
已有关于光色对兔繁殖性能的影响的研究相对较少。宋亚鹏[17]研究发现,白色LED灯对提高母兔繁殖性能的效果要优于白炽灯,说明LED光源控制母兔同期发情技术可应用于商品肉兔繁殖生产中。李士栋[18]采用不同补光方式对哺乳期母兔的生产性能进行比较,发现在夏季使用LED灯后,母兔受胎率和产仔率比使用节能灯的母兔高5%。崔双保等[19]研究发现,LED光源显著提高了母兔同期发情率、受胎率、产仔数、活仔数、出生窝质量和断奶窝质量。Kalaba等[20]研究发现,红光可增加母兔繁殖性能及仔兔存活率。本试验中,LED红光组母兔同期发情率显著高于白炽灯组和LED白光组,说明LED光可用在规模兔场周期化繁殖生产模式中,并且能够提高母兔的繁殖性能。
3.4 不同LED光色对母兔视网膜黑视蛋白Opn4 mRNA表达的影响
在光周期影响哺乳动物繁殖行为的研究中,前期研究主要围绕视锥和视杆细胞2个方向,但是近年来对视锥和视杆细胞缺失的小鼠研究发现,还存在光信号的黑视蛋白调节途径。黑视蛋白是一种视网膜神经节细胞表达的感光视蛋白,其由Opn4基因编码,被认为是视锥和视杆细胞之后的哺乳动物第3种光感受性细胞[21]。环境中的不同光信息在视网膜经节细胞中黑视蛋白的作用下,通过视网膜-下丘脑束视神经传递到SCN,进而影响松果体调控褪黑激素的分泌,而褪黑激素再参与调节腺垂体促性腺激素的分泌最终发挥对动物繁殖行为的调节。本试验中,与白炽灯对照组和LED绿光组相比,LED红光显著增加兔视网膜内Opn4基因的表达,与靳二辉等[22]在鸡视网膜上Opn4-2 mRNA表达模式一致。
3.5 不同LED光色对MLT分泌及MLT合成限速酶Aanat mRNA表达的影响
松果体褪黑激素分泌属于昼夜波峰波谷连续分泌。孟繁臣[23]研究发现,不同光色对母兔在授精之前血清中褪黑激素水平有显著影响,红光和绿光能显著降低母兔血清褪黑激素水平。Kalaba等[24]研究发现,红色灯光能降低母兔血液中褪黑激素水平,提高FSH和雌二醇水平,且母兔的分娩率及产仔情况都有所提高。Jin等[25]研究了松果体和视网膜中不同单色光对鸡血浆褪黑激素水平和AanatmRNA表达的影响,其研究结果显示,与绿光和红光相比,白光和蓝光表现出对MLT分泌的微弱抑制,其抑制作用的大小为绿光<红光<白光<蓝光,绿光的松果体和视网膜中AanatmRNA的表达与血浆MLT水平的变化一致。本研究中,红光能够降低母兔血清褪黑激素水平,且AanatmRNA表达也受到LED不同光色的影响,LED红光能显著抑制松果体内AanatmRNA表达,这一结果与Jin等[25]在禽类上的研究结论不一致,分析这一差异的原因可能是哺乳动物只有眼球这1个光输入来源来调控从感知到神经内分泌再到相关基因表达的级联反应,而禽类拥有多个光信号输入路径,这种生理上的差异可能是导致机体感受不同光色后的下游基因响应差异不同的原因。
3.6 母兔MLT受体Mel1a/b表达谱及不同LED光色对Mel1a/b mRNA表达的影响
褪黑激素受体在很多动物中枢系统和外周组织中都有分布[26],表明褪黑激素受体对于机体生理功能的调节从神经内分泌系统到外周组织均起作用[27]。在人类上,褪黑激素受体在SCN、视上核、室旁核、海马、大、小脑皮质、神经胶质细胞及神经中枢以外的系统如心血管系统、消化系统、生殖系统中均有表达[28];在禽类中,褪黑激素更是直接调节禽类的神经内分泌功能,特别是在季节性繁殖的禽类,在不同靶细胞类型中,尤其是卵泡膜表面和颗粒细胞表层,褪黑激素通过激活受体信号通路参与禽类卵巢功能[29]。本研究发现,母兔褪黑激素受体Mel1a mRNA在母兔视网膜、SCN、松果体、下丘脑、垂体、卵巢均有表达,并且在松果体中表达量显著增加;MLT受体Mel1b mRNA在母兔视网膜、SCN、松果体、下丘脑、垂体、卵巢表达;LED红光组母兔松果体中Mel1a mRNA表达量显著高于LED绿光组和蓝光组,但与白炽灯组和LED白光组相比差异不显著。这一表达谱分析结果与上述褪黑激素受体在不同物种体内神经中枢及生殖系统上的表达模式基本一致,表明褪黑激素受体直接参与了母兔繁殖调控进程。
4 结论
规模兔场短日照季节周期化繁殖生产中,每周连续三次,每次晚间18:00至凌晨6:00之间,采用100 lx强度的LED红光进行补光,可提高母兔同期发情率;LED红光能够通过Opn4-MLT通路对兔行为及同期发情产生影响。
猜你喜欢
四川畜牧兽医(2022年10期)2022-10-28
孩子(2020年8期)2020-08-13
小天使·五年级语数英综合(2019年10期)2019-11-09
中国医学创新(2019年36期)2019-01-15
百科知识(2016年23期)2016-12-24
戏剧之家(2016年22期)2016-11-30
戏剧之家(2016年22期)2016-11-30
农家科技(2016年4期)2016-06-21
科技与创新(2015年3期)2015-03-31
农村农业农民·B版(2011年5期)2011-06-16
