
PEG处理下3种耧斗菜属植物萌发耐旱性评价
2020-11-26 06:49王冯熠李家绮周蕴薇陈丽飞
福建农林大学学报(自然科学版) 2020年6期
孟 缘,王冯熠,李家绮,白 云,周蕴薇,陈丽飞
(吉林农业大学园艺学院,吉林 长春 130118)
近年来东北地区年降水量、降水日数呈下降趋势,降水分布不均,旱灾发生概率明显增加,导致干旱成为制约东北农业生产、景观绿化发展的重要因素之一[1-2].同时东北播种时节正值春季,气候干燥少雨,风沙较大,土壤表层易风干失水,导致土壤容重下降[3],确定观赏植物耐旱耐极限以及筛选抗性植物材料对改善东北地区的生态环境具有重要意义.
PEG(聚乙二醇)是一类高分子渗透剂,本身不易自由通过植物细胞壁进入种子细胞内;PEG可调节种子细胞渗透压,减缓种子萌发时水分进入种子的速度,防止种子在吸胀初始时期因吸水过快导致细胞膜受损,缩短种子的修复时间,以达到促进种子萌发的目的[4].PEG还可调节细胞代谢,具有条件可控、操作简单、易重复等优点[5].如经10%的PEG处理下3个大豆品种的发芽率和发芽势显著高于对照组[6];15%~20%的PEG处理下朝天椒的发芽率、发芽势、发芽指数均升高[7];赛菊芋种子在5%PEG处理下的发芽势、活力指数和胚根长度均为各处理组中最高值[7].PEG水溶液常作为植物耐旱性选择剂[8],目前已广泛用于经济作物[9-10]、园艺植物[11-12]的耐旱性评价及筛选.
耧斗菜属(AquilegiaL.)为多年生宿根草本植物,隶属于毛茛科.原产地位于北美洲和欧洲,在北半球国家广泛分布.耧斗菜属植物株型优美、花型奇特、野趣盎然,是庭院与城市景观的优秀观赏花卉[13].目前对于耧斗菜属植物逆境萌发的研究较少,仅限于黄凤耧斗菜、白鹭耧斗菜[14]、耧斗菜矮重品种和矮化品种[15]的PEG胁迫研究,对北方地区常见的耧斗菜属植物萌发抗旱性评价和筛选未见报道.本研究以北方地区常见的以播种繁殖为主要繁殖途径的3种耧斗菜属植物为研究对象,包括尖萼耧斗菜(A.oxysepala)、小花耧斗菜(A.parviflora)、耧斗菜(A.viridiflora),经不同浓度的PEG-6000溶液,测定各项萌发指标,对萌发情况和耐旱能力进行评价,以期为耧斗菜属植物在北方的推广应用提供依据.
1 材料与方法
1.1 试验材料
本试验于2018年12月在吉林农业大学园艺学院人工气候培养箱内进行,供试材料小花耧斗菜、尖萼耧斗菜、耧斗菜种子购于江苏省沭阳县思途园林绿化苗木场.
1.2 试验设计
选取颗粒饱满的3种耧斗菜种子,用0.1% KMnO4溶液浸泡消毒5 min,消毒后用蒸馏水冲洗3~5次,直至无KMnO4残留为止,室温晾干备用.将PEG-6000设置6个浓度水平,分别为0%(CK)、5%、10%、15%、20%、25%.发芽试验在9 cm的培养皿中进行,每个培养皿铺2层滤纸,并加入5 mL的处理液.每个培养皿放置30粒种子,每品种每处理重复3次.
将人工气候箱设置为12 h白天/12 h黑夜,20 ℃恒温.每隔24 h统计一次出芽率,以胚根长度≥种子直径的2倍作为发芽的标准计数,从游标卡尺测定子叶基部与根部分生处到根尖的长度为胚根长度(mm),各处理的种子发芽数量连续2 d无变化时视为试验结束.每天用称重法补充适量蒸馏水,以维持各梯度浓度稳定[7].
1.3 指标计算方法和抗旱能力综合评价
1.3.1 指标计算 日相对发芽率/%=单日正常萌发种子个数/供试种子数量×100;最终发芽率/%=试验结束时萌发的种子数/供试种子数×100;发芽势/%=(种子发芽数达到高峰时的发芽数量/供试种子数)×100;发芽指数(Gi)=Σ(Gt/Dt)×100,Gt位第t天发芽的种子数,Dt为相应的发芽天数;种子活力指数(Vi)=Gi×S,S为胚根长度.
1.3.2 耐旱临界浓度和耐旱极限浓度 对种子发芽率和PEG浓度进行回归分析,根据回归方程计算出最终发芽率是对照处理50%和0%时的PEG浓度,即相应的耐旱临界浓度和耐旱极限浓度[8].
1.3.3 抗旱性综合评价 采用模糊数学中隶属函数[18]的方法,对3种耧斗菜种子萌发时的抗旱性进行综合评价,计算方法为:
Yij=(Xij-Ximin)/(Ximax-Ximin)
(1)
Yij=1-(Xij-Ximin)/(Ximax-Ximin)
(2)
式中:Xij为i种j的指标值;Xjmin,试验种类中j指标中的最小值;Xjmax为试验种类品种中j指标中的最大值;Yij为i种类j指标的抗性隶属值.当测定指标与抗旱性成正比时使用公式(1),当测定指标与抗旱性呈反比时使用公式(2).
1.4 数据处理
数据采用SPSS 20进行统计分析,方差分析采用单因素方差分析法和最小显著差异法,显著性水平设定为P<0.05.采用Excel 2010制图.
2 结果与分析
2.1 PEG处理对3种耧斗菜种子萌发进程的影响
由图1可知,不同浓度PEG处理下,3种耧斗菜种子的萌发进程均受到不同程度的影响.3种耧斗菜种子最终发芽率下降,耧斗菜与小花耧斗菜的萌发起始时间延迟.由图2可知,耧斗菜种子各对照组的萌发起始时间为第11天;尖萼耧斗菜、小花耧斗菜、耧斗菜的萌发高峰分别在第12、15、13天;在5%~15%PEG处理下,小花耧斗菜、耧斗菜种子的萌发高峰延迟;当PEG浓度达到20%时,尖萼耧斗菜、小花耧斗菜和耧斗菜的萌发高峰时间分别为第19、18、18天,比对照组延迟了7、3和5 d;当PEG浓度为25%时,3种耧斗菜种子均无法正常萌发.

图1 不同浓度PEG处理对3种耧斗菜种子萌发进程的影响Fig.1 Effects of PEG treatments on the germination process of 3 Aquilegia species
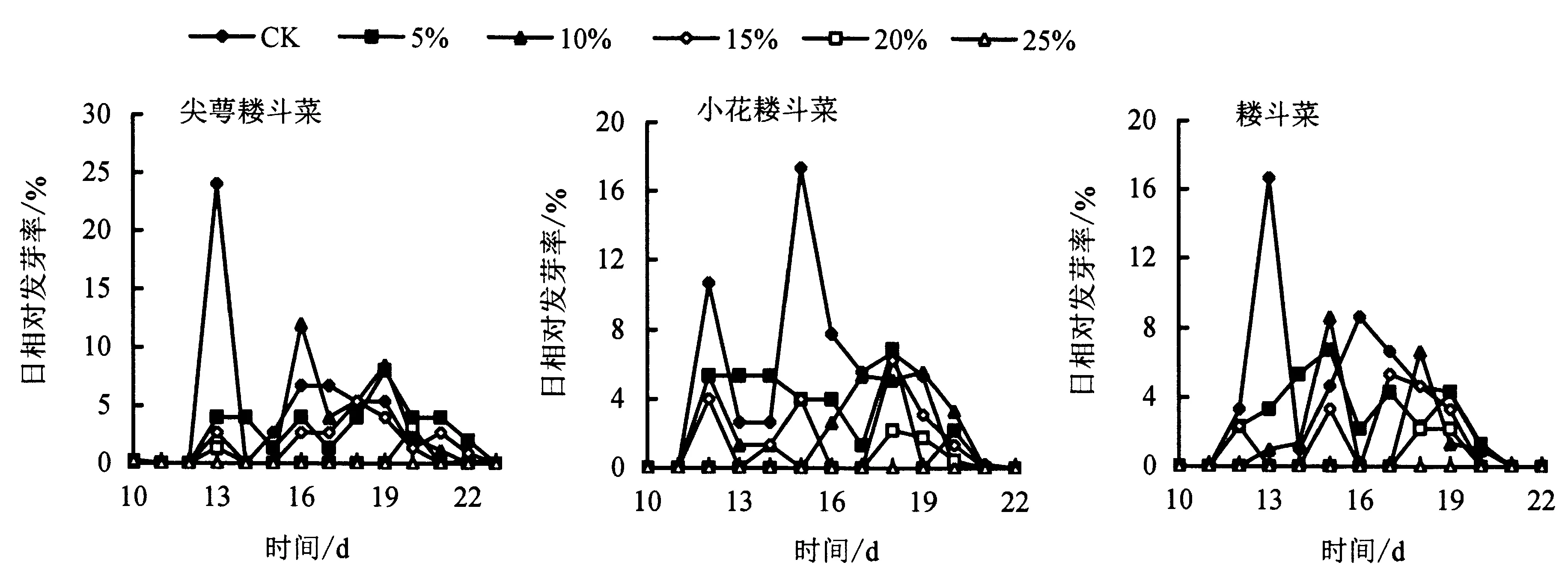
图2 不同浓度PEG处理对3种耧斗菜种子日相对发芽率的影响Fig.2 Effects of PEG treatments on relative daily germination rate of 3 Aquilegia species
2.2 PEG处理对3种耧斗菜种子最终发芽率和发芽势的影响
由表1可知,PEG处理下3种耧斗菜最终发芽率均受到不同程度的抑制.各种类对照组最终发芽率均最高,种间无显著差异.当PEG浓度为5%时,相比对照组,降幅从大到小依次为小花耧斗菜>耧斗菜>尖萼耧斗菜,分别下降了41.52%、36.96%和29.78%.方差分析表明,3种耧斗菜最终发芽率均显著下降,种间差异不显著.当PEG浓度为10%时,3种耧斗菜最终发芽率均下降,耧斗菜最终发芽率显著低于尖萼耧斗菜,与小花耧斗菜差异不显著;当PEG浓度为15%~20%时,3种耧斗菜的发芽率均大幅下降,各处理组种间差异不显著;当PEG浓度为25%时,3种耧斗菜最终发芽率均为0.
随着PEG浓度的上升,3种耧斗菜的发芽势总体受到抑制(表1).对照组小花耧斗菜的发芽势显著高于其他两个种类.尖萼耧斗菜的发芽势随PEG浓度呈先上升后降低趋势,而小花耧斗菜、耧斗菜的发芽势则持续下降;当PEG浓度为10%时,相比对照组,尖萼耧斗菜发芽势增加了27.29%,小花耧斗菜、耧斗菜的发芽势分别下降了48.65%和44.14%.方差分析表明,各处理组之间只有小花耧斗菜的发芽势与对照差异显著,各物种之间尖萼耧斗菜的发芽势显著高于耧斗菜.当PEG浓度为15%~20%时,3种耧斗菜种子的发芽势持续下降,各处理组种间差异不显著.当PEG浓度为25%时,3种耧斗菜种子的发芽势均为0.

表1 PEG处理对3种耧斗菜种子萌发指标的影响1)Table 1 Effects of PEG treatments on the germination indexes of 3 Aquilegia species
2.3 PEG处理对3种耧斗菜种子发芽指数和活力指数的影响
由表1可知,3种耧斗菜的发芽指数随着PEG浓度的升高而降低.对照组各种类发芽指数均为所有处理最大值,种间无显著差异.当PEG浓度为5%时,相比对照组,3种耧斗菜的发芽指数降幅从大到小依次为:小花耧斗菜>尖萼耧斗菜>耧斗菜,分别下降了40.17%、39.29%和38.24%,方差分析表明,3种耧斗菜的发芽指数与对照相比均显著下降,种间差异不显著.当PEG浓度为10%~20%时,3种耧斗菜发芽指数持续下降,各处理组种间差异不显著;当PEG浓度为25%时,3种耧斗菜发芽指数均为0.
随着PEG浓度的升高,3种耧斗菜种子活力指数均降低(表1),不同种类降低程度不同.对照组3种耧斗菜的活力指数均为所有处理的最大值,其中耧斗菜的活力指数显著低于其他两个种类.当PEG浓度为5%时,相比对照组,3种耧斗菜活力指数降幅从大到小依次为:尖萼耧斗菜>小花耧斗菜>耧斗菜,分别下降了53.88%、48.67%和35.28%,方差分析表明,只有耧斗菜的活力指数与对照组相比差异不显著,种间无显著差异.当PEG浓度为10%~20%时,3种耧斗菜活力指数均进一步受到抑制,各处理组种间无显著差异;当PEG浓度为25%时,3种耧斗菜活力指数均为0.
2.4 PEG处理对3种耧斗菜种子胚根长度的影响
由表1、图3可知,随着PEG浓度的上升,3种耧斗菜种子的胚根长度均受到不同程度的抑制.对照组尖萼耧斗菜胚根长度显著高于其他两个种类.当PEG浓度为5%时,相比对照组,尖萼耧斗菜与小花耧斗菜的胚根长度分别下降了24.73%和11.74%,耧斗菜的胚根长度增加了5.29%,方差分析表明,只有尖萼耧斗菜的胚根长度与对照相比差异显著,种间无显著差异.当PEG浓度为10%~20%时,3种耧斗菜的胚根长度持续下降,各处理组种间差异不显著;当PEG浓度为25%时,3种耧斗菜的胚根长度均为0.
2.5 3种耧斗菜种子的PEG耐旱临界浓度和极限浓度
以PEG浓度(%)为自变量x,种子最终发芽率为因变量y作回归分析,回归方程、PEG耐旱临界浓度和耐旱极限浓度如表2所示.3种耧斗菜最终发芽率与PEG浓度回归方程R2均>0.8,拟合度高.3种耧斗菜种子PEG临界浓度从大到小为:尖萼耧斗菜>小花耧斗菜>耧斗菜,分别是11.98%、10.36%和10.21%;PEG极限浓度从大到小为:尖萼耧斗菜>小花耧斗菜>耧斗菜,分别是24.33%、23.43%和23.36%,其中尖萼耧斗菜的PEG耐旱临界浓度、极限浓度均最高,而耧斗菜的PEG耐旱临界浓度和极限浓度均最低.

A:尖萼耧斗菜;B:小花耧斗菜;C:耧斗菜;a~f分别表示CK、5%、10%、15%、20%、25%PEG处理组.图3 不同浓度PEG处理20 d时3种耧斗菜胚根萌发情况Fig.3 Radicle germination of 3 Aquilegia species under different concentrations of PEG for 20 d
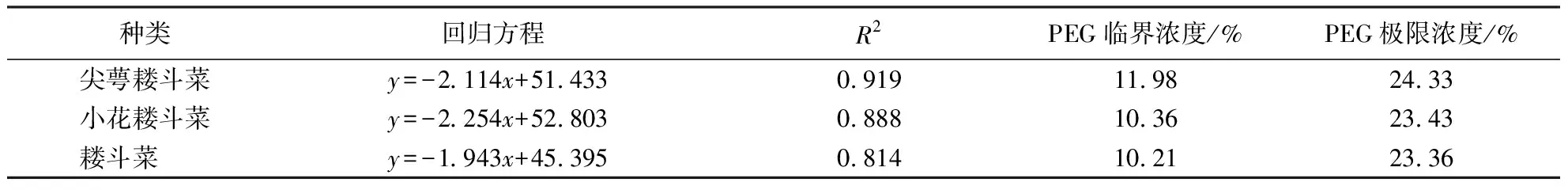
种类回归方程R2PEG临界浓度/%PEG极限浓度/%尖萼耧斗菜y=-2.114x+51.4330.91911.9824.33小花耧斗菜y=-2.254x+52.8030.88810.3623.43耧斗菜 y=-1.943x+45.3950.81410.2123.36
2.6 双因素差异比较与抗旱性综合评价
不同浓度PEG与不同种类双因素方差分析结果如表3所示.PEG处理对3种耧斗菜的发芽率、发芽势、发芽指数、活力指数、胚根长度影响极显著(P<0.01),不同种类对发芽势和胚根长度影响显著(P<0.05),而PEG浓度与不同种类间的交互作用对萌发的各项指标影响不显著.

表3 PEG浓度、种类和两者交互对3种耧斗菜萌发影响的双因素分析Table 3 Two-factor analysis of variance on PEG concentration and species on the germination of 3 species of Aquilegia
3种耧斗菜种子在不同浓度PEG处理下各项萌发指标隶属函数处理如表4.尖萼耧斗菜、小花耧斗菜和耧斗菜的平均隶属函数值分别为0.394、0.389和0.292,因此3种耧斗菜种子的抗旱性由强到弱依次为:尖萼耧斗菜>小花耧斗菜>耧斗菜.

表4 PEG处理下3种耧斗菜种子萌发指标的隶属函数值Table 4 The membership function values of seed germination index in 3 Aquilegia species under PEG treatment
3 讨论
研究表明,一定浓度的PEG可通过启动种子体内的保护机制,避免种子吸胀过快导致细胞膜损伤,促进种子萌发[4-6].本试验中,尖萼耧斗菜的发芽势在5%~10%的PEG处理下有所升高,耧斗菜的胚根长度在5%处理下相比对照组稍有增加,但与对照差异均不显著;其余指标(最终发芽率、发芽指数、活力指数)大幅下降,PEG处理下3种耧斗菜萌发总体受到抑制,与对黄凤耧斗菜、白鹭耧斗菜[5]、耧斗菜矮重矮化品种[6]以及葛缕子[15]的研究结果一致.可能是这3种耧斗菜对PEG胁迫较敏感,PEG对其萌发无刺激效应,或者在本试验所设定的浓度未对3种耧斗菜种子在吸胀初始时期产生有效的保护作用.
不同基因型植物对PEG胁迫的响应程度不尽相同,如在对58种豇豆的研究中,'C28'和'C48'在-0.075 MPa处理下,发芽率比对照组下降了约47%,而'C45'的发芽率提高了60%[20].在对10种埃及大麦栽培种的研究中,耐旱性较强的品种'Giza134'在5%PEG处理下发芽率仅降低了10%,相对不耐旱品种'Giza 124'发芽率降幅达48.11%[21].在对5种苜蓿的研究中,25%PEG处理下耐旱性较弱的品种发芽率降幅达98.7%,而耐旱性较强品种甘农三号的降幅为60.8%[22].在本试验中,3种耧斗菜的萌发均受到PEG处理的抑制,尖萼耧斗菜、小花耧斗菜和耧斗菜的耐旱极限浓度分别为24.33%、23.43%和23.36%.鉴于这3种耧斗菜萌发期对干旱条件较为敏感,在东北地区春季播种后应确保足够的水分供应.
基于PEG浓度和种类的双因素分析,PEG浓度对3种耧斗菜的萌发影响最显著,不同种类对3种耧斗菜发芽势和胚根长度影响显著.单用某一种指标往往难以全面有效地评价种间的抗旱性,综合评价可以更全面客观地反映植物抗性;隶属函数法是对植物抗性进行综合评价的有效方法之一,在对菊科植物[18]、白三叶[23]和花海花卉[24]的研究中均采用这方法来评价植物抗性.本研究采用隶属函数对3种耧斗菜萌发耐旱性进行评价,得出3种耧斗菜种子的抗旱性由强到弱依次为:尖萼耧斗菜>小花耧斗菜>耧斗菜.
猜你喜欢
小学生学习指导(当代教科研)(2021年6期)2021-05-23
黄河之声(2021年2期)2021-03-29
学生天地(2020年11期)2020-08-25
农业与技术(2019年19期)2019-11-11
热带农业科学(2019年2期)2019-04-28
农民致富之友(2018年9期)2018-06-27
现代农业研究(2017年3期)2017-04-17
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年11期)2016-01-27
吉林农业(2014年5期)2014-07-09
