
福建红壤侵蚀区马尾松林补植套种阔叶树对土壤氮矿化的影响
2020-12-22 11:41尤龙辉
防护林科技 2020年10期
尤龙辉
(1.福州市林业局自然保护地规划发展中心,福建 福州 350007;2.福建省林业科学研究院,福建 福州 350012)
土壤中氮素主要以有机氮形式存在,约占土壤氮库的92%~98%。自然界中绝大多数植被不能直接吸收有机氮,必须通过微生物矿化作用将有机氮转化成有效氮供植被吸收利用[1]。土壤中的有效氮主要是以铵态氮和硝态氮形式存在的矿质氮[2]。矿质氮的组分、数量及氮矿化速率反应土壤氮素积累和供应的有效性,是森林生态系统生产力及其生态功能重要的反馈指标[3,4],在同一研究区域,不同林分类型主要通过植被组成、凋落物及细根数量和质量、根系分泌物、土壤理化性质及土壤动物和微生物等影响有机氮矿化[4]。因此,深入了解森林土壤氮矿化过程,对揭示森林生态系统土壤氮循环和养分循环及评价森林生态系统的可持续经营提供理论依据。
马尾松(Pinusmassoniana)在我国南方亚热带地区广泛种植,是我国主要植树造林树种之一,长期以来产生了巨大的生态和经济效益,在我国林业生产建设中有着举足轻重的重要地位[5]。但是长时期采取单一针叶纯林的经营管理方式,特别是在立地较差或水土流失较严重的山区,森林病虫害、林地土壤肥力衰退、生产力下降及生态服务功能降低等问题逐渐凸显,目前已有近1/4的马尾松林严重退化,形成低质低效林[5,6]。因此如何有效改造低效马尾松林,以提升其林分生产力及生态服务功能一直倍受学界关注。对针叶林进行阔叶化改造,即通过抚育调整林分密度,进行林下补植套种乡土或适生阔叶树种,将林分逐步改造成复层、异龄、混交林,形成具有生物多样性、稳定性和多功能性的综合高效近自然森林生态系统的经营模式,该模式已在国内外积累大量实践经验:如Wang等[7]研究认为,红锥(Castanopsishystrix)与马尾松混交后,林地土壤碳贮量均高于红锥及马尾松纯林,Hart等[8]发现北美云杉(Piceasitchensis)和花旗松(Pseudotsugamenziesii)混交红枝桤木(Alnusrubra)能显著提高林地氮水平。王华等[9]人在中亚热带同会森林实验站研究表明固氮树种桤木(Alnuscremastogyne)与杉木(Cunninghamialanceolata)混交林的生态系统氮储量高于非固氮树种刺楸(Kalopanaxseptemlobus)与杉木混交林。徐海东等[10]研究杉木与蚊母(Distyliumracemosum)、紫楠(Phoebesheareri)、杨桐(Adinandramillettii)和红茴香(Illiciumhenryi)等阔叶树种混交后提出阔叶树种生物学特性是影响针阔复层林碳氮储量的重要影响因素。相关研究表明土壤有效氮库是限制林分生产力的主要因素[11],目前,有关补植套种阔叶树对马尾松林影响的相关研究主要集中在土壤理化性质[10,12]、林分生产力[13]及林下植被物种多样性[14]等方面,而探究影响其生态系统碳氮循环及林分生产力重要过程之一的土壤氮矿化作用的研究鲜见报道。
福建长汀是南方红壤重点水土流失区,早期推行种植的以马尾松纯林为主的大面积林分开始逐渐凸显各种问题,树种单一、林分结构简单、林下水土流失等现象使造林成效无法长期巩固,甚至出现生长一二十年但树体矮小的“小老头松”低质低效林。近年来,我国南方开展了大量关于红壤侵蚀区经济树种的种植试验[12-14],为红壤侵蚀区低效针叶林下套种经济树种,构建生态经济型针阔混交林提供了理论基础。有鉴于此,以长汀红壤侵蚀区低效马尾松林补植套种阔叶树种的改造试验林为研究对象,选取其中补植套种杨梅(Myricarubra)、无患子(Sapindusmukurossi)和黄栀子(Gardeniajasminoides)等3种典型改造林分,并以同地区未改造的低效马尾松纯林为对照,分析比较不同补植套种模式对林地土壤活性氮库及氮矿化速率的影响,为提升相似立地低效马尾松林分生产力及其改造树种选择和林分氮素管理提供参考。
1 材料与方法
1.1 研究区概况
研究区位于福建省龙岩市长汀县(25°33′—25°48′ N,116°18′—116°31′ E),气候类型为亚热带季风气候,年均气温17~18 ℃,年均降水量1 697 mm,年均日照时数1 900 h,无霜期平均270 d。该区为低山丘陵地貌,土壤为花岗岩发育的砖红壤。
供试林地位于长汀县河田镇(25°37′27″—25°37′59″ N,116°27′08″—116°28′06″ E),海拔309~412 m,坡度8°~ 32°。区域马尾松林为20世纪90年代末荒山造林,造林前期林地砾石含量较高,初植密度为2 m×2 m,采用裸根苗造林,该区域为典型的水土流失区,造林成活率和保存率不高,林龄为20~23 a,郁闭度0.5左右,后期纳入生态林管护,林下植被稀少,林木蓄积量明显低于同龄正常生长量,形成低效马尾松生态林。2014年通过套种不同阔叶树种对该地低效马尾松林进行改造。具体为:套种前期,对原有林分的枯病死木及被压木进行适当的卫生伐,保留林分郁闭度为0.5左右,然后在林隙挖规格为50 cm×50 cm×40 cm种植穴,穴底施腐熟有机肥1.5 kg,套种密度为2 300~2 500株hm-2,分别于定植后第2年与第3年6月,结合抚育扩穴除草进行追肥,在定植穴周边开小沟环施氮磷钾复合肥100 g。供试林地基本情况见表1,表中立地类型分类依据:坡位分为山脊、坡面、沟底;坡度等级分为平缓坡(0°~ 15°)、斜坡(16°~ 25°)、陡坡(26°~ 35°)和急坡(>35°);坡向中南、东南、西南和西坡归为阳坡,北、西北、东北和东坡归为阴坡。供试样地土壤理化性质、细根和凋落物质量见表2,试验期间样地气温及降雨量的季节变化见图1。
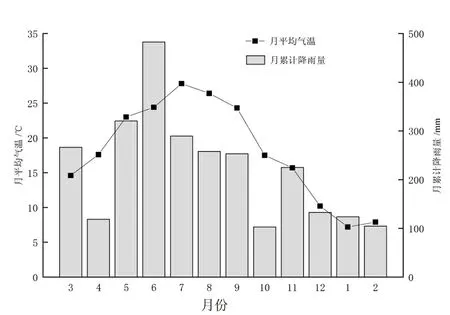
图1 2018~2019年研究区域月平均气温及月累计降雨量
1.2 样地设置与研究方法
3种模式林分及对照组均随机设置4个垂直投影为25 m×25 m的标准样地,样地间隔50 m以上,共计16个样地,总面积10 000 m2。每个样地随机选取一条对角线,等距布设10个点,采用PVC管原位培养法进行氮矿化试验,方法参考文献[15]:每个培养点分别打入内径5 cm,高20 cm的PVC管,随后在紧邻培养点处,再选取1个土壤采集点,打入尺寸规格相同的PVC管。培养点PVC管土壤原位培养30 d后,用PVC管顶盖盖住两头带回实验室,用手锯在管中部10 cm位置将管一分为二,并做好标记,将同一样地同一土层的PVC管内土壤混匀,拣去石砾和细根,四分法取部分土样过2 mm筛后,用以测定土壤铵态氮和硝态氮含量。与培养点紧邻的另外一根PVC管则在布管完成后当天带回实验室,也按照培养点PVC管的方法分成0~10 cm和10~20 cm两部分,将同一样地同一土层的PVC管内土壤混匀,然后拣去石砾,拣出细根(<2 mm),根据外形、颜色、弹性化区分死根和活根,并估算生物量[16],烘干粉碎过0.147 mm筛后备用,再四分法取部分土样过2 mm筛后用以测定土壤铵态氮、硝态氮、微生物生物量碳和微生物生物量氮含量,取部分土样研磨过0.147 mm筛后用以测定土壤全碳、全氮含量。每次收集PVC管后,在留下的管孔处采用浙江托普仪器有限公司生产的TZS-2X便携式数字瞬时温湿度计测量并记录取不同土层的土壤温湿度。PVC管的埋设时间分别在2018年4月(春季)、2018年7月(夏季)、2018年10月(秋季)和2019年1月(冬季)。
2018年4月,在每个样地随机均匀布设5个离地0.5 m,长×宽×高为1 m×1 m×0.25 m的凋落框,每个月收集一次框内凋落物,截至2019年3月。凋落物带回实验室,称鲜质量后,置于鼓风干燥箱内50 ℃烘干至恒质量,测含水率换算生物量。2018年5月,在每个样地的东南角,挖60 cm深的土壤剖面,分0~10、>10~20、>20~30、>30~40、>40~50、>50~60 cm 6个土层,用环刀取样,每层3个重复,测土壤容重和孔隙度。
1.3 测定项目与方法
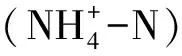
1.4 数据处理
利用Microsoft Excel 2016整理数据,SPSS 25.0统计分析,Origin 9.5绘制图表。采用三因素重复测定方差分析(Three-way Repeated Measures ANOVA)检验季节、模式和土层及其交互作用对土壤矿质氮和净氮矿化速率的影响;采用Pearson相关分析检验土壤理化性质、细根和凋落物质量与土壤净氮矿化速率的相关关系。
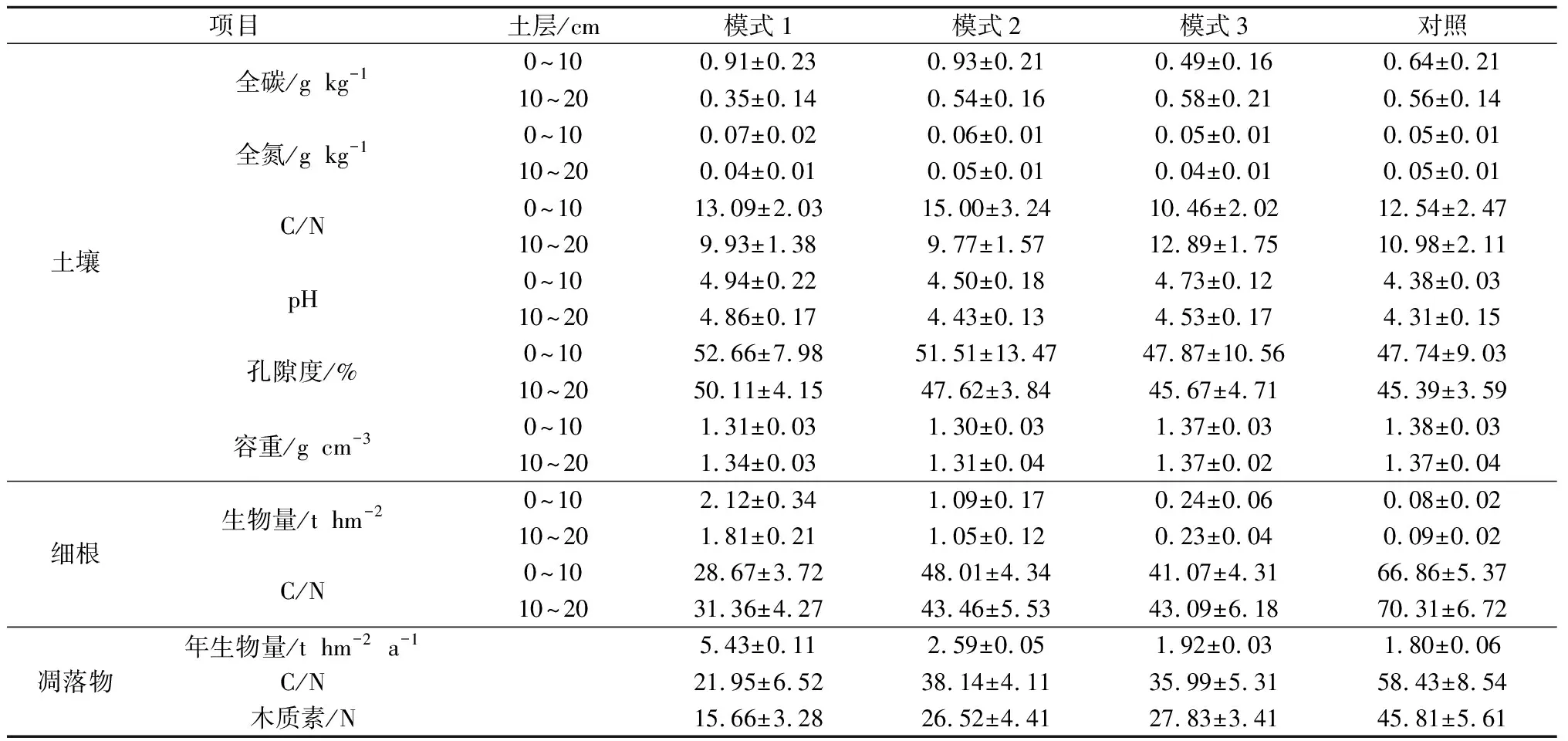
表2 供试样地土壤理化性质、细根和凋落物质量
2 结果与分析
2.1 3种模式林分土壤温度和湿度的季节变化
由图2可知,3种模式林分及对照组土壤温度均表现为夏季最高,春季和秋季次之,冬季最低,且随土层加深逐渐下降;土壤年均温度总体表现为对照组>模式3>模式2>模式1。土壤湿度总体表现为春季最高,夏季和冬季次之,秋季最低,0~10 cm和10~20 m土层无显著差异(P<0.05,下同)。
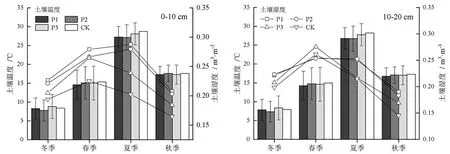
图2 3种模式林分土壤温度和湿度的季节变化
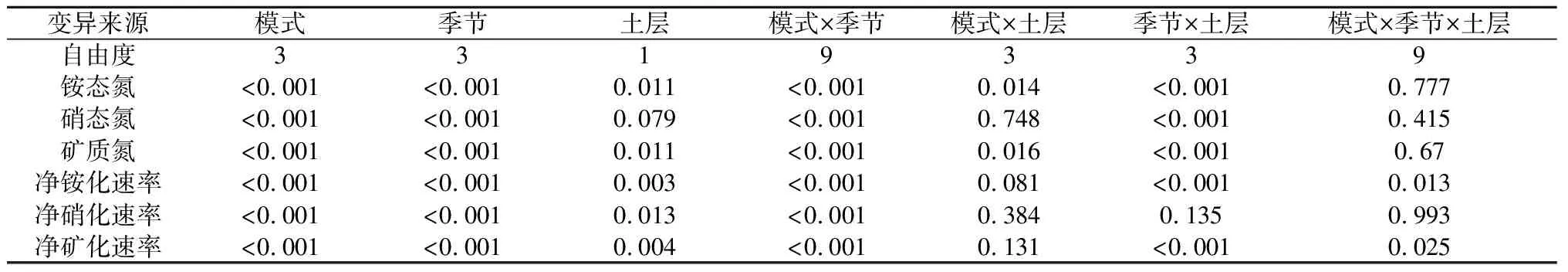
表3 模式、季节和土层对土壤氮矿化的重复测定方差分析
2.2 3种模式林分土壤铵态氮、硝态氮和矿质氮含量的季节变化
由图3可知,各模式林分及对照组土壤铵态氮、硝态氮及总矿质氮含量存在明显季节差异,均表现为夏季和秋季显著高于春季和冬季。0~20 cm土层平均土壤铵态氮和硝态氮含量总体上从大到小依次为模式1>模式2>模式3>对照组。其中模式1土壤铵态氮和硝态氮含量相比于对照组分别提升64.57%~125.03%和72.28%~166.52%,模式2分别提升55.36%~105.65%和77.87%~135.27%,模式3分别提升31.39%~58.62%和34.93%~69.33%。0~10 cm土层各季节土壤铵态氮含量显著高于10~20 cm土层,而硝态氮含量0~10 cm土层夏季和秋季略高于10~20 cm土层,冬季和春季则相反,各模式林分及对照组土壤年均铵态氮含量占年均矿质氮含量的89.35%~90.82%。重复测定方差分析显示(见表3,下同),模式×季节与季节×土层对土壤铵态氮含量、硝态氮含量和矿质氮含量均有极显著的交互影响(P<0.01,下同)。
不同大写字母代表不同模式间显著差异,P<0.05;不同小写字母代表不同季节间极显著差异,P<0.01;下同
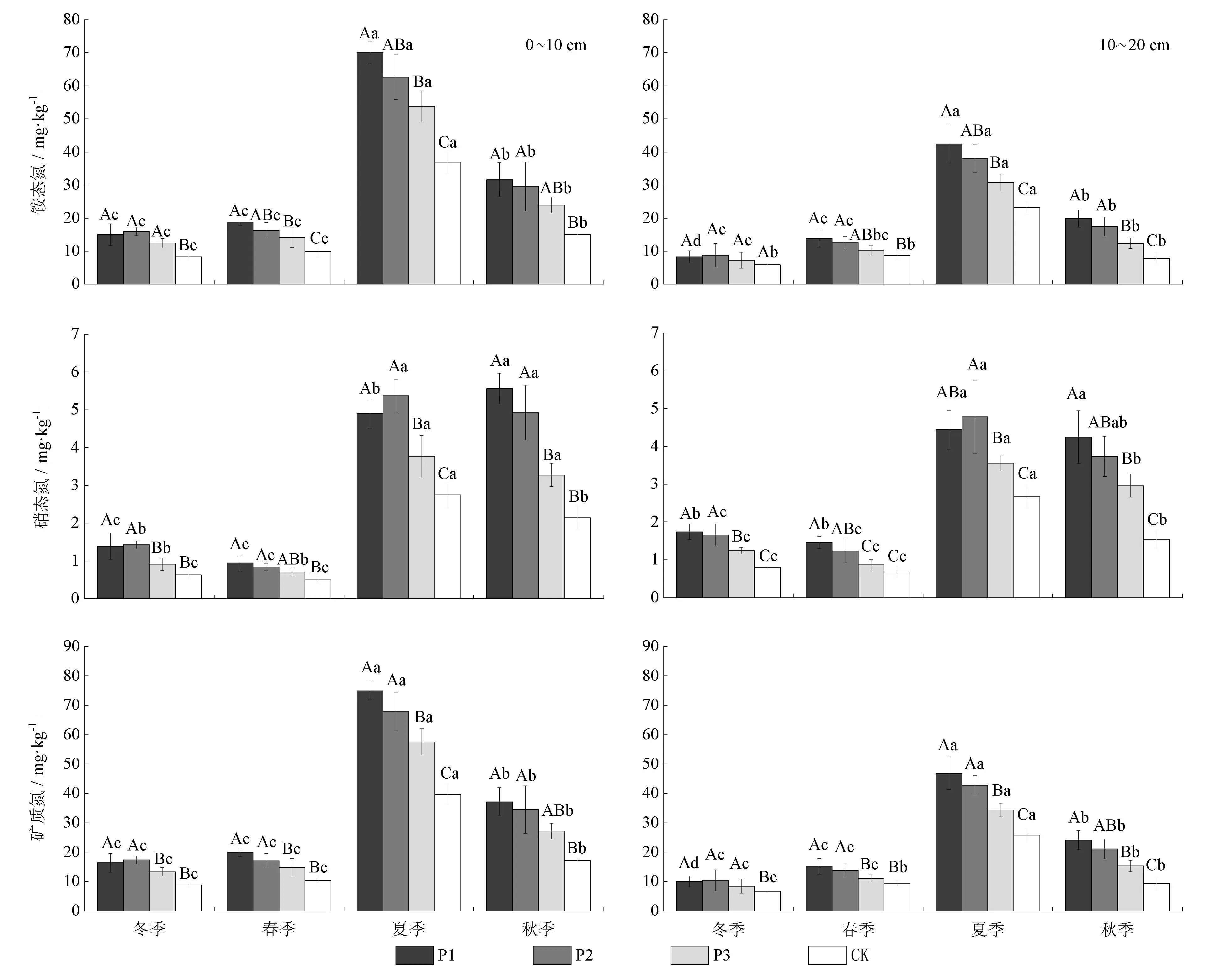
2.3 3种模式林分土壤净铵化速率、净硝化速率和净氮矿化速率的季节变化
由图4可知,3种模式林分及对照组净铵化速率表现为夏季最高,春季次之,秋季和冬季较小;净硝化速率则表现为夏季和秋季较高,春季次之,冬季最小为负值;净氮矿化速率表现为夏季最高,春季和秋季次之,冬季最小。在0~20 cm土层,土壤年均净铵化速率、年均净硝化速率和年均净氮矿化速率均表现为模式1>模式2>模式3>对照组。其中,模式1土壤净铵化速率和净硝化速率相比于对照组分别提升4.28%~168.09%和27.34%~158.11%;模式2分别提升12.0%~157.29%和48.79%~140.51%,模式3分别提升2.12%~64.31%和27.75%~55.29%。模式1~3及对照组土壤净铵化速率均随土层深度的增加而减小,0~10 cm土层相对于10~20 cm土层分别提升33.40%~213.33%、25.26%~163.63%、31.74%~116.66%和35.80%~104.00%;模式1~3及对照组土壤净硝化速率则相反,10~20 cm土层相对于0~10 cm土层分别提升11.38%~5.72%、8.87%~58.06%、7.04%~48.12%和14.92%~56.67%。各模式林分及对照组年均净铵化速率是年均净硝化速率的3.9~4.7倍,表明研究区试验林土壤氮矿化以铵化作用为主导。重复测定方差分析显示,模式×季节、季节×土层和模式×季节×土层均对土壤净氨化速率和净氮矿化速率有极显著交互影响。
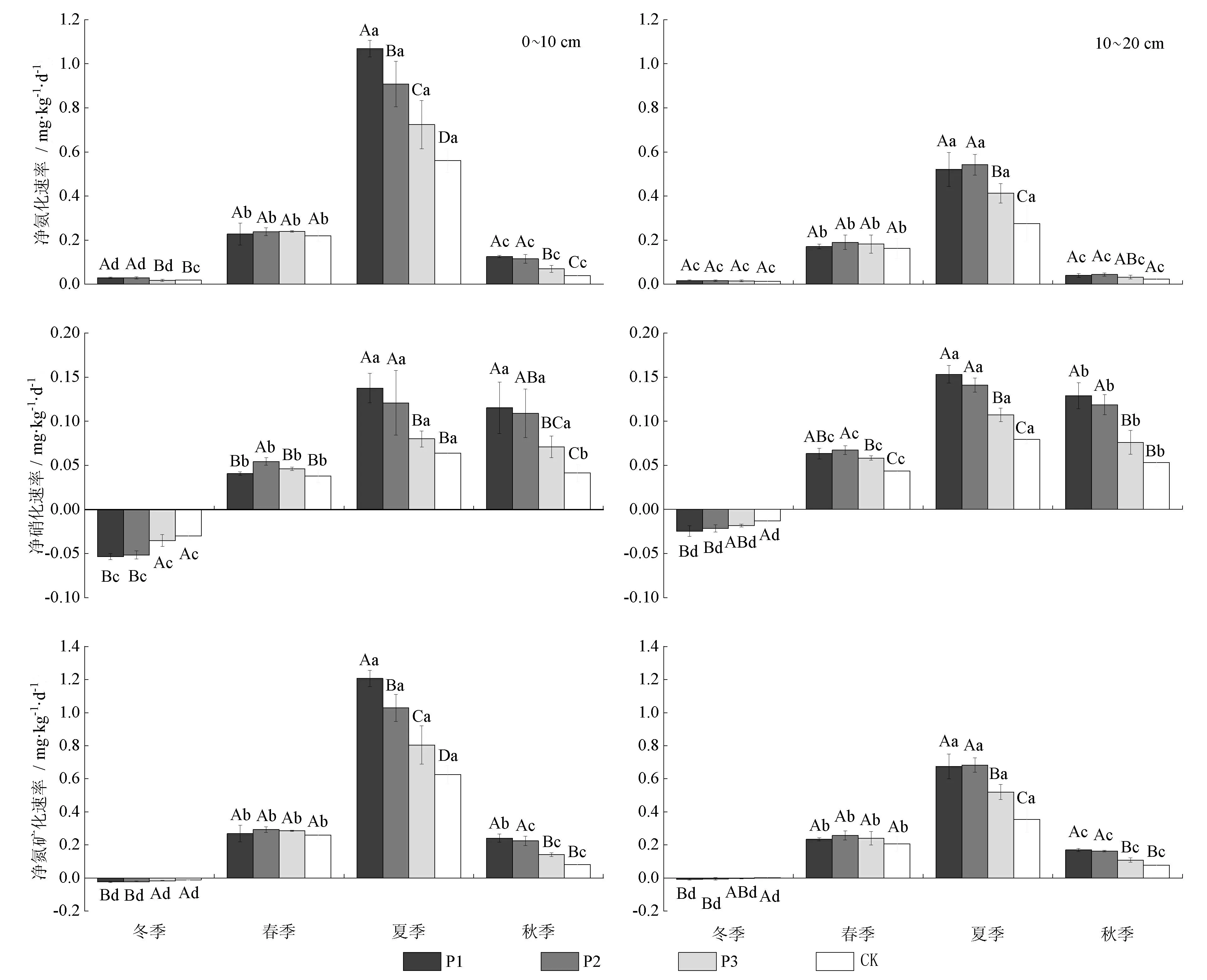
图4 3种模式林分土壤铵化、硝化和净氮矿化速率的季节变化
2.4 3种模式林分微生物生物量碳氮含量的季节变化
由图5可以看出,各模式林分及对照组土壤微生物生物量碳、氮总体表现出春季和秋季较高,夏季次之,冬季较低的季节动态。0~20 cm土层,土壤微生物生物量碳、氮含量大小排序均表现为模式1>模式2>模式3>对照组。模式1土壤微生物生物量碳、氮含量分别比对照组提升97.76%~306.66%和153.92%~548.72%;模式2分别提升61.63%~265.31%和130.13%~402.20%;模式3分别提升35.42%~148.13%和26.00%~403.66%。土壤微生物生物量碳含量和氮含量均表现为0~10 cm土层(77.05~475.6 mgkg-1和2.71~32.76 mgkg-1)高于10~20 cm土层(47.06~298.2 mgkg-1和1.77~18.27 mgkg-1)。
图5 3种模式林分土壤微生物生物量碳氮含量的季节变化
2.5 土壤理化性质、凋落物和细根质量与土壤净氮矿化速率的相关分析
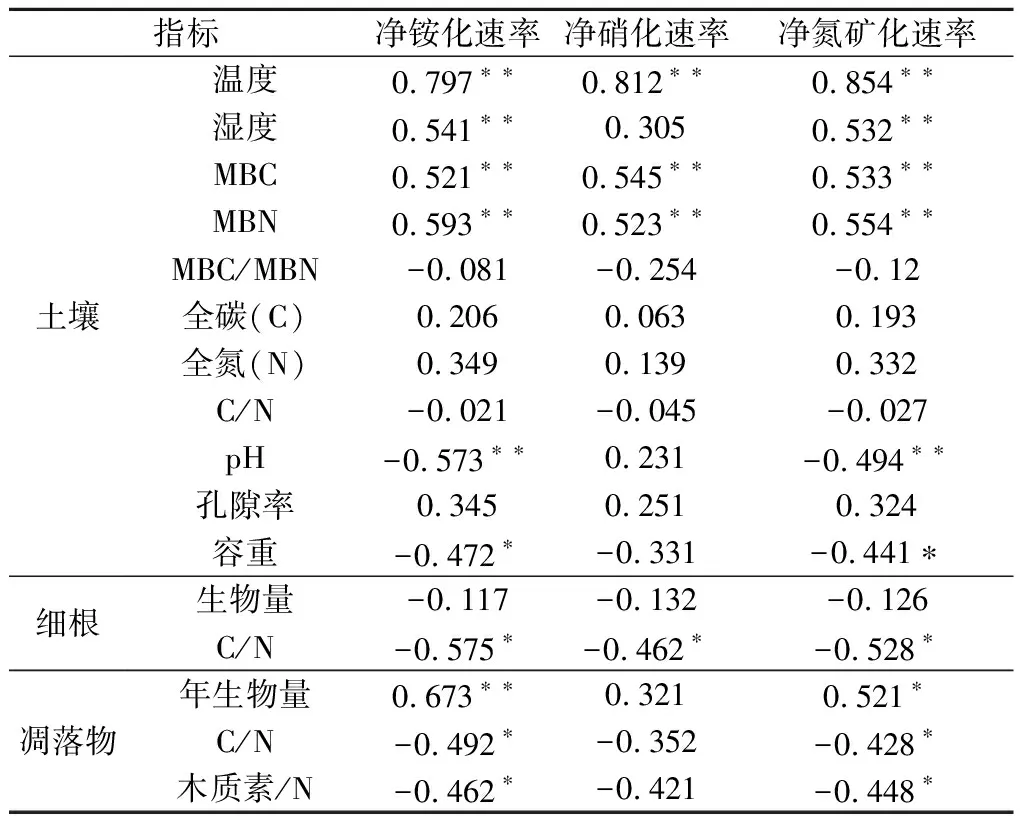
表4 土壤理化性质、细根和凋落物质量与土壤氮矿化速率的相关分析
Pearson相关分析表明(见表4),土壤净铵化速率、净氮矿化速率分别和土壤温湿度、土壤微生物生物量碳氮含量和年凋落物生物量呈极显著相关(P<0.01),与土壤容重、细根C/N、凋落物C/N、凋落物木质素/N呈显著负相关(P<0.05),而与土壤pH呈极显著负相关。土壤净硝化速率分别和土壤温度、土壤微生物生物量碳氮呈极显著正相关,与细根C/N呈显著负相关。
3 讨论
3.1 3种补植套种模式林分土壤氮矿化的时间变异性
重复测定方差分析表明,季节的主效应及模式和季节的交互作用显著影响试验林的土壤氮矿化速率,一方面与氮矿化底物有关,不同的补植套种树种物候各不相同,使不同模式林地凋落物量和根系周转的季节动态表现不同,而凋落物及死根作为土壤氮矿化作用的主要底物来源,其数量和质量直接影响微生物群落和数量,进而影响土壤氮矿化速率[15,17];另一方面则与土壤温湿度有关,土壤的氮矿化过程本质是微生物的生理代谢过程[17],而土壤温湿度对微生物的活性有重要影响。Cookson[18]等和Guntias[19]等研究表明,在一定土壤温度范围内,土壤氮矿化速率随土壤温度的增加呈指数递增。土壤水分则通过参与微生物生理代谢过程及改变土壤透气性[20],影响土壤氮矿化速率,在一定土壤湿度范围内,土壤微生物活性也随土壤湿度的增加呈现递增的趋势[20,21]。本研究相关分析也发现土壤微生物量碳氮含量、土壤温湿度与土壤氮矿化速率呈极显著正相关关系。试验地雨热同期,各模式林分的矿质氮含量和氮矿化速率总体上表现为夏季和秋季显著高于冬季和春季。本研究显示,3种模式林分及对照组不同季节土壤铵态氮含量及净铵化速率均显著高于硝态氮含量及净硝化速率。这与段伟[17]等和莫江明[22]等研究的亚热带和南亚热带马尾松林下土壤氮矿化规律一致,而与肖好燕[15]等对亚热带天然林、杉木人工林、格木人工林的土壤氮矿化规律相悖。这可能一方面与研究区酸性红壤会抑制氨氧化菌生长,以减少亚热带红壤侵蚀区水热丰沛的气候条件下土壤氮素通过硝态氮淋失和反硝化作用逸失的森林保氮机制有关[23],另一方面与马尾松偏好于通过较低能耗吸收利用铵态氮以适应水土流失区瘠薄立地[22,24],使铵态氮成为土壤矿质态氮的主要形式有关。3种模式林分及对照组土壤净硝化速率在冬季表现为负值,可能与冬季温湿度较低,试验林土壤铵态氮含量和氨氧化菌活性均下降,而植物根系吸收、微生物固持及降雨淋溶流失逐渐消耗土壤硝态氮库,使净硝化速率呈现负值有关。这与Yan[25]等对我国东部常绿阔叶林土壤净氮矿化速率最低值出现在冬季,且为负值的研究结果相一致,而与殷睿[26]等人研究得出的川西亚高山森林非生长季土壤净氮矿化速率高于生长季的结论不同,这可能是由于不同研究区域气候、植被-土壤系统不同,其氮矿化作用机制不同所致。
3.2 3种补植套种模式林分土壤氮矿化的空间变异性
3种补植套种模式及对照组土壤矿质氮含量均随土层深度的增加而减少,这与刘丹[27]等研究赣南红壤侵蚀区湿地松(Pinuselliottii)0~30 cm土层土壤有效氮库变异规律一致,主要是由于红壤侵蚀区土壤相对贫瘠,表层土壤由于丰富的凋落物输入及水热条件,有利于土壤微生物的生长、繁殖,进而促进土壤团聚体的形成和矿质氮库的累积,随着土层的加深,可矿化的有机底物减少,土壤孔隙度下降,透气性降低,水、肥、气、热等生态因子逐渐限制了土壤微生物的生长与繁殖,其种类和数量亦随土层加深而逐渐降低,土壤氮矿化速率下降,矿质氮库累积逐渐减少[27,28]。此外,本研究发现马尾松细根(<2 mm)生物量随土层深度呈倒金字塔分布,其中0~10 cm土层的细根生物量占比达到42.6%,这与程瑞梅[29]等的研究结果相似。细根的垂直分布结构,也是形成土壤氮矿化速率和矿质氮含量空间垂直分布差异的重要因素,因为细根分泌物中的有机物质、胞外酶及周转过程产生的死根对氮矿化过程也有一定的积极影响[30,31]。本研究中,各模式林分0~10 cm土层土壤铵态氮含量均显著高于10~20 cm土层,呈表聚性;0~10 cm土层硝态氮含量夏季和秋季略高于10~20 cm土层,冬季和春季则相反。这是由于红壤中的胶体粒子主要为高岭石和铁铝氧化物,易吸附固持带正电荷的铵态氮,而硝态氮带负电荷不易被红壤胶体吸附,在重力作用下,容易随土壤水向下淋溶[32],夏季和秋季由于干湿交替频繁,氨氧化菌较活跃,表层土壤净硝化速率较高,硝态氮的累积速率高于淋失速率及植被根系吸收和土壤微生物固持消耗的速率,使表层土壤硝态氮库高于下层土壤,冬季和春季氨氧化菌活性降低,上层硝态氮累积速率降低,垂直淋溶使下层土壤硝态氮库高于上层土壤。重复测定方差分析中也显示,土层的主效应和土层与模式的交互作用对铵态氮、净铵化速率影响显著,而硝态氮、净硝化速率的土层主效应不显著。
3.3 3种补植套种阔叶树种对马尾松林分土壤氮矿化的影响
3种补植套种模式林分土壤矿质氮含量和氮矿化速率相对于对照组均有显著提高,其中补植套种杨梅模式提升幅度最高,其次为补植套种无患子模式,而补植套种黄栀子模式提升幅度最小,重复测定方差分析也表明模式的主效应影响土壤矿质氮含量和净氮矿化速率。这可能与各套种树种的生物学特性有关:须根系树种杨梅虽根系分布较浅,但须根发达,且易与土壤中的Frankia放线菌共生形成固氮根瘤菌[33],吸收空气中的氮气并转化铵态氮,供杨梅吸收利用,大大减少与上层木马尾松竞争氮肥的矛盾,而增加的根系及其分泌物中的酶也能促进有机质分解,大大提升养分归还效率[34];无患子幼树时期较耐荫,细根分布在较深的土层,与细根集中于土壤表层的上层木马尾松形成良好的根系垂直分布结构,种间竞争矛盾较小,且无患子是落叶阔叶树种,凋落物凋落量大,一方面能为土壤微生物增加代谢底物,另一方面能有效拦截降雨径流,减少土壤有效氮素的流失;黄栀子须根也较发达,且细根周转速度较快[35],也能为土壤微生物提供丰富的代谢底物,但是黄栀子属矮灌树种,根系生物量与前两者相比较小,且易与上层木马尾松争夺氮素养分。此外,补植套种树种凋落物和细根质量不同也会影响林分的土壤氮矿化过程。覃宇等[36]研究发现,马尾松凋落叶分别与香樟(Cinnamomumcamphora)、香椿(Toonasinensis)、檫木(Sassafrastzumu)等阔叶树种的凋落叶混合后,均不同程度地促进了马尾松凋落叶中缩合单宁和总酚等难分解物质的分解。章晴等[30]表明固氮树种桤木与杨树混交通过改善细根养分活化能力和增加根际分泌物种类形成特有的混合根系根际环境提高杨树纯林土壤氮素供应能力。本研究中,净氮矿化速率与凋落物C/N、木质素/N和细根C/N呈显著负相关,凋落叶和细根C/N表现为对照组>补植套种无患子模式>补植套种黄栀子模式>补植套种杨梅模式,凋落叶木质素/N则表现为对照组>补植套种黄栀子模式>补植套种无患子模式>补植套种杨梅模式,说明补植套种阔叶树后均不同程度改变了原有马尾松林凋落物和细根的质量,其中马尾松林补植套种杨梅后,增加凋落叶和细根中氮素比例的幅度最大,能够有效激发土壤中氮源性微生物的生长繁殖,这与Santos等[29]和李贵才等[37]研究指出降低凋落物、细根C/N和木质素/N能够显著提高土壤的氮矿化速率的结论一致。再者,王传华[38]等和Ohashi[39]等指出,在中低纬度地带,森林土壤微生物活性受土壤湿度的影响较大。马尾松补植套种阔叶树后,改变了原有的冠层结构及林下光照微环境,减少了林地蒸发散,提高了林地土壤湿度,对土壤微生物活性也存在一定程度的影响,其中杨梅或无患子的树冠伸展及林下凋落物量显著高于黄栀子,能够形成较好的“保湿层”,对土壤微生物活性的影响也较大。
3.4 低效马尾松林补植套种阔叶树种的氮肥管理策略
马尾松补植套种阔叶树种后,表层土壤氮矿化速率、矿质氮库及微生物生物量均有所提升,这对于改善低质低效马尾松林分生产力和生态功能,均有重要作用:氮矿化速率加快,可以缩短马尾松林养分循环的周转时间;矿质氮库容增加,可为林分提供更多的活性氮养分,一定程度缓解低效林缺氮的困境;微生物生物量提高,虽然与植物根系存在一定的养分竞争的矛盾,但是微生物通过固持氮素养分,会减少氮矿化富氮时期的淋溶流失,而在缺氮时期通过周转重新归还于林分,提高氮素利用效率,培肥土壤。本研究表明,短期内通过补植套种杨梅或无患子对低效马尾松林分活性氮库的积累效应较明显,因此在抚育追肥中,可适当减少外源性氮肥的施入,避免氮素流失污染环境,另外过量的无机氮肥的施入会对土壤微生物产生毒害作用[24,40],反而抑制土壤的氮矿化作用,减少活性氮含量。
4 结论
红壤侵蚀区补植套种杨梅、无患子和黄栀子等3种阔叶树种对低效马尾松林不同季节或不同土层深度的土壤氮矿化过程均有显著影响。3种阔叶树种主要通过影响低效马尾松林下土壤温湿度、土壤微生物、土壤容重、土壤pH、细根和凋落物的数量和性质提高土壤矿质氮的转化。马尾松林下补植套种杨梅模式土壤净铵化速率和净硝化速率相比于对照组分别提升4.28%~168.09%和27.34%~158.11%,马尾松补植套种无患子模式分别提升55.36%~105.65%和77.87%~135.27%,马尾松补植套种黄栀子模式分别提升31.39%~58.62%和34.93%~69.33%。因此,短期内补植套种阔叶树有利于改善低效马尾松林土壤氮素循环,增加活性氮库的累积和供应,提升低效马尾松林分生产力和生态服务功能,进而保障马尾松林的可持续经营发展。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
今日农业(2022年16期)2022-11-09
今日农业(2022年14期)2022-09-15
建材发展导向(2022年10期)2022-07-28
新农业(2021年12期)2021-11-29
南方农业·中旬(2021年5期)2021-10-25
电脑迷(2020年5期)2020-12-18
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
