
毛竹入侵对常绿阔叶林碳储量的影响
2020-11-26 06:49池鑫晨王晓雨王懿祥叶静雯朱向涛白尚斌
福建农林大学学报(自然科学版) 2020年6期
宋 超,池鑫晨,王晓雨,王懿祥,叶静雯,朱向涛,白尚斌,
(1.浙江农林大学林业与生物技术学院;2.浙江省森林生态系统碳循环及固碳减排重点实验室,浙江 临安 311300;3.暨阳学院,浙江 诸暨 311800)
森林是重要的陆地生态系统,其植被部分碳库占全球植被碳库的77.1%[1],森林土壤碳库储存了全球土壤碳储量的约40%[2,3].因此,森林在增加碳汇和减缓大气CO2浓度升高方面所发挥的作用越来越突出[4-6].大多数研究表明[7-12],森林碳储量及固碳能力与植被类型、物种组成、森林生产力、林龄及人类活动等有着密切的关系.以往研究广泛认同,植物入侵对森林生态系统的结构和功能有负面影响.但是,在森林碳储量方面,入侵植物因其快速生长和繁殖的特性,可能在短时期维持较高的生长速率,如果入侵群落的快速生长能够抵消原有群落的碳损失,则区域水平上,入侵过程可能不会导致碳损失,甚至可能引起碳储量在短时期内升高.根据以往研究,不同森林群落对入侵的反应可能有较大差异,入侵植物的不同,以及被入侵生态系统的差异,使得植物入侵对原有植被碳储量的影响具有复杂性.例如,木本植物乌桕(Sapiumsebiferum)从中国引进美国后,由于其生长速度快、繁殖能力强、抗逆性强的特性,相对原生种群表现出很强的竞争能力[13],沿海岸线快速扩张,最终导致原群落物种多样性及生物量下降[14];但在入侵成功后,乌桕的地上及地下部分碳储量均大于原产地种群[15].又如柽柳(Tamrixspp.),入侵美国水系附近的多年生植被群落,导致原群落的生物量降低[16].而桉树(EucalyptusrobustaSmith)引进我国广东、云南等地后,虽然被列为入侵植物之一,但是相比原有植被有着更高的碳储量[17].
毛竹(Phyllostachyedulis)是原产于我国的禾本科刚竹属竹种,广泛分布在我国热带和亚热带地区,毛竹具有极高的经济价值,其茎秆、竹笋皆可利用,并且具有生长速度快、产量高等经济优势[18],竹林经营收入曾是山区收入的主要来源之一,竹产业也是竹农发展经济的主要产业,是我国一种重要的森林资源[19].然而近几年来,竹产业遭受产业结构转型升级、环保高压等种种压力的严峻考验,导致毛竹竹材价格急剧下降,加上劳动力成本升高,农民竹林经营收益较低甚至亏损.越来越多的毛竹林被减少或放弃经营,甚至成为完全抛荒的竹林.
毛竹具有强大的竹鞭系统,可以向邻近常绿阔叶林、针阔混交林等森林蔓延入侵,并快速蔓延蚕食其他树种[20],形成竹-阔混交林甚至是毛竹纯林[21].入侵后,由于群落结构、土壤性质改变以及毛竹自身无法二次生长等原因[22],不可避免地会影响原有森林碳储量.杨清培等[23]的研究发现,毛竹入侵常绿阔叶林后,总碳储量、植被碳储量和土壤碳储量等均在降低,但植被年固碳量却上升.而赵雨虹[20]等研究结果表明毛竹入侵常绿阔叶林导致森林生态功能的综合水平呈现下降趋势,竹阔混交林的碳储量却大于毛竹林和阔叶林.但是由于研究区气候、土壤、林分以及研究方法等的差异,关于毛竹入侵对阔叶林生态系统碳储量变化的影响还具有不确定性.
毛竹生长迅速、固碳能力强,被视为应对全球变化的重要树种大力发展.而阔叶林生态系统是亚热带重要的地带性植被,发挥着重要的固碳功能.那么当毛竹扩张进入阔叶林后,对原生植被碳储量会带来怎样的影响?生态系统碳储量分配格局又会有怎样的改变?研究结果对人们正确认识全球变化背景下毛竹林作用以及合理调控其入侵有重要的理论和实践意义.基于此,本研究以杭州午潮山林场毛竹向常绿阔叶林入侵的区域为研究对象,通过设立长期固定监测样地(毛竹纯林、毛竹阔叶混交林、原生常绿阔叶林),研究常绿阔叶林在毛竹扩张后引起的生物量及碳储量变化,研究结果对估算区域森林生物量及碳储量有重要意义.
1 研究区概况
研究区位于浙江省杭州市午潮山试验林场(33°41′N″—120°0′E″),该区域属于亚热带季风气候区,四季分明,雨量充沛.全年最高温度38.6 ℃,最低温度3.3 ℃,平均气温19 ℃,区域平均年降水量1 454 mm,年日照时数超过1 765 h.土壤为山地黄壤.
午潮山林场有常绿阔叶林、竹阔混交林和毛竹纯林等森林类型.常绿阔叶林主要优势树种包括木荷(Schimasuperba)、苦槠(Castanopsissclerophylla)及青冈(Quercusglauca),其他树种有麻栎(Quercusacutissima)、女贞(Ligustrumlucidum)、桂花(Osmanthusfragrans)、丝栗栲(Castanopsisfargesii)等,主要灌木草本有檵木(Loropetalumchinense)、马银花(Rhododendronovatum)、映山红(Rhododendronsimsii)、山矾(Symplocossumuntia)、蕨类(Pteridophyta)、黄精(Polygonatumsibiricum)等.竹阔混交林主要优势树种包括木荷、苦槠、青冈及毛竹.毛竹纯林中乔木层为单一树种毛竹,而林下植物较为丰富.毛竹林下灌木主要以山橿(Linderareflexa)、蓬蘽(Rubushirsutus)、山胡椒(Linderaglauca)、六月雪(Serissajaponica)等为主,草本层主要是淡竹叶(Lophatherumgracile)、中日金星蕨(Parathelypterisnipponica)、大油芒(Spodiopogonsibiricus)等.
林场内试验区域不允许经营和采伐等活动,在常绿阔叶林受到保护的同时也使得毛竹存在明显的扩张现象,形成竹阔混交林或毛竹纯林.阔叶林主要为次生植被,林龄约有40年,毛竹林是毛竹入侵常绿阔叶林形成的毛竹纯林,混交林是毛竹向常绿阔叶林扩张后形成的混交林.
2 研究方法
2.1 样地调查
2018年3月,分别在毛竹林、竹阔混交林和阔叶林中各设立3个20 m×20 m的固定样地,共计9个样地.所有样地均选取于林分区分明显、有完整的毛竹入侵样带的同一区域中.各样地坡向为西南,坡度相近,平均海拔约为330 m.对所有样地进行群落调查,对样地内所有DBH≥5.0 cm的树木进行挂牌标记,并记录树种名.测定乔木胸径和树高,以及毛竹胸径,并记录毛竹龄级.
2.2 乔木层生物量估算
采集常绿阔叶林和竹阔混交林内优势树种的各器官样品若干,在65 ℃下烘至恒重,用于测量干重以及碳含量.乔木生物量采用模型法估算,将测得的胸径DBH及树高H数据输入模型进行生物量估算.其中木荷、青冈、苦槠等优势树种和麻栎等其他树种均选用相关系数(R2)较大的单木生物量模型[24-27].毛竹林则通过胸径和度数数据代入模型得出地上部分生物量[26],并根据根茎比推算出地下生物量[27],其总和即为毛竹总生物量.
毛竹生物量模型:
(1)
式中:f(D,A)为毛竹单株生物量(kg),A为单株毛竹年龄(度) ,D为单株毛竹胸径(cm).
苦槠生物量模型:
W=0.815 659 659(D2H)0.568 276 048
(2)
青冈生物量模型:
ln(W)=-1.989+2.554ln(D)
(3)
麻栎生物量模型:
W=0.018 851 976(D2H)1.091 681 772
(4)
木荷生物量模型:
ln(W)=-1.170+2.187ln(D)
(5)
式(2)、(3)、(4)、(5)中:W为乔木单株生物量,D为乔木胸径,H为乔木高度.
2.3 灌木层、草本层生物量
分别在相应固定样地外相邻区域随机设置5个2 m×2 m的小样方,采用收获法将样方内灌木草本包括根系全部收集称重.采集部分样品带回实验室,烘至恒重,用于含水率及碳含量的分析.
2.4 凋落物生物量
在样地内每100 m2设立1个1 m×1 m的凋落物采集器.采集器主体由100目防虫网构成,4角用PVC管进行固定.每季(1、4、7、11月)采集所有凋落物,在烘箱中65 ℃下烘至恒重,用于生物量及碳含量分析.
2.5 土壤样品采集
在每个固定样地内使用土钻随机选择3个点位钻取深度为0~20 cm、20~40 cm、40~60 cm的土壤,用环刀法测定土壤容重(ρb)及含砂率.将土样带回实验室风干、过筛,以备碳含量的测定.
2.6 碳储量计算
植物碳含量使用元素分析仪(CX-9800)进行测定,土壤碳含量用K2Cr2O7-H2SO4氧化法[28-30]测定.
各组分碳储量CS(carbon stock)=各组分生物量×相应碳含量
土壤有机碳储量SOCR(storage)=土壤容重ρb×采样深度depth×(1-含砂率)×土壤有机碳含量生态系统碳储量=乔木层碳储量+灌草层碳储量+凋落物碳储量+土壤层碳储量
2.7 数据分析与处理
数据分析在SPSS 21.0中进行,采用单因素方差分析比较不同林分碳含量、生物量、碳储量以及不同土壤碳储量的差异性,显著性水平设定为α=0.05.
3 结果与分析
3.1 林分生长及乔木碳含量
常绿阔叶林、竹阔混交林、毛竹林的乔木胸径及株数密度具有显著差异(表1),常绿阔叶林平均胸径为14.58 cm,显著小于混交林乔木的平均胸径18.86 cm(P<0.05);毛竹林中毛竹平均胸径略低于竹阔混交林中的毛竹平均胸径,二者无显著差异(P>0.05).常绿阔叶林乔木密度显著大于混交林(P<0.05),约为混交林的2倍;毛竹林的密度与竹阔混交林毛竹密度差异显著(P<0.05),分别为3 067株·hm-2和1 492株·hm-2.

表1 各样地林分株数密度及平均胸径1)Table 1 Density and average DBH of trees or bamboo in different sample plots
不同植物不同器官之间的碳含量不同(表2).木荷平均碳含量最高,为51.52%,其中干、枝、叶、皮、根分别为52.67%、51.64%、53.21%、48.89%、50.21%.青冈、苦槠等乔木碳含量介于44.65%~49.72%.样地中共有8株木质藤本,其平均碳含量约为46.24%.灌木草本的平均碳储量与木质藤本接近,为46.18%,上述植物碳含量差异均不显著,而凋落物中的平均碳含量相对偏低,仅为37.74%,与植物间存在显著差异(P<0.05).毛竹平均碳含量取50%[31].
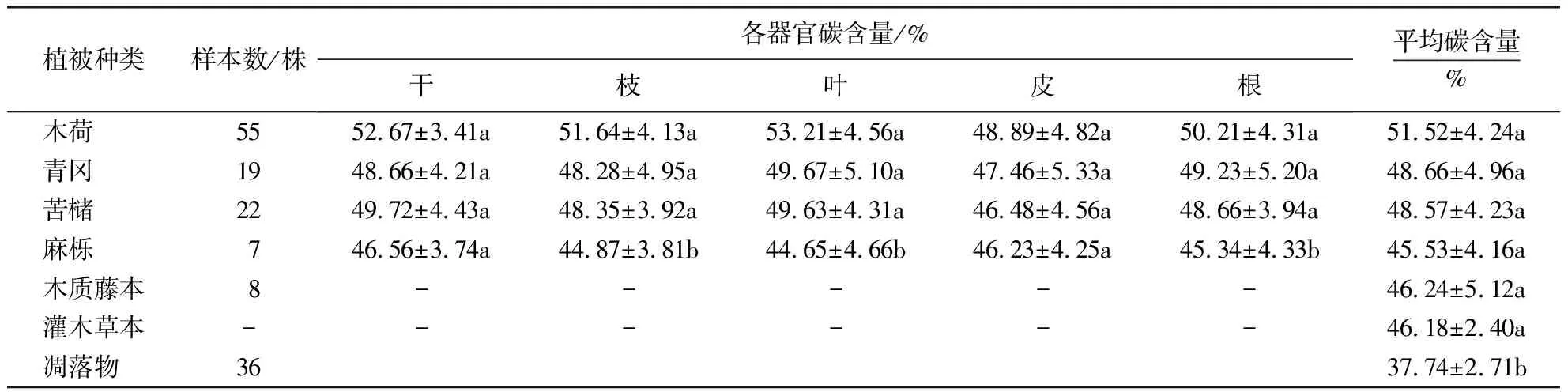
表2 乔木各器官与凋落物碳含量1)Table 2 Carbon content of different tree organs and litter
3.2 林分生物量及碳储量
毛竹林林分生物量及碳储量分别为68.61和33.97 t·hm-2,均显著低于常绿阔叶林的169.26和84.78 t·hm-2及竹阔混交林的185.71和88.64 t·hm-2,常绿阔叶林的生物量和碳储量低于竹阔混交林,但差异不显著(P>0.05).常绿阔叶林灌木草本层生物量及碳储量在3种林分中最高,生物量为5.30 t·hm-2,碳储量为2.45 t·hm-2.各林分间凋落物生物量差异不显著(P>0.05),在1.40~1.54 t·hm-2之间,碳储量0.53~0.58 t·hm-2(表3).
3.3 土壤碳储量
由表4可知,土壤容重在深度0~20 cm毛竹林最大,为0.90 g·cm-3,阔叶林最小,为0.81 g·cm-3,20~40 cm及40~60 cm深度毛竹林容重均为最大值,所有样地容重随着深度增加而变大.
土壤深度在0~20 cm时,含砂率介于4.22%~6.66%,在20~40 cm时介于8.72%~10.37%,40~60 cm时介于12.34%~14.84%.各个土壤深度中,毛竹林土壤碳含量均为最低,阔叶林土壤碳含量均为最高,除少量混交林碳含量有波动外,基本有着阔叶林碳含量>混交林碳含量>毛竹林碳含量的趋势.

表3 各样地林分生物量及碳储量分配格局1)Table 3 Distribution pattern of stand biomass and carbon storage in different plots
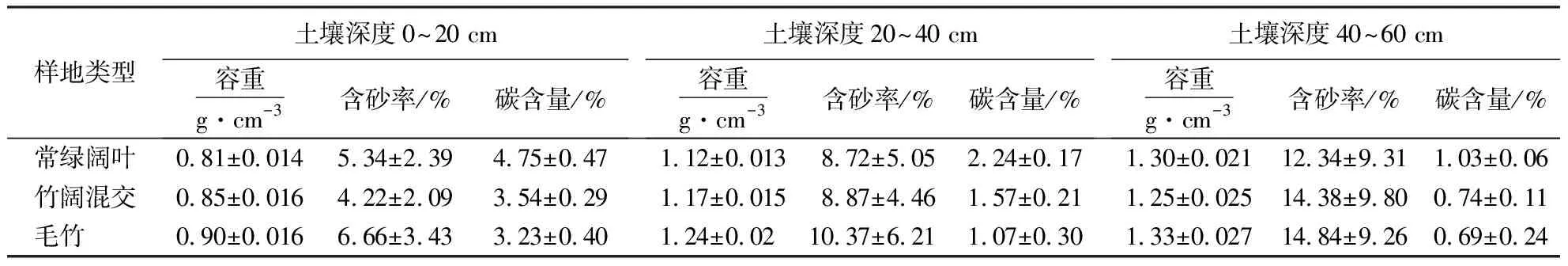
表4 各样地土壤碳含量分配格局1)Table 4 Distribution pattern of soil carbon content in different plots
由图1中所示,3种林分间土壤碳储量存在明显的差异.毛竹林土壤碳储量最低,为106.16 t·hm-2,常绿阔叶林最高为142.06 t·hm-2,竹阔混交林土壤碳储量略低于常绿阔叶林,为139.33 t·hm-2.3种林分土壤碳储量随土壤深度的变化规律相似,随土壤深度增加呈降低的趋势.同一土层深度3种林分间存在差异,其中土层深度0~20 cm的常绿阔叶林土壤碳储量(71.21 t·hm-2)>竹阔混交林(57.90 t·hm-2)>毛竹林(54.34 t·hm-2).在20~40 cm深度范围,竹阔混交林土壤碳储量(48.49 t·hm-2)>常绿阔叶林(45.87 t·hm-2)>毛竹林的33.51 t·hm-2.土壤深度在40~60 cm时,常绿阔叶林、混交林、毛竹林土壤碳储量依次为24.98、32.94、18.31 t·hm-2.
3.4 生态系统碳储量
常绿阔叶林、竹阔混交林、毛竹林生态系统碳储量按顺序分别为226.84、227.97、140.13 t·hm-2.毛竹林的总碳储量显著低于其他两种林分,其中植被碳储量差异最大;土壤碳储量毛竹林显著低于常绿阔叶林及竹阔混交林,常绿阔叶林与竹阔混交林间差异不显著.竹阔混交林生态系统碳储量最大,与阔叶林间差异不显著,但二者均显著高于毛竹林(图2).
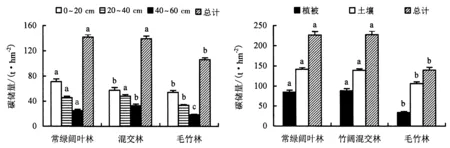
图1 3种林分土壤碳储量分布格局Fig.1 Distribution pattern of soil carbon storage in 3 types of stands
4 讨论
毛竹扩张过程致使生态系统碳储量发生变化,竹阔混交林生态系统碳储量略高于常绿阔叶林,但是毛竹纯林化后生态系统碳储量显著低于常绿阔叶林.这与杨清培等[23]和Fukushima et al[32]的研究结果基本相似,赵雨虹等[20]研究表明,混交林生态系统碳储量高于毛竹林,阔叶林生态系统碳储量最低.这可能与林分结构、入侵阶段等差异有关.本研究主要通过固定样地监测毛竹自然条件下入侵形成不同林分的生态系统碳储量.
原生态系统碳储量在毛竹入侵影响下发生改变,这可能与毛竹入侵会导致阔叶林群落组成和结构简单化、物种多样性下降[33,34]有关,竹阔混交林中乔木株数密度的降低与灌木草本层生物量的减少也说明了这一点.值得注意的是,竹阔混交林中乔木的平均胸径却均大于阔叶林,这与赵雨虹等[20]的研究结果相同.这可能是毛竹入侵初期主要对小径级乔木的影响较大,大径级乔木在混交林的留存率相对较高.这使得竹阔混交林阔叶树的碳储量并未减少太多,而毛竹通过其强大的根系形态可塑性[35]和快速积累生物量的能力,扩散到周边林分地带的毛竹胸径、株高均优于纯林[36].这些使得在一定时间内竹阔混交林的林分生物量和碳储量与原阔叶林相比较高.Fukushima et al[32]研究表明,毛竹入侵初级阶段混交林碳储量低于阔叶林,在入侵中期阶段混交林碳储量则高于阔叶林.本研究的混交林竹木株数比与Fukushima et al的研究相似,毛竹入侵后碳储量增加的结论与其相符.毛竹入侵虽然导致阔叶林物种多样性下降,但是入侵前中期淘汰的基本为小乔木以及灌木,碳储量高的大胸径阔叶树种在短期内受到的影响较小,而毛竹通过其快速生长积累碳储量[37],相应地弥补了小乔木以及灌木损失的碳储量,从而使得竹阔混交林在前中期阶段生态系统碳储量大于常绿阔叶林.
毛竹林的碳储量不及常绿阔叶林高,在林业生产中需科学合理经营毛竹林,以更好地保护好周边原生的常绿阔叶林.尽管试验结果估算的生态系统碳储量竹阔混交林最大,但是这并不意味着竹阔混交林优于常绿阔叶林.在竹阔混交林中,毛竹高于阔叶乔木,将阔叶林的树冠遮住,减少了阔叶树种对光的吸收[38],同时毛竹的化感作用抑制了阔叶林乔木树种的种子萌发和根系发育[39],导致竹阔混交林中木荷、苦槠等的幼苗数量少于常绿阔叶林.毛竹的这些特性可能会使竹阔混交林随着时间的推移对拥有较强固碳能力的阔叶树种产生排挤.因此对于毛竹扩张,还需要更长的时间追踪研究才能有合理的认知和相应的管控.竹阔混交林碳储量的波动也可以为毛竹林的扩张调控提供启发.
本研究中,竹阔混交林的竹木比例类型较为单一,还需今后进一步通过增加不同竹木比例混交林,研究不同入侵时期林分碳储量变化.同时,需要扩展研究区域,通过长时间多区域的观察测量,得出更准确的结果.当前毛竹竹材价格急剧下跌,加之廉价劳动力日渐缺乏,使得强集约型经济模式的竹林还将会不断被放弃经营,造成生产力低下,且弃管后加速毛竹对周边森林的入侵,可能对森林生态系统造成负面影响.因此,须对放弃经营的大片毛竹林采取营林手段进行改造,同时对扩张的毛竹林通过采笋伐竹等措施加以调控.
猜你喜欢
绿色天府(2022年9期)2022-11-19
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年2期)2022-05-07
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
