
应用环介导等温扩增技术检测江苏省水稻苗期与成株期的恶苗病病菌
2020-12-14 04:18王晓莉李哲杨红福
江苏农业科学 2020年20期
关键词:苗期
王晓莉 李哲 杨红福


摘要:为了解2018年江苏省田间水稻苗期和成株期的恶苗病病菌组成,应用可特异性识别藤仓镰孢、层出镰孢、拟轮枝镰孢和Fusarium andiyazi的环介导等温扩增技术检测疑似水稻恶苗病样本47份。结果表明,苗期与成株期检测到的恶苗病病菌组成不同,29份苗期样本中检测到藤仓镰孢、层出镰孢2种水稻恶苗病病菌的检出率分别为8966%、27.59%;18份成株期样本中4种恶苗病菌均有检出,藤仓镰孢、拟轮枝镰孢、层出镰孢和F. andiyazi的检出率分别为88.89%、50.00%、27.78%、16.67%。结果表明,藤仓镰孢是江苏省水稻苗期与成株期恶苗病病原菌的优势种。
关键词:水稻恶苗病;苗期;成株期;环介导等温扩增技术
中图分类号:S435.111.4+4 文献标志码: A 文章编号:1002-1302(2020)20-0110-04
水稻恶苗病是水稻重要的病害之一,对世界各地的水稻均造成了不同程度的危害,严重可造成水稻减产50%[1]。水稻恶苗病对我国水稻也造成了不容忽视的危害,江苏省水稻种植区在不同年份均发生过恶苗病大流行[2]。水稻恶苗病在整个发育期均可发生,其中苗期和成株期是最易发病的2个时期。苗期受害的典型症状为秧苗徒长、茎细长、叶片褪绿、须根不发达,有些受害秧苗表现为褪绿或矮化,受害严重的出现整株枯死。成株期水稻受害后症状与苗期相似,也会表现徒长,有些在茎间或节间倒生不定根,根部和茎基部腐烂,严重的整株枯死[3]。水稻恶苗病病原菌主要通过种子传播[4]。由于我国尚未发现对恶苗病高抗的水稻品种,因此,目前有效的防治手段是使用化学药剂进行防治,处理的最佳时期为播种前浸种[5]。
在已知的4种水稻恶苗病病原菌中,对水稻致病力最强的是藤仓镰孢(Fusarium fujikuroi),其次是层出镰孢(F. proliferatum)、拟轮枝镰孢(F. verticilliodes)、F. andiyazi[6]。它们的寄主非常广泛,包括水稻、玉米、高粱、大豆等重要作物[7-9]。上述病原菌可分泌次生代谢物,如赤霉素、镰孢菌素和伏马菌素等。赤霉素为促生长素,可促进细胞快速伸长、细胞间隙增大,使植株节间伸长表现徒长现象;其他代谢物可引起叶片褪绿、植株矮化、死苗等症状[10]。在上述4种病原菌中,藤仓镰孢菌引起的水稻恶苗病为系统侵染,病菌在发病水稻全株组织内均有分布,其他3种病原菌为局部侵染,仅在发病植株的根部和茎基部有分布[11] 。
水稻恶苗病在江苏省稻区广有发生,但对在田间引起该病的病原菌组成及发生频率知之甚少。本研究应用笔者所在实验室研发的可特异性识别藤仓镰孢、层出镰孢、拟轮枝镰孢、F. andiyazi的环介导等温扩增(loop-mediated isothermal amplification,LAMP)检测技术对2018年来自江苏省不同地区采集的水稻苗期和成株期恶苗病疑似发病样本进行检测,以期了解当前江苏省水稻苗期和成株期引起水稻恶苗病的病原菌组成及其出现频率。旨在为该病的防治提供参考。
1 材料与方法
1.1 供试样本
2018年6月从江苏省淮安、盐城、南通、镇江、南京、常州、无锡和苏州等地采集水稻恶苗病疑似发病样本29份(表1),2018年9—10月从江苏省盐城、淮安、南通、南京、镇江、无锡、常州和苏州等地采集水稻成株期疑似恶苗病样本18份(表3)。将收集来的样本装袋,记录信息并进行编号。
1.2 病组织DNA的提取
在每份疑似稻田发病样本中选取10株发病症状明显的植株剪取其根茎部,长度为5 mm左右,放进无菌培养皿内混匀,其中一半病组织用来提取DNA,另一半用来分离病原菌。将植物组织放进灭菌的研钵中,用液氮研磨后,参照DNA提取试剂盒说明的步骤进行DNA提取,采用天根生化科技(北京)有限公司的新型植物基因组DNA提取试剂盒DNAsecure Plant Kit。提取好的DNA放-20 ℃冰箱保存。
1.3 水稻恶苗病病菌的LAMP检测
采用笔者所在实验室研发的4个分别可特异性检测藤仓镰孢、层出镰孢、拟轮枝镰孢和F. andiyazi的环介导等温扩增检测技术[12-14],以从疑似恶苗病组织提取的DNA为模板进行检测。
LAMP反应体系为2.5 μL 10×ThermoPol Buffer [0.1% Trion-X,20 mmol/L Tris-HCl,10 mmol/L KCl,10 mmol/L (NH4)SO4,pH值=88],4 μL MgSO4(50 mmol/L),4 μL 甜菜碱(5 mol/L),3.5 μL dNTPs(10 mmol/L),内引物FIP和BIP(20 μmol/L)各2 μL,外引物F3和B3(10 μmol/L)各0.5 μL,环引物LF和LB(10 μmol/L)各1 μL,2 μL HNB(2.4 mmol/L),1 μL Bst DNA 聚合酶(8 U/μL)以及2 μL模板(从恶苗病组织中提取)DNA。由反应结束后的颜色直接判定结果,蓝色为阳性,紫色为阴性。
1.4 水稻恶苗病病原菌的分离与验证
为进一步验证水稻恶苗病病菌的LAMP检测结果,选取部分LAMP结果呈阳性的水稻恶苗病样本的根茎部进行病原菌分離。用70%乙醇消毒 30 s,2%次氯酸钠消毒2.5 min,无菌水冲洗3次,用灭菌滤纸片吸去表面自由水后放置于马铃薯葡萄糖琼脂培养基(PDA)培养基平板上,在25 ℃黑暗条件下培养48 h。从长出的菌落边缘切取2 mm×2 mm菌丝块移至新的PDA培养基上纯化。纯化后的分离物根据形态和碱基序列比对进行鉴定。分离物DNA提取采用天根生化科技(北京)有限公司的新型植物基因组DNA提取试剂盒,通过扩增TEF-1α基因序列及序列比对进行恶苗病病菌菌种的鉴定。
2 结果与分析
2.1 苗期水稻疑似恶苗病样本的LAMP检测结果
4个LAMP检测体系对来自苏州市凤凰镇的武运粳30稻苗的疑似发病样本的检测结果如图1所示。结果表明,可特异性识别藤仓镰孢(4号反应管)和层出镰孢(5号反应管)的LAMP检测结果呈阳性,判定武运粳30稻苗发病样本中包含藤仓镰孢和层出镰孢2种水稻恶苗病病菌,说明该样本混合有由藤仓镰孢或层出镰孢侵染发病的病苗,或是由两者复合侵染引起的。其他2种病原菌(2号和3号反应管)的LAMP检测结果呈阴性,表明武运粳30的样本中不含拟轮枝镰孢和F. andiyazi。
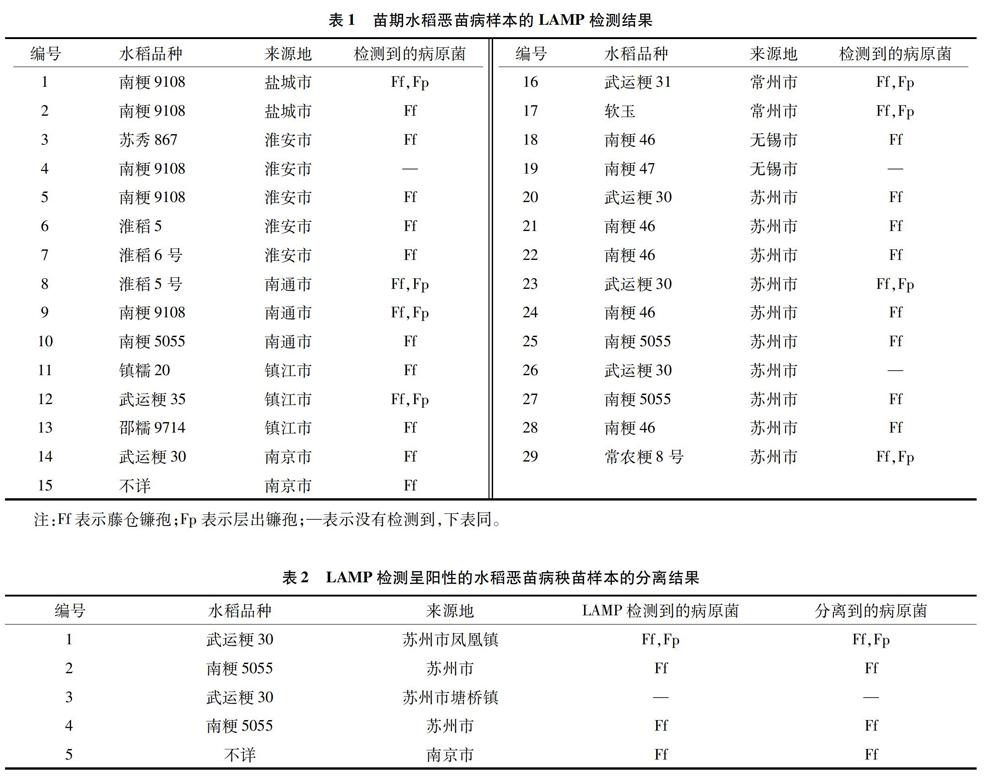
供试29份苗期水稻恶苗病疑似样本的检测结果(表1)显示,有26份检测到藤仓镰孢,检出率达89.66%;8份样本检出层出镰孢,检出率为2759%;有3份未检测出恶苗病病菌。
在上述26份阳性样本中,有18份仅检测到藤仓镰孢,说明上述发病样本是由该病原菌单独侵染的;另有8份样本同时检测到藤仓镰孢和层出镰孢,说明这8份发病样本由两者复合侵染引起。结果表明,藤仓镰孢是2018年江苏省水稻苗期恶苗病的优势病原菌。
2.2 苗期样本的病原菌分离
为验证苗期LAMP检测结果的可靠性,选择5份LAMP检测阳性的秧苗样本进行分离,包括武运粳30、2份来自苏州市不同地域的南粳5055、南京市的1个未知品种,以及1份未检测到恶苗病病菌的武运粳30稻苗样本(表2)。分离结果表明,与LAMP的检测结果一致,说明本研究的LAMP检测结果是可靠的。
2.3 成株期疑似水稻恶苗病样本的LAMP检测结果与分离
由表3可知,有16份样本检测出藤仓镰孢,检出率达88.89%;拟轮枝镰孢、层出镰孢和F. andiyazi的阳性样本数分别为9、5、3份,检出率依次为50.00%、27.78%、16.67%。与苗期样本的检出率相比,拟轮枝镰孢、层出镰孢、F. andiyazi的检出率有所升高。
在16份LAMP检测阳性样本中,有7个样本仅检测出藤仓镰孢,说明这7份样本是由藤仓镰孢单独侵染引起发病。另有9份样本中检测出2种或2种以上水稻恶苗病病菌,占总样本的50.00%,表明这些发病样本可能是由2种及2种以上恶苗病病菌复合侵染引起。在上述9份样本中有2份样本检出4种恶苗病病菌,4份样本中检出3种恶苗病病菌(其中检出藤仓镰孢、层出镰孢、拟轮枝镰孢的样本有3份,检出藤仓镰孢、拟轮枝镰孢、F. andiyazi的有1份),3份样本检测出2种恶苗病病菌(均为藤仓镰孢和拟轮枝镰孢)。在供测的所有样本中,未发现由层出镰孢、拟轮枝镰孢或F. andiyazi单独侵染发病的样本。
进一步对检测阳性样本进行病原菌分离,有14份样本分离出藤仓镰孢,7份样本分离出层出镰孢,1份样本分离出F. andiyazi,所有样本均未分离到拟轮枝镰孢。需要指出的是,7份分离出层出镰孢的样本亦同时分离出藤仓镰孢。病原菌分离与鉴定结果表明,藤仓镰孢是江苏省水稻成株期引起水稻恶苗病的优势种,其他3种恶苗病病菌亦有不同程度出现,但仅起辅助作用。关于拟轮枝镰孢检出率较高(50.00%)而分离率为零的原因尚不清楚。可能原因之一为藤仓镰孢为强势致病菌株,而拟轮枝镰孢可能仅起伴随侵染作用,并在发病组织中处极弱势地位。
3 结论与讨论
對2018年采集自江苏省8个不同地区的47份水稻恶苗病疑似样本进行LAMP检测的结果表明,已知的4种恶苗病病菌在江苏省均有分布,其中藤仓镰孢是当前江苏省稻区引起水稻恶苗病的优势种,其他3种恶苗病病菌也有检测到,但罕有单独侵染引起恶苗病的病例,而似多与藤仓镰孢复合侵染引起发病,提示上述3种病原菌在江苏省水稻恶苗病的发生中可能起辅助作用。本研究结果对了解当前江苏省稻区水稻恶苗病病原菌组成具有重要价值,也可为该病的防治提供参考。
LAMP检测技术是21世纪初建立的一种快速分子检测技术,与传统PCR方法相比,该检测技术特异性更强、灵敏度更高(是传统PCR的10~100倍)、检测所需时间更短(完成1个样本的检测仅需2 h左右)。该检测技术不需要昂贵的仪器设备,对模板DNA的质量要求不高,可以从发病组织提取的DNA中特异性识别目标病原菌的靶标序列,因此可以在快速诊断病害的同时完成病原菌的鉴定[15]。本研究采用的4个恶苗病病菌的LAMP检测技术体系,不仅可以用于恶苗病的快速诊断,也可用于种子携带恶苗病病菌的检测和土壤带菌检测[11]。
种子携带恶苗病病菌是该病的重要传播途径。有报道指出,水稻种子主要携带藤仓镰孢和层出镰孢[16],本研究对发病秧苗的检测结果与袁咏天等的研究结果[16]基本一致。该病在大田成株期的病原菌组成较苗期复杂,提示拟轮枝镰孢和F. andiyazi的主要侵染可能来自秧苗移植后的稻田田间。
参考文献:
[1]Desjardins A E,Manandhar H K,Plattner R D,et al. Fusarium species from nepalese rice and production of mycotoxins and gibberellic acid by selected species[J]. Applied and Environmental Microbiology,2000,66(3):1020-1025.
[2]秦玉金,刘学儒,杨 进,等. 水稻恶苗病重发原因浅析及防治对策[J]. 上海农业科技,2013(5):111,95.
[3]韩焕忠,朱 萍,王晓艳. 青冈县水稻常见病害的发病症状及防治措施[J]. 现代农业科技,2018(1):120,123.
[4]Matic S,Gullino M L,Spadaro D. The puzzle of bakanae disease through interactions between Fusarium fujikuroi and rice[J]. Frontiers in Bioscience,2017,9(2):333-344.
[5]彭小林,王 军,王远坚. 2016年永新县水稻恶苗病的发生特点及防治对策[J]. 现代农业科技,2018(8):128,130.
[6]Wulff E G,Sürensen J L,Lübeck M,et al. Fusarium spp. associated with rice Bakanae:ecology,genetic diversity,pathogenicity and toxigenicity[J]. Environmental Microbiology,2010,12(3):649-657.
[7]任 旭,朱振东,李洪杰,等. 轮枝镰孢SSR标记开发及在玉米分离群体遗传多样性分析中的应用[J]. 中国农业科学,2012,45(1):52-66.
[8]Arias M M,Munkvold G P,Leandro L F. First report of Fusarium proliferatum causing root rot on soybean (Glycine max) in the United States[J]. Plant Disease,2011,95(10):1316.
[9]Marasas W O,Rheeder J P,Lamprecht S C,et al. Fusarium andiyazi sp. nov.,a new species from sorghum[J]. Mycologia,2001,93(6):1203-1210.
[10]Niehaus E M,Kim H K,Münsterktter M,et al. Comparative genomics of geographically distant Fusarium fujikuroi isolates revealed two distinct pathotypes correlating with secondary metabolite profiles[J]. PLoS Pathogens,2017,13(10):e1006670.
[11]戎振陽. 应用环介导等温扩增技术快速诊断水稻恶苗病[D]. 南京:南京农业大学,2018:43-52.
[12]Rong Z Y,Yuan Y T,Ye W W,et al. Rapid diagnosis of rice bakanae caused by Fusarium fujikuroi and F. proliferatum using loop-mediated isothermal amplification assays[J]. Journal of Phytopathology,2018,166(4):283-290.
[13]曾丹丹,戎振洋,袁咏天,等. LAMP快速诊断由拟轮枝镰孢引起的水稻恶苗病[J]. 南京农业大学学报,2018,41(2):286-292.
[14]戎振洋,袁咏天,曾丹丹,等. 基于环介导等温扩增技术快速诊断由Fusarium andiyazi引起的水稻恶苗病[J]. 植物病理学报,2018,48(2):256-262.
[15]王芝涵,王春伟,高海馨,等. 引起玉米穗腐病的禾谷镰刀菌LAMP快速检测方法的建立[J]. 江苏农业学报,2019,35(3):581-585.
[16]袁咏天,戎振洋,叶文武,等. 应用环介导等温扩增技术检测江苏水稻种子携带的水稻恶苗病菌[J]. 中国水稻科学,2018,32(5):493-500.宋兆欣,曹坳程,李 林,等. 50%草除灵悬浮剂防除稻旱轮作油菜田1年生阔叶杂草及其作物安全性评价[J]. 江苏农业科学,2020,48(20):114-117.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
今日农业(2021年9期)2021-11-26
今日农业(2020年23期)2020-12-15
今日农业(2019年12期)2019-08-13
山东林业科技(2019年2期)2019-06-03
现代园艺(2018年3期)2018-02-10
浙江农业学报(2016年7期)2016-06-15
广西林业科学(2016年3期)2016-03-16
作物研究(2014年6期)2014-03-01
植物营养与肥料学报(2012年5期)2012-10-26
