
锰胁迫对降香黄檀幼苗期生理特性的影响
2020-11-23 01:55杨红兰冯守富尹永昌敖福春黄秋婵
南方农业·下旬 2020年9期
关键词:丙二醛
杨红兰 冯守富 尹永昌 敖福春 黄秋婵

摘 要 为了探明锰胁迫下降香黄檀幼苗期的生理生化机制,以降香黄檀幼苗为实验材料,通过沙培法,研究了不同Mn2+浓度胁迫下降香黄檀生长和生理特性的变化。研究结果表明:随着Mn2+浓度的增加,降香黄檀植物体中的丙二醛(MDA)含量、过氧化物酶(POD)活性和活性糖含量均呈现出先升高后下降的趋势;在Mn2+浓度为4 mmol·L-1下,降香黄檀幼苗的POD值和MDA含量达到峰值;活性糖含量则在Mn2+浓度为20 mmol·L-1时出现峰值;同时,随着Mn2+浓度的不断增加,降香黄檀幼苗根、茎、叶积累锰的含量明显增多,根部是积累Mn2+的主要部位,其次是茎和叶。
关键词 降香黄檀;锰胁迫;POD酶;丙二醛
中图分类号:S792.28 文献标志码:B DOI:10.19415/j.cnki.1673-890x.2020.27.070
降香黄檀(Dalbergia odorifera T.Chen)又称黄花梨、花梨木,属蝶形花科(Papilionaceae)黄檀属(Dalbergia)植物,常绿半落叶乔木树种[1]。降香黄檀的药用价值很高,但其野生资源日益减少,再加上国内外市场对降香黄檀木材及其衍生品需求持续不断增加,导致其木材价格一直居高不下。巨大的市场潜力吸引了众多的社会资金投入造林,营建降香黄檀珍贵用材人工林。当前,国外对降香黄檀的研究非常少,基本上花精力在降香黄檀多样性、栽培技术以及活性物质提取等方面的研究上,针对锰胁迫对降香黄檀幼苗期的生理特性研究较少。目前,国内对重金属胁迫的研究相当广泛,包括农业、林业、经济作物、微生物、动物,对降香黄檀的研究有干旱胁迫、木材DNA的提取、混交林土壤细菌多样性等,但关于锰胁迫对降香黄檀的影响研究目前还相对缺乏。锰元素是植被生长发育所必须的元素,但环境中锰含量过高,也会对植物造成毒害[2]。研究锰胁迫下降香黄檀幼苗的形态和生理特性的变化以及各项指标的测定,可以更好地了解降香黄檀的生理特性,掌握在锰胁迫下降香黄檀对锰的耐性和其最适浓度,以了解降香黄檀对改良重金属污染土壤的可能性,也为降香黄檀的推广种植提供理论依据。
1 材料与方法
1.1 试验材料
采集当年成熟的降香黄檀种子进行幼苗培育。
1.2 试验方法
试验采用沙培法,当幼苗长出2片叶时进行移栽,选长势良好且相似的幼苗,用小勺子将沙培的幼苗轻慢挑出,尽量减少对根的伤害。移出后,每一个培养盆移植5棵,共移植30盆。自然光下培养,当降香黄檀幼苗长出5片叶时进行锰胁迫试验。使用药品为MnSO4,锰离子浓度梯度设置为0 mmol·L-1(对照)、0.8 mmol·L-1、4.0 mmol·L-1、20 mmol·L-1、100 mmol·L-1和200 mmol·L-1。胁迫时间为7 d,每次浇灌60 mL处理液,每隔24 h浇一次。胁迫期间,每天对降香黄檀幼苗的生长情况做好详细的记录。胁迫结束分别测定幼苗根、茎、叶的丙二醛(MDA)含量、过氧化物酶(POD)活性、可溶性糖含量,测定方法参照《植物生理学实验手册》[3]。用TAS-990F火焰原子吸收分光光度计测定吸光值,不同浓度下植物不同部位(根、茎、叶)的锰积累量根据公式1计算[4]。
式中:C为从标准曲线上查得元素的质量浓度,μg·mL-1;m为称量植物样品质量,g;V为植物样品试液的总体积,mL。
1.3 数据分析
以上实验每组重复做3次,取3次实验的平均值进行计算,再采用Excel 2013进行数据处理和制图。
2 结果与分析
2.1 不同Mn2+浓度对降香黄檀幼苗的影响
在对降香黄檀幼苗进行胁迫期间,前4天降香黄檀幼苗长势良好,无明显变化。第5天,高浓度(100 mmoL·L-1和200 mmol·L-1)Mn2+处理的降香黄檀幼苗叶子开始出现萎焉现象,且都有落叶(从嫩叶开始)。其中,100 mmol·L-1的Mn2+浓度处理降香黄檀幼苗个别植株出现黄叶(从老叶开始出现黄斑至整片叶子黄色),有些降香黄檀幼苗叶子还出现黑色斑点,叶面干燥缺水。到了第6天,这两个高浓度Mn2+处理的降香黄檀幼苗持续落叶,落叶非常严重,100 mmol·L-1的Mn2+濃度处理降香黄檀幼苗第一、第二片叶开始掉落。第7天,200 mmol·L-1的Mn2+浓度处理降香黄檀幼苗叶已经全部掉完。其他Mn2+浓度(0 mmol·L-1、0.8 mmol·L-1、4.0 mmol·L-1、20 mmol·L-1)处理的降香黄檀幼苗长势较好。
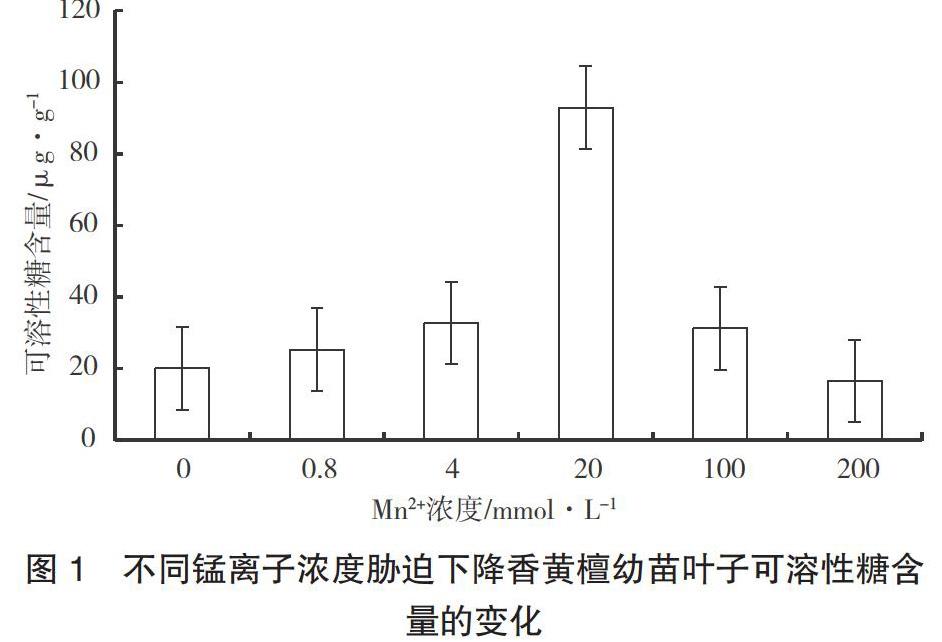
2.2 不同Mn2+浓度处理下降香黄檀幼苗叶子的可溶性糖含量变化
可溶性糖作为渗透调节物质之一,在植物适应盐分环境中发挥着重要作用,能够提高细胞的渗透调节能力,降低因渗透失水而造成的细胞膜、酶及蛋白质结构与功能损伤[5]。如图1所示,在不同Mn2+浓度处理下,降香黄檀幼苗的可溶性糖含量呈现出先升高后下降的趋势。在Mn2+浓度达到20 mmol·L-1时,降香黄檀幼苗的可溶性糖含量上升,达到最高值,为92.95 μg·g-1;在此之后,随着浓度的增加,可溶性糖含量有明显的下降。
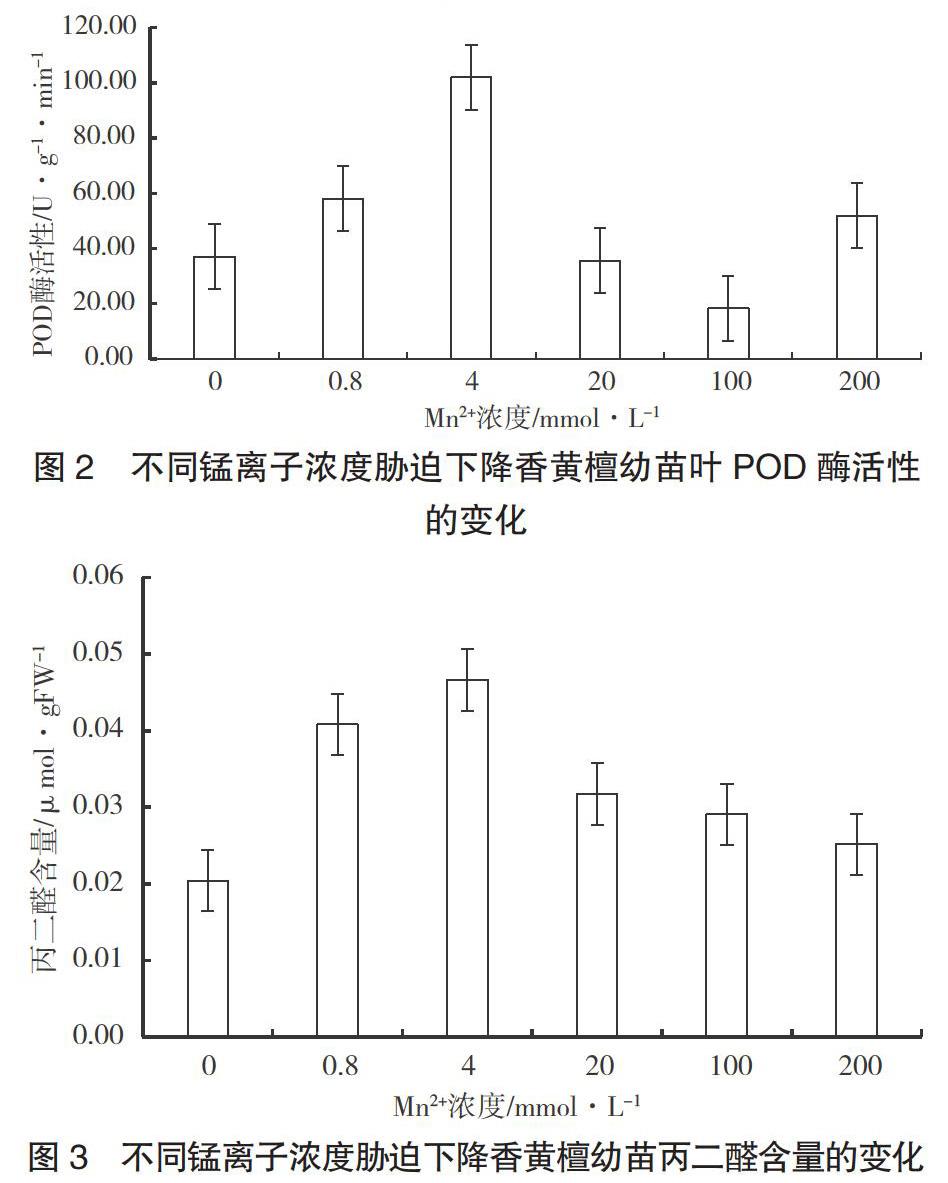
2.3 不同Mn2+浓度对降香黄檀幼苗叶片POD酶活性的影响
POD是抗氧化防御系统重要的抗氧化酶,其活性越高,植物对活性氧伤害的防御能力越强[6]。由图2可知,在较低Mn2+浓度下,降香黄檀幼苗叶的POD酶活性均明显高于对照组;处理浓度为0.8 mmol·L-1、4.0 mmol·L-1时,其POD值比对照组分别高20.80 U·g-1·min-1、64.83 U·g-1·min-1;而处理浓度为20 mmol·L-1、100 mmol·L-1时,POD值都比对照组低,分别低了1.52 U·g-1·min-1和18.69 U·g-1·min-1。Mn2+处理浓度为200 mmol·L-1时,降香黄檀幼苗叶的POD酶活性比对照组高,高了14.71 U·g-1·min-1。
从图中可以看到,当Mn2+浓度为4 mmol·L-1时,降香黄檀幼苗叶的POD酶活性最高。前段上升的原因,可能是锰胁迫对降香黄檀幼苗POD活性起到了诱导作用;后段下降原因可能是随着外源锰浓度的增大,降香黄檀幼苗超氧自由基的生成量较高,超出了POD酶清除超氧自由基的能力,导致植物体内自由基的清除与产生失衡[7]。
2.4 不同Mn2+浓度对降香黄檀幼苗MDA含量的影响
MDA是细胞膜脂过氧化反应的产物,其含量的变化可作为检测逆境条件下膜系统受损程度的指标[8]。从图3可以看出,在不同Mn2+浓度处理下,降香黄檀幼苗叶子的丙二醛含量相对于对照组均有不同程度增加,其趋势为先增加后降低。当Mn2+浓度为4 mmol·L-1时,丙二醛含量达到最高,随后有所降低。
2.5 Mn2+胁迫下降香黄檀幼苗不同部位锰的积累
从表1中可以看出,随着Mn2+处理浓度的增加,降香黄檀的根、茎、叶中的锰积累量也逐渐增加。而且从数据中可以看到,降香黄檀根部的Mn2+积累量明显高于茎和叶的Mn2+积累量,说明降香黄檀根部是主要吸收Mn2+的部位,且根部积累的Mn2+只有部分转移到了茎和叶,从而保护了地上部分。当Mn2+浓度为100 mmol·L-1时,降香黄檀幼苗表现出了受害症状。当Mn2+浓度为200 mmol·L-1时,降香黄檀幼苗已经萎焉,此时Mn2+的积累量到达最高,根部的Mn2+积累量为77.64 mg·kg-1,茎和叶的Mn2+积累量分别为77.22 mg·kg-1和39.66 mg·kg-1。
3 讨论
锰是自然界中广泛存在的元素,是维持植物正常生理活动的必需微量元素[9]。适量浓度的锰可以促进植物的生长,但环境中锰离子浓度过高就会对植物产生毒害作用。在本次研究中,降香黄檀幼苗在Mn2+浓度为0.8 mmol·L-1、4 mmol·L-1、20 mmol·L-1时长势良好,当Mn2+浓度为100 mmol·L-1时,降香黄檀幼苗开始出现叶子发黄、落叶现象。当Mn2+浓度为200 mmol·L-1时,降香黄檀幼苗开始萎焉,直至死亡。这说明在Mn2+浓度为100 mmol·L-1时,降香黄檀幼苗生长开始受到抑制,之后降香黄檀幼苗不能正常生长。因此,在种植降香黄檀时要注意土壤中Mn2+浓度。
在逆境条件下,植物器官通常会发生膜脂过氧化反应,MDA则是膜脂过氧化反应的产物,其含量的变化可表明植物细胞膜损害程度和植物对氧化胁迫的反应[10]。即MDA含量越多,植物受损害越严重;随着Mn2+浓度的增加,降香黄檀幼苗的MDA含量先升高后减低;在Mn2+浓度为4 mmol·L-1时,MDA含量达到最高值,之后开始下降。MDA下降的原因可能是高浓度金属胁迫处理导致植株幼苗代谢机能发生紊乱。高浓度锰离子使降香黄檀幼苗体内积累了大量的活性氧,导致植物细胞失去正常的代谢功能,从而不能积累MDA[11]。
POD酶是植物体内重要保护酶。植物体在Mn2+的胁迫下会形成过量的氧自由基,植物通过体内保护酶系统清除或减少逆境胁迫所产生的活性氧,从而避免其对组织细胞的伤害[12]。在本次试验中,降香黄檀幼苗叶子的POD酶活性随着Mn2+浓度的增加呈先升高后下降的趋势。POD酶活性在Mn2+浓度为4 mmol·L-1时达到最大值。在达到最大值之后,随着Mn2+浓度不断增加,POD酶活性开始下降。出现这样现象的原因是刚开始Mn2+浓度增加时,植物体内出现较多的过氧化物,诱使POD酶活性上升,分解这些有毒物质,提高植物体的抗逆能力。但随着Mn2+浓度的不断增加,植物体内的过氧化物数量超出了其能够分解的范围,植物开始受到毒害,进而导致了POD酶活性的下降。
可溶性糖作为植物体内主要渗透调节物质,在植物体调节细胞渗透压方面具有重要作用。本次试验结果表明,随着Mn2+浓度不断增加,降香黄檀幼苗体内的可溶性糖含量呈先升高后下降的趋势。在Mn2+浓度为20 mmol·L-1时,植物体内的可溶性糖含量达到了峰值,随后开始下降。植物体受到锰离子胁迫时,植物体内细胞渗透压失衡,植物体通过增加可溶性糖含量来调节细胞的渗透压,使其达到稳定状态。随着Mn2+浓度不断增加,可溶性糖含量下降的原因是高浓度锰胁迫超出了植物体所能承受的范围,植物体受到毒害,植物细胞的细胞膜通透性增大,糖类和酶大量运出[13]。
随着Mn2+浓度的不断增加,降香黄檀幼苗根、茎、叶积累锰的含量明显增多,富集量表现为根>茎>叶。在本次试验中,当Mn2+浓度达到最大值200 mmol·L-1时,降香黄檀幼苗根部的Mn2+积累量达到77.64 mg·kg-1,茎和叶的Mn2+积累量分别为77.22 mg·kg-1和39.66 mg·kg-1。虽然降香黄檀这种乔木植物不是超富集植物,但考虑到降香黄檀容易成活、耐旱、耐脊薄,因此可作为土壤植物修复的备选植物。
4 结论
1)锰是植物生长必需微量元素之一,适量的Mn2+浓度处理对降香黄檀幼苗的生长有一定的促进作用。但当Mn2+浓度达到20 mmol·L-1时,降香黄檀幼苗生长开始受阻,出现各种不良症状,直至死亡。
2)低浓度Mn2+对降香黄檀幼苗的活性糖含量、POD酶活性有一定的促进作用。高浓度Mn2+胁迫则对上述两个指标有抑制作用。其中MDA含量随着Mn2+浓度的增加,MDA含量呈先升高后降低的趋势。
3)降香黄檀对Mn2+的积累虽然没有达到超富集标准,但综合各方面来看,可以作为土壤植物修复的备选植物。
参考文献:
[1] 贾瑞丰,徐大平,杨曾奖,等.干旱胁迫对降香黄檀幼苗光合生理特性的影响[J].西北植物学报,2013,33(6):1197-1202.
[2] 尚伟伟,刘杰,张学洪,余轲.锰胁迫对青葙生长及叶绿素荧光特性的影响[J].生态环境学报,2013,22(8):1353-1357.
[3] 蔡庆生.植物生理学实验手册[M].北京:中国农业大学出版社,2013.
[4] 杨芬,张永伍,刘品华.植物中重金属含量的测定[J].科技创新与应用,2016(33):14-15.
[5] 杨宏伟,刘文瑜,沈宝云,等.NaCl胁迫对藜麦种子萌发和幼苗生理特性的影响[J].草业学报,2017,26(8):146-153.
[6] 邓勇,黄思齐,李建军,等.镉胁迫下红麻幼根的生理响应[J].中国麻业科学,2016,38(2):89-94.
[7] 张静,梁克中,陈秀红,等.铜胁迫对双穗雀稗生理特性及铜积累的影响[J].重庆三峡学院学报,2015,31(3):109-113.
[8] 文珂,郭晓玉,谭娜娜,等.锰胁迫对野大豆种子萌发及幼苗生理生化特征的影响[J].种子,2018,37(3):40-45.
[9] 曹恭,梁鸣早.锰—平衡栽培体系中植物必需的微量元素[J].土壤肥料,2004(1):49-50.
[10] Ramin L, Mohammad P, Puriya GK, et al. Physiolog-icalresponses of Brassica napus to fulvic acid under water stress:Chlorophyll a fluorescence and antioxidant enzyme activity[J].The Crop Journal,2015,3(5):434-439.
[11] 趙淑玲,王瀚,王让军,等.Cu2+胁迫对花椰菜种子的萌发及幼苗生理特性的影响[J].北方园艺,2017(5):16-20.
[12] 逄洪波,张雨欣,刘宁,等.镉胁迫对欧洲千里光幼苗生理生化指标的影响[J].沈阳农业大学学报,2015,46(4):492-496.
[13] 曾小飚,李毅,陈冠喜,等.麻风树幼苗对锰胁迫的生理响应研究[J].安徽农学通报,2015,21(22):22-24.
(责任编辑:赵中正)
猜你喜欢
广西植物(2022年5期)2022-06-18
中国医学创新(2016年34期)2017-02-28
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2015年11期)2016-01-27
天津农业科学(2015年12期)2015-12-03
中国当代医药(2015年18期)2015-08-06
江苏农业科学(2015年1期)2015-04-17
湖北农业科学(2014年9期)2014-08-08
食品与生活(2014年2期)2014-03-04
