
钙离子对海假交替单胞菌生物被膜形成及厚壳贻贝附着的影响
2020-12-17 00:59常睿珩许康豪蔡雨珊杨金龙梁箫
大连海洋大学学报 2020年6期
常睿珩,许康豪,蔡雨珊,杨金龙、2,梁箫、2*
(1.上海海洋大学 国家海洋生物科学国际联合研究中心,水产种质资源发掘与利用教育部重点实验室,上海 201306; 2.南方海洋科学与工程广东省实验室(广州),广东 广州 511458)
生物被膜是附着在物体外侧或相互接触面的具备结构的微生物群体[1],由微生物及其分泌的胞外基质组成,为顺应生活环境而形成在活性或惰性材质外层[2]。在水体环境中,绝大部分固体附着基质的表层都存在着生物被膜。细菌附着到基质表层并产生胞外产物形成了生物被膜,这是生物被膜形成的重要部分[3]。影响生物被膜形成的因素有很多,如温度和盐度[4]、培养基中的营养盐[5]、不同种细菌[6]等。生物被膜能影响贻贝、藤壶等幼虫的附着变态[7-9],该现象是海洋经济贝类养殖和海洋防污损的研究热点。
钙离子是高度通用的细胞内信号,可以调节多种不同细胞的功能[10]。在海洋环境中,钙是一种经常在物体表面富集的元素,既可以是沉积的钙沉积物,也可以与其他海洋生物结合在一起[11]。钙能影响生物被膜的形成,特别是钙参与了细胞与基质之间的特异性和非特异性相互作用[12-13]。已有试验证实,钙离子能够促进大肠杆菌生物被膜和铜绿假单胞菌生物被膜形成[14-15],抑制霍乱弧菌生物被膜形成[16]。
1 材料与方法
1.1 材料
试验用海假交替单胞菌Pseudoalteromonasmarina(ECSMB14103) 为上海海洋大学贝类分子生物学实验室(以下简称本实验室)的保种菌株(保藏号为MCCC 1K03544)。该菌株来自浙江省嵊泗县枸杞岛(122°46′E、30°43′N)的自然生物被膜表面,使用Zobell 2216E平板划线进行培育,再分离菌株得到单一菌株,利用甘油-生理盐水保种液保种,在-80 ℃超低温冰箱中保存。
自然海水(nature seawater,NSW)取自浙江省嵊泗县枸杞岛附近,经测定钙离子浓度为11.78 mmol/L。考虑到要调节海水中的钙离子含量,用自然海水无法减少其中钙离子的含量,故本试验中配制与自然海水成分最为相似的人工海水(artificial seawater,ASW),以此来达到人为调控海水钙离子浓度的目的。试验设置钙离子浓度为10 mmol/L的对照组,以及钙离子浓度为0、1、5、20、50 mmol/L的5个试验组。人工海水的配方如下:蒸馏水、NaCl、MgSO4·7H2O、MgCl2·6H2O、KCl、CaCl2、NaNO3、K3PO4、β-五水甘油磷酸二钠盐、Na2SiO3·9H2O、维生素、pⅡMetals、S2Metals、Tris、氨基三乙酸。
试验用厚壳贻贝稚贝取自浙江省舟山市嵊泗县(122°46′E、30°69′N),在本实验室用自然海水暂养一周后使用。挑选壳长为(0.56±0.03)mm、壳高为(0.38 ± 0.02)mm的稚贝用于附着试验。
试验用激光共聚焦显微镜由Leica公司生产(×630倍镜)。
1.2 方法
1.2.1 海假交替单胞菌的分离 参考Yang等[17]的方法,将附着在载玻片上的自然生物被膜(浙江省嵊泗县枸杞岛)从载玻片表面刮到过滤过的海水中,将混有细菌的海水均匀涂在Zobell 2216E平板上,平板倒置在温度25 ℃且黑暗的条件下培育48 h,从中挑取目的菌落。再将选取出来的菌株反复分离纯化,最后将确定的纯种海假交替单胞菌(经Adapt Noise Immunity分析,上海美吉生物医药科技有限公司)用0.9%生理盐水配制成30%的甘油保种液保种,再放入-80 ℃超低温冰箱中保存。
1.2.2 生物被膜的制备 参考Yang等[17]的方法,用Zobell 2216E液体培养基培育纯种海假交替单胞菌,在25 ℃、黑暗条件下扩大培育16 h。用无菌海水洗涤培养的菌液3次,最后定容至50 mL悬浊菌液。使用Olympus BX-51荧光显微镜确定菌液细胞密度并在培养皿(直径64 mm×19 mm)中加入相应的菌液,再加入无菌海水使得培养皿终体积为20 mL。在18 ℃、黑暗条件下培育48 h形成试验用生物被膜。
菌液的初始细胞密度分别设定为1×106、1×107、1×108、5×108cells/mL,自然海水(NSW)与人工海水(ASW)形成生物被膜的比较试验中使用这4组密度,其余试验均选用最佳密度1×108cells/mL。
1.2.3 生物被膜细菌密度的计算 参考Yang等[17]的方法,将5%福尔马林溶液固定过的生物被膜用0.1%的吖啶橙溶液染色5 min后,晾干制片,在1 000倍荧光显微镜(Olympus BX-51)下选择30个区域计算细菌个数。计算公式为
时代向中国画家提出一个艰巨的历史课题:要在人物画创作上突破传统的观念、创作模式和表现技法,直接面向现代,表现现实生活。
生物被膜细胞密度(cells/cm2)=区域内细菌数(cells)/(区域个数×区域面积10-4cm2)。
1.2.4 厚壳贻贝稚贝附着试验 在培养皿中加入载有生物被膜的玻片,加入20 mL灭菌海水,放入10枚稚贝,每组设9个重复。在18 ℃、黑暗环境下培育,依据放入稚贝的时间分别观察12、24、48 h的附着情况,并计算附着率(%),即
附着率=载玻片上附着个数/总稚贝数×100%。
因12、24、48 h附着率结果基本相似,故本试验中仅出示24 h时的附着率结果。
1.2.5 激光共聚焦显微镜标本染色 参考González-Machado等[18]的方法,用表1的染料进行共聚焦避光染色,染色20 min,染色前后分别用0.9%生理盐水清洗生物被膜,并计算生物量体积(μm3),即
生物量体积=生物量总面积(Image J软件处理)×层厚0.2 μm。
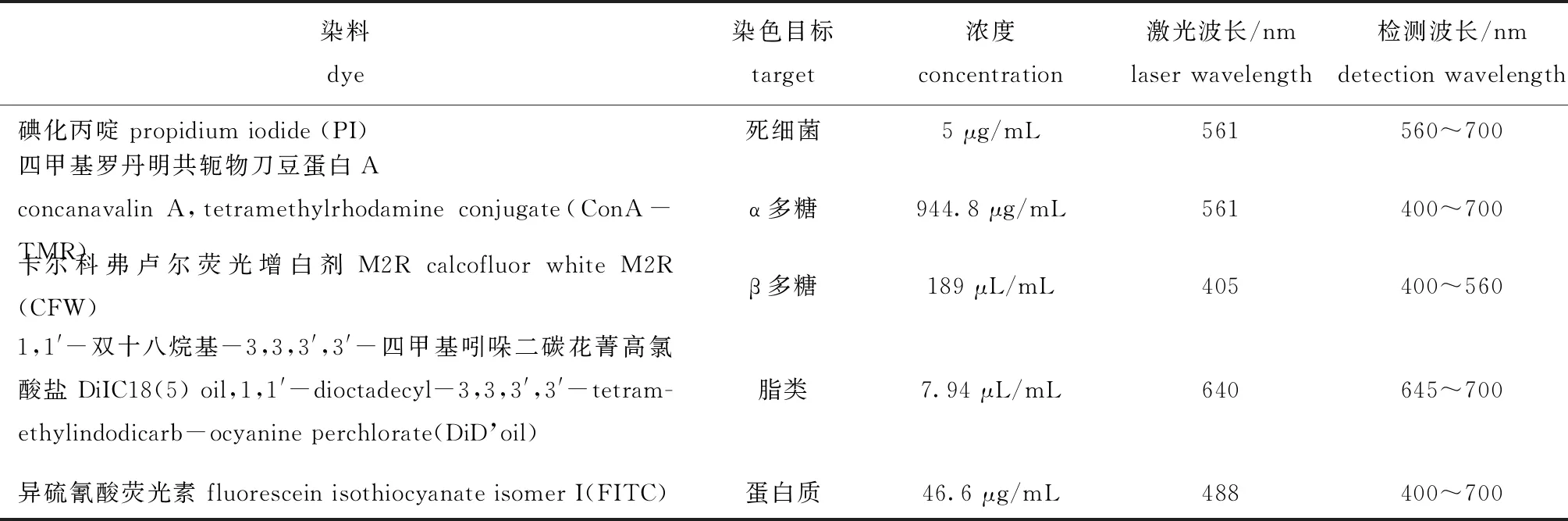
表1 激光共聚焦显微镜标本染料成分及染色对象Tab.1 Dye composition and targets for confocal laser scanning microscopy
1.3 数据处理
所有数据均使用JMP 10.0.0软件进行统计分析及相关性检验。在统计分析前,对所有数据进行正态性检测,满足正态分布,则使用单因素方差分析。通过多元分析方法验证稚贝附着率和钙离子浓度与细菌密度、膜厚、胞外产物生物量之间是否具有相关性,显著性水平设为0.05。用Image J软件处理共聚焦图像,计算胞外产物共聚焦的面积。
2 结果与分析
2.1 自然海水和人工海水形成的生物被膜细菌密度及稚贝附着率
从图1A可见:在初始菌液浓度为1×106cells/mL条件下,自然海水形成的生物被膜细菌密度为1.24×106cells/cm2,人工海水形成的生物被膜细菌密度为1.19×106cells/cm2,二者间无显著性差异(P>0.05);随着初始菌液密度的增加,自然海水和人工海水培养的生物被膜的细菌最终密度均呈现显著增加的趋势(P<0.05),且二者间终密度并无显著性差异(P>0.05)。
从图1B可见:初始菌液密度为1×106、1×107cells/mL条件下,加入稚贝24 h后自然海水和人工海水培养的生物被膜诱导的厚壳贻贝稚贝附着率与空白组的稚贝附着率(13.33%)相比,均无显著性差异(P>0.05);在初始菌液浓度为1×108cells/mL条件下,自然海水生物被膜的稚贝附着率为35.56%,人工海水生物被膜的稚贝附着率为36.67%,均显著高于前两个初始密度组(P<0.05),但自然海水与人工海水两组间稚贝附着率并无显著性差异(P>0.05);在初始菌液密度为5×108cells/mL条件下,自然海水和人工海水两组间稚贝附着率并无明显差异(P>0.05),且与1×108cells/mL密度组也无显著性差异(P>0.05)。
2.2 不同钙离子浓度下生物被膜的细菌密度
从图2可见:人工海水形成的生物被膜在菌液初始密度全部为1×108cells/mL时,海假交替单胞菌生物被膜的细菌终密度在钙离子浓度为10 mmol/L时最高,为2.53×107cells/cm2;当钙离子浓度升高或降低时,生物被膜的终密度分别呈现显著下降趋势(P<0.05),钙离子浓度为50 mmol/L时生物被膜的细菌终密度最低,仅为1.63×107cells/cm2。
2.3 不同钙离子浓度下生物被膜的稚贝附着率
从图3可见:人工海水形成的生物被膜在菌液初始密度为1×108cells/mL条件下,加入稚贝24 h后,在钙离子浓度为0、1 mmol/L时,稚贝附着率均为22.22%,与空白对照相比并无明显差异(P>0.05);随着钙离子浓度增加,稚贝附着率也呈现上升趋势,在钙离子浓度为10 mmol/L时稚贝附着率达到最大值(35.56%),随后稚贝附着率开始下降,在钙离子浓度为50 mmol/L时稚贝附着率仅为23.33%。
2.4 不同钙离子浓度下生物被膜的形态与厚度
通过用共聚焦显微镜对不同钙离子浓度下培养48 h的海假交替单胞菌生物被膜上细菌的分布形态进行观察(图4A),在钙离子浓度为0 mmol/L时,细菌的分布最为稀疏,膜厚也较薄,仅有4.00 μm;当钙离子浓度增加到10 mmol/L时,细菌开始聚集,细菌密度逐渐增加,此时膜厚达到最大值,为5.62 μm;当钙离子浓度增加到50 mmol/L时,细菌数量逐渐减少至稀疏的状态,生物被膜的厚度呈现显著降低的趋势(P<0.05)(图4B)。
2.5 不同钙离子浓度对生物被膜胞外多糖的影响
通过共聚焦显微镜观察不同钙离子浓度下培养48 h的生物被膜的胞外多糖(图5A、图6A),海假交替单胞菌生物被膜的胞外α多糖和β多糖的生物量在钙离子浓度为0 mmol/L时最少,分别为32.28、34.55 μm3;在钙离子浓度为10 mmol/L时,胞外多糖的生物量增加至最多,其中胞外α多糖的生物量为258.35 μm3,胞外β多糖的生物量为174.74 μm3;随着钙离子浓度继续升高,生物被膜的胞外多糖生物量呈现显著下降趋势(P<0.05)(图5B、图6B)。
2.6 不同钙离子浓度对生物被膜胞外蛋白的影响
通过共聚焦显微镜观察不同钙离子浓度下培养48 h的生物被膜的胞外蛋白(图7A),海假交替单胞菌生物被膜胞外蛋白的生物量在钙离子浓度为0 mmol/L时最少,为117.47 μm3;在钙离子浓度为 10 mmol/L时,胞外蛋白的生物量升高至最多,为334.15 μm3;随着钙离子浓度继续升高,生物被膜的胞外蛋白生物量呈现显著下降趋势(P<0.05),并且除钙离子浓度为10 mmol/L外,其余浓度下生物量均无显著性差异(P>0.05)(图7B)。
2.7 不同钙离子浓度对生物被膜胞外脂类的影响
通过共聚焦显微镜观察不同钙离子浓度下培养48 h生物被膜的胞外脂类(图8A),海假交替单胞菌生物被膜胞外脂类的生物量在钙离子浓度为0 mmol/L时最少,仅为11.52 μm3;在钙离子浓度10 mmol/L时,胞外脂类的生物量上升至最多,为22.02 μm3;随着钙离子浓度的继续升高,生物被膜的胞外脂类生物量呈显著下降趋势(P<0.05),并且除钙离子浓度为10 mmol/L外,其余浓度下的生物量均无显著性差异(P>0.05)(图8B)。
2.8 相关性分析
从表2可见:稚贝附着率与生物被膜细菌密度、膜厚、胞外多糖和胞外蛋白呈显著相关(P<0.05);钙离子浓度仅与生物被膜膜厚呈显著相关(P<0.05),与其他因子均无显著相关性(P>0.05)。
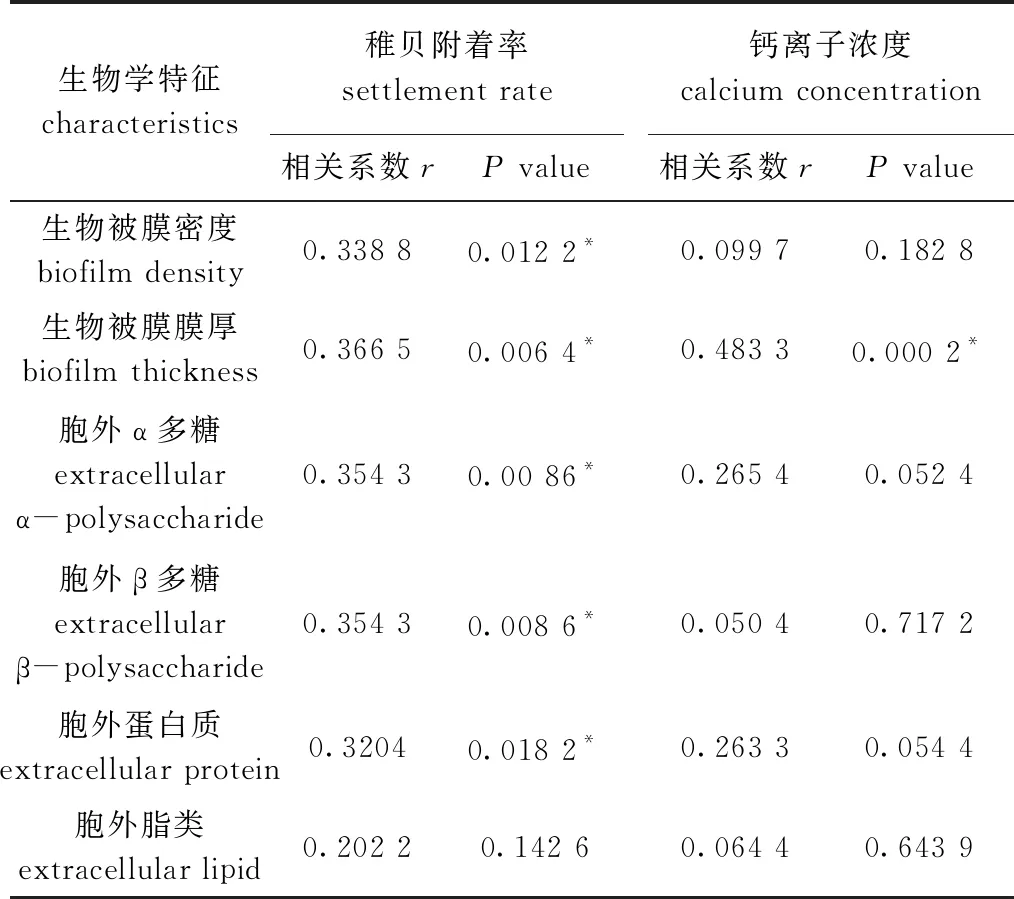
表2 生物被膜生物学特征与稚贝附着率、钙离子浓度的相关性分析
3 讨论
低钙离子浓度能显著诱导莫桑比克罗非鱼的黏液细胞增殖,以抵抗外界胁迫[19]。同时,钙离子浓度能显著影响三角帆蚌的生长[20]。但是,目前尚未有试验验证钙离子浓度对海假交替单胞菌生物被膜和厚壳贻贝稚贝附着的影响。
3.1 自然海水和人工海水的比较
由于本试验中要研究钙离子浓度变化对海假交替单胞菌生物被膜和稚贝附着的影响,需要调节钙离子的浓度,故使用人工海水来调节钙离子浓度。与自然海水相比较,本试验中使用的人工海水在培养4个不同初始菌液密度下的海假交替单胞菌生物被膜形成中未产生任何显著性影响。在4个不同初始菌液密度下,人工海水培养的细菌生物被膜的细菌密度与自然海水培养的生物被膜相比无显著性的差异,两种海水分别培养的细菌生物被膜对稚贝附着的诱导活性也均无显著性差异。多次重复试验所得到的结果相同,表明该人工海水对于细菌生物被膜的形成具有极强的稳定性。说明该人工海水可用于本试验中替代自然海水进行试验。另一方面,人类活动会造成海水中氮元素、磷元素和石油类物质超标等污染问题[21],海水养殖的品质和效益受到海水水体污染影响,海洋环境污染会导致养殖品种的质量降低。因此,未来可以考虑使用人工海水替代天然海水进行陆基人工养殖。
3.2 不同钙离子浓度对生物被膜形成的影响
通过试验可以观察到生物被膜细菌密度和生物被膜厚度在钙离子浓度为10 mmol/L时达到最大值,而在其他钙离子浓度下均降低。依据共聚焦显微镜拍摄结果观察,生物被膜在钙离子浓度为10 mmol/L时细菌呈现聚集状态,在其余钙离子浓度下细菌分散且密度减少,生物被膜形成能力变差。对假交替单胞菌Pseudoalteromonassp.的研究也有相似的结果,在钙离子浓度为10 mmol/L时,Pseudoalteromonassp.生物被膜最厚,在其余浓度下生物被膜厚度均降低[11],该试验结果与本试验结果一致。
本研究中,依据相关性分析,钙离子浓度仅与生物被膜膜厚显著相关,生物被膜膜厚受到胞外产物生物量的影响。生物被膜的形成是连续的进程,通过浮游细菌附着到表面上,然后形成生物被膜,其中细菌被包埋在由核酸、蛋白质和多糖组成的基质中[22-24]。在生物被膜中生长的细菌产生一种或多种胞外产物,它们会充当支架,将生物被膜群落的细菌维持在一起;多糖是生物被膜基质的主要组分,其有助于生物被膜的整体结构及抗性[23, 25-26]。钙离子对微生物各方面的影响,最终主要体现在生物被膜及其胞外产物的变化上[27]。在本试验中,将培养48 h的胞外产物进行共聚焦显微镜拍摄,以验证钙离子是否也影响胞外产物。生物被膜的3种胞外产物包括多糖、脂质和蛋白质,多糖和蛋白质在胞外产物中占主导地位。在钙离子浓度10 mmol/L时,海假交替单胞菌生物被膜形成的胞外产物最多,而在其余浓度下,胞外产物减少。这些试验结果可以证实,过高或过低的钙离子浓度均能影响生物被膜的形成。
3.3 不同钙离子浓度培养的生物被膜对厚壳贻贝稚贝附着的影响
许多海洋无脊椎动物幼虫的附着变态可以被生物被膜影响[7]。皱纹和半透明两个突变菌形成的假交替单胞菌生物被膜诱导厚壳贻贝幼虫的附着变态的活性显著降低,这表明具有较强的防污损能力[28]。本试验中,除10 mmol/L钙离子浓度以外,其他钙离子浓度培养的海假交替单胞菌生物被膜均抑制了厚壳贻贝稚贝的附着,即增加或减少钙离子浓度,形成的生物被膜均可以减少稚贝的附着。
依据相关性分析,稚贝附着率与生物被膜密度、膜厚、胞外多糖和胞外蛋白显著相关。参考Yang等[17]的方法,希瓦氏菌会促进厚壳贻贝幼虫的附着变态,可能与其释放的信号分子和胞外产物有关。胞外产物主要是多糖、蛋白质、核酸和脂质,其提供了生物被膜的机械稳定性,介导了与表面的黏附,构成了一个内聚的三维聚合物网络[29]。生物被膜的胞外产物在除10 mmol/L钙离子浓度以外的其他浓度下均减少,与稚贝附着率的变化趋势一致,胞外产物的减少与稚贝附着率的减少有一定相关性。
对Pseudoalteromonassp.的研究发现,10 mmol/L钙离子浓度导致其鞭毛蛋白表达量减少,外膜蛋白表达量增加,生物被膜形成能力增强[11]。大肠杆菌外膜蛋白OmpA已被证明可以结合非生物表面,OmpA在非生物表面上的生物被膜形成中起作用[30]。大肠杆菌ompA基因缺失突变菌形成的生物被膜厚度大大降低[31]。结合本试验结果推测,钙离子可能影响了海假交替单胞菌外膜蛋白的表达量,从而抑制了胞外产物的分泌,进而影响了生物被膜的形成和稚贝附着。
4 结论
1)钙离子浓度高于或低于与自然海水最接近的浓度10 mmol/L时,均抑制了海假交替单胞菌生物被膜的形成。
2)过高或过低钙离子浓度抑制了生物被膜密度、膜厚的形成,以及胞外多糖、胞外蛋白的分泌,进而抑制了厚壳贻贝稚贝的附着。
猜你喜欢
当代水产(2022年3期)2022-04-26
感染、炎症、修复(2021年1期)2021-07-28
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
红蜻蜓(2017年2期)2017-03-29
作文周刊·小学一年级版(2016年39期)2017-03-03
中学生数理化·高一版(2016年7期)2016-12-07
试题与研究·中考化学(2016年1期)2016-09-30
中学化学(2016年4期)2016-05-30
中学科技(2015年8期)2015-08-08
