
青藏高原不同种源麻花艽光合特征比较
2020-11-17 03:03:26周党卫师生波王环王亚艺王久利李松龄陈世龙沈建伟程庭峰李小娟
甘肃农业大学学报 2020年5期
周党卫,师生波,王环,王亚艺,王久利,李松龄,陈世龙,沈建伟,程庭峰,李小娟
(1.中国科学院高原生物适应与进化重点实验室,青海 西宁 810008;2.中国科学院西北高原生物研究所分析测试中心,青海 西宁 810008;3.青海省植物分子育种重点实验室,青海 西宁 810008;4.青海民族大学生态环境与资源学院,青海 西宁 810007);5.青海大学土壤肥料研究所,青海 西宁 810008)
青藏高原平均海拔4 000 m,被誉为世界第三极.独特的自然地理环境,孕育了许多特有的植物资源.这些植物长期适应高原低气压、强辐射、低温等逆境环境,在生理生化和形态结构等方面具有独特的适应特征[1-3],已引起了国内外学者的普遍关注[4-7].早在1991年,就有高山植物的移栽驯化试验,并通过与原产地比较,揭示了叶片CO2同化速率和水蒸气交换速率的适应性具有异质性[8].夏明忠[9]对分布于不同海拔梯度的高原粳稻进行了研究,发现其光合适应特性存在明显适应差异.师生波等[10]对青藏高原的3种人工栽培植物的研究也表明其光合特性差异较大,且与低海拔植物无相似性.近期,赵玉文等[11]对西藏砂生槐的引种研究也发现不同种源材料的光合特征存在差异.但目前关于不同种源高山植物移栽驯化后光合特性的研究还十分有限.
叶绿素荧光测定是一种快速、无损伤的检测植物光合作用运转的技术,叶绿素荧光动力学分析在监测叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用[12].由于最大光量子效率(Fv′/Fm′),实际光量子效率(ΦPSⅡ),光适应光化学猝灭系数(qP),非光化学猝灭系数(NPQ)等荧光参数对植物非生物胁迫具有较好的指示性,因此,在植物表型鉴定和农作物品质选育方面已广为使用[13];并且在木本植物、农作物和园林植物筛选方面得到应用[12-15],对植物的合理栽培与生理适应了解也具有重要作用[15-16].但该技术在高山植物种质筛选方面还鲜有报道.
麻花艽(GentianastramineaMaxim.)是广布于青藏高原的高寒草甸代表性伴生种,为龙胆科(Gentianaceae)龙胆属(GentianaL.)植物,分布于海拔2 500~4 700 m的山坡、河滩、草地和灌丛[17-18].麻花艽为一种重要的中藏药材[19-20],在高原地区人为采挖野生麻花艽现象较为严重,不仅严重破坏了高寒草甸的生态环境[21],也造成麻花艽种质资源的损失.尽管,目前麻花艽的人工栽培已经取得一定进展[22-23],但关于优良种质研究仍极为缺乏,对异地移植后其生理适应性也缺乏了解.已有研究表明,麻花艽光合速率和叶绿素荧光参数与所在地的海拔有较大关系[10,21].但异地移栽后这种现象与原产地海拔间的关系是否依然存在,其光合特性是否会呈现趋同适应?对此尚缺乏必要的探究.鉴于此,本研究对不同海拔种源的麻花艽进行了1 a的人工驯化栽培,并分析了光合色素、光合气体代谢参数和叶绿素荧光参数的变化,以期揭示麻花艽的光合特征差异,为麻花艽的优质种质筛选,遗传多样性保护和引种栽培提供科学依据.
1 材料与方法
1.1 试验样地与材料
试验样地位于青海省湟中县多巴镇实验苗圃,距西宁市研究所约25 km;N 36°39′,E 101°31′,海拔2 360 m.该区域为湟水河山前河谷耕地(蔬菜和春油菜种植区),土壤为灰钙土,有机质为11.26 g/kg,全氮为1.04 g/kg,全磷为2.31 g/kg,全钾为30.03 g/kg,碱解氮为73.00 g/kg,速效磷为9.70 g/kg,速效钾为235 g/kg,pH 8.32;年均温2.6~5.3 ℃,平均降水量378.3 mm,且多集中于6~8月份.年均日照时数2 588.3 h,日照百分率达59%.2017年植物生长季的7~8月份,在青藏高原的不同地点(表1)选择生长健壮的野生麻花艽植株各6~10株,带土移回并栽培于验苗圃内;栽培前土壤平整后每667 m2使用10 kg尿素和5 kg磷酸二铵作为底肥,定植株间距为20 cm.经越冬适应后,在生长季6月初,人工祛除杂草后,架设遮阳网,避免中午过强光照;生长期给予适时浇水.在2018年植物开花期的8月份选取生长健壮、大小接近的植株进行试验.

表1 不同种源移栽麻花艽采样点信息
1.2 试验方法
1.2.1 叶绿素和类胡萝卜素含量测定 参照朱广廉等[24]的方法,在完成光合气体代谢参数和叶绿素荧光参数测定后,取成熟叶片,每一类群植物用打孔器取0.7 cm叶圆片30片,分3组,分别浸泡于丙酮乙醇混合水溶液中(乙醇∶丙酮∶水=45%∶45%∶10%).密封4 ℃暗浸提10 d后用岛津UV-1602测定663、652、645、470 nm等处的光吸收值,按照Arnon[25]的方法计算叶绿素各组分和类胡萝卜素的含量.
1.2.2 光合气体代谢参数测定 选择天气较为稳定的全晴天,于苗圃选取健康植株,采用Yaxin-1102型光合蒸腾测定仪(北京雅欣公司),依次测定不同种源麻花艽叶片的净光合速率(Pn)和气孔导度(Gs)等气体代谢参数;测定气源来自样地5 m开外约2 m高的大气.测定时间选太阳日照强度及大气温湿度较为稳定的9∶00~11∶00[10],气温为(27±1)℃,相对湿度(42±2)%, CO2体积浓度(385±4)μmol/mol,测定空气流速控制在0.1 L/min.不同驯化类群,选择大小一致的3个独立株进行测定,测定时选择成熟无损伤叶片中部进行测定,测定后,用尺子测定选择测定部分的叶面积,对测定结果进行校正.
1.2.3 叶绿素荧光参数测定 叶绿素荧光参数采用英国产便携式脉冲调制荧光仪FMS-2(Hansatech Instruments LTD.,Norfolk,UK)测定.选取健康植株上长势一致的麻花艽3片叶片,经暗适应30 min后测定初始荧光(Fo),最大荧光(Fm),计算最大光化学效率Fv/Fm;开启荧光仪器内置光源,测定860 μmol/(m2·s)稳态光强下的叶绿素荧光参数(Fs),光化学作用光下的最大荧光强度(Fm′);光化学猝灭系数qP= (Fm′ -Fs)/(Fm′ -Fo′),非光化学猝灭系数NPQ=(Fm-Fm′)/Fm′,PSⅡ反应中心实际光化学效率ΦPSⅡ= (Fm′ -Fs)/Fm′.快速光响应曲线(RLC)测定采用内置光源获取,对自然光诱导叶片1~1.5 h后,进行连续光化学梯度(0、21、57.5、125、232、380、580、820、1 120、1 480、1 880)μmol/(m2·s),仪器发射饱和脉冲,直到1 880 μmol/(m2·s)结束,获取不同光强下的叶绿素荧光动力学参数,每一光强下的平衡时间为10 s.光合电子传递速率ETR=ΦPSⅡ×PAR×0.84×0.5[26],相对电子传递速率的初始斜率α、ETRm和EK值按照Eilers and Peeters的公式[27]进行光响应曲线拟合,参数数值按照下列公式进行计算:
ETR=PAR×(a×PAR2+b×PAR+c)-1
式中,α=1/c;ETRm=(b+2×(ac)1/2)-1;EK=c×(b+2×(ac)1/2)-1
1.3 数据统计分析
2 结果与分析
2.1 不同种源麻花艽的光合色素比较
叶绿素a(Chl a)和叶绿素b (Chl b)是植物捕获光能进行光合作用的主要组分.如表2所示,Chl a含量在不同类群麻花艽有较大差异,其中碌曲最高,河南最低,二者相差41.376 mg/g,差异极为显著(P<0.05);达日和碌曲的Chl a含量尽管略高于互助类群,但却并无显著差异.Chl b含量仍为碌曲类群最高,互助最低,其中碌曲和达日类群显著高于互助类群,Chl b含量随海拔有增高趋势(P<0.05)(表2).类胡萝卜素(Car)含量在不同种源麻花艽中也存在差异,其中达日和碌曲类群显著高于互助,达日和碌曲驯化类群总体Car含量较高.碌曲类群的总叶绿素(Chl)含量显著高于其他种源的麻花艽(P<0.05),尽管河南类群Chl含量与互助和同仁类群无明显差异,但仍显著低于碌曲和达日类群,说明Chl含量在高海拔驯化类群中呈现增高趋势.Chl a与Chl b的比值(Chl a/b),互助类群最大,显著高于其他类群(P<0.05),河南类群值最小,其比值在高海拔驯化类群中有降低趋势;而Car与Chl比值在各驯化类群中则随原产地海拔增高呈增高趋势.
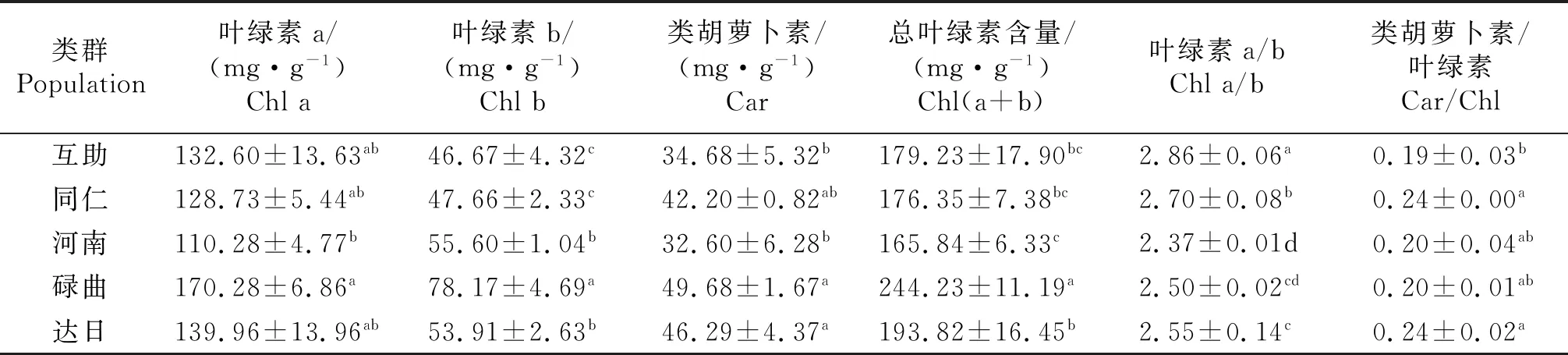
表2 不同种源麻花艽叶绿素与胡萝卜素含量的比较
2.2 不同种源麻花艽净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)及胞间CO2浓度(Ci)比较
光合速率Pn在不同种源麻花艽驯化类群中差异并不显著(P>0.05).由图1-A可以看出,碌曲类群的Pn较高,平均光合速率达到18.11 μmol/(m2·s),其次为达日类群,Pn最小的类群为互助类群.海拔最高的达日类群Pn较互助高38%,是海拔最低互助类群的1.21倍.进一步分析光合速率Pn与起源地海拔的相关性表明,其相关系数为0.782.说明麻花艽移栽驯化类群的Pn随海拔升高呈现出增高的变化趋势.

图1 不同种源麻花艽的光合特征比较Figure 1 Comparison of photosynthetic characteristics among G.straminea from different sites
不同类群麻花艽的蒸腾速率Tr分析表明,互助类群最高,而同仁类群最低;但与其余3个驯化类群的Tr无明显差异(P>0.05);互助类群的Tr值显著高于同仁、河南、碌曲和达日类群(P<0.05)(图1-B).不同种源麻花艽气孔导度Gs显示同仁类群的最大,河南类群最小且显著低于其他种源地麻花艽类群(P<0.05),碌曲和达日驯化类群的Gs略有降低趋势(图1-C).叶片细胞间CO2浓度Ci则在不同种源间存在明显差异(图1-D),其中互助和同仁麻花艽类群叶片的Ci分别为103.14 μmol/mol和123.30 μmol/mol,高于河南、碌曲和达日驯化类群,且碌曲类群Ci最低.在不同移栽类群中,同仁类群Ci值显著高于河南、碌曲和达日3个麻花艽驯化类群(P<0.05),但与互助类群的差异并不显著(P>0.05),Ci在驯化类拔类群随原产地海拔呈降低趋势.
2.3 不同种源麻花艽PSⅡ有效光化学效率与实际光化学效率的比较
PSⅡ反应中心有效光化学效率(Fv′(/Fm′)是反映PSⅡ原初光反应中心光能捕获效率的重要指标.自然环境下,麻花艽主要分布在水分较好的滩地及山坡半阴面,高光强易引起光抑制,因此设稳态作用光强为860 μmol/(m2·s).如图2-A所示,在860 μmol/(m2·s)光强下,尽管统计分析显示不同类群间Fv′/Fm′无明显差异(P>0.05),各个种源间同仁麻花艽类群的Fv′/Fm′为最高,其次为互助类群,河南类群最低;而河南、碌曲和达日类群Fv′/Fm′略低于低海拔的互助和同仁类群.同样,不同种源麻花艽的实际光化学效率ΦPSⅡ也无显著差异(P>0.05),其中低海拔的互助和同仁类群ΦPSⅡ差异不大,略高于其他3个类群(图2-B).说明,PSⅡ反应中心的Fv′/Fm′和ΦPSⅡ在高海拔引种驯化类群中较低,其中ΦPSⅡ在不同驯化类群中与起源地海拔类群中呈现负相关性,相关系数为-0.837(P=0.077).
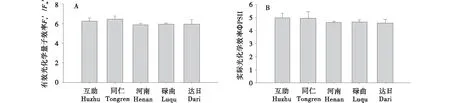
图2 不同种源麻花艽叶片实际光化学效率和有效光化学效率的比较Figure 2 Comparison of the PSⅡmaximal and actual photochemical efficiency among the leaves of G.straminea from different sites
2.4 不同种源麻花艽光适应下光化学猝灭和非光化学猝灭比较
光化学猝灭系数qP是反映光反应中质体醌氧化状态的重要参数,其数值大小与反应中心光合电子传递的效率相关,是光合作用PSⅡ原初反应中心光能利用的重要指标.稳态860 μmol/(m2·s)光强下,不同种源的比较结果显示,qP在不同驯化类群麻花艽叶片之间差异极小(图3-A),而非光化学猝灭系数NPQ在不同驯化类群间存在一定差异(图3-B),其中达日类群NPQ最高,同仁类群的最低,较达日降低31%,二者差异显著(P<0.05).

图3 不同种源麻花艽叶片光化学及非光化学猝灭系数的比较Figure 3 Comparison of photochemical and non-photochemical quenching coefficient in the leaves of G.straminea from different sites
图4为光化学和非光化学猝灭系数的光响应特性.表明,qP值随着光合有效辐射PAR增高,呈现出几乎直线降低的趋势;在较低光强范围,除互助类群外其他几个驯化适应类群之间均差异较小,随PAR增加,高海拔的达日类群,其qP明显高于低海拔互助类群(图4-A).NPQ的变化随PAR的增加呈现增加的态势(图4-B),在高光强下,达日驯化类群明显较高,而河南与达日类群的NPQ接近.说明,不同种源麻花艽qP和NPQ随光强的变化趋势差异明显.并且即使在1 850 μmol/(m2·s)时,不同驯化类群NPQ仍呈增加趋势,呈现对强光的较强适应性.其中当光强高于1 200 μmol/(m2·s),达日类群的qP和NPQ值均高于其他驯化类群,其强光适应性呈现出互助<同仁<碌曲<河南<达日.

图4 不同种源麻花艽类群叶片光化学猝灭系数与非光化学猝灭系数的变化Figure 4 Variation of PSⅡphotochemical and non-photochemical quenching coefficient in the leaves of G.straminea from different sites
2.6 不同海拔麻花艽类群表观电子传递效率ETR的光响应比较
由图5-A可以看出,相对表观电子传递速率ETR随着光合有效辐射PAR的增加呈增强趋势,大于1 000 μmol/(m2·s)后逐渐趋于平稳.尽管高光强时不同种源麻花艽类群的ETR均较稳定,但达日类群的ETR值明显较高,而河南、碌曲和同仁类群的ETR值差异较小,且仍明显高于互助类群.其大小顺序为:达日>河南>碌曲>同仁>互助.说明高海拔麻花艽驯化类群的表观光合电子传递效率较高.
在中低光强范围内(0~600 μmol/(m2·s)),不同类群麻花艽叶片表观电子传递速率ETR与光合有效辐射PAR增加几乎为线性(图5-B),直线回归的拟合相关系数在达日、碌曲、河南、同仁、互助分别为0.96、0.95、0.98、0.97和0.99(P<0.01),呈现随海拔升高而升高的趋势.光响应曲线的初始斜率α值在同仁、河南与碌曲类群斜率较为接近(0.237 5、0.230 5和0.238 0),明显高于低海拔的互助类群0.159 7,而低于达日驯化类群(0.257 6).说明,麻花艽类群在较低光强范围ETR与PAR之间呈现极显著直线相关关系,高海拔类群具有更高效的光能利用效率.达日、碌曲、河南、同仁、互助不同类群麻花艽的最大电子传递速率ETRm分别为103.44、140.53、155.44、147.28和195.90 μmol/(m2·s).而不同类群间最小的饱和光强度EK值[23]也存在差异,其中互助和同仁类群的EK值分别为467 μmol/(m2·s)和406 μmol/(m2·s),而河南、碌曲和达日驯化类群EK值分别为565、510和658 μmol/(m2·s).说明,起源于高海拔达日.河南和碌曲驯化适应类群的ETRm和EK值都高于低海互助拔类群.

图5 不同种源麻花艽类群叶片光合电子传递ETR的光响应Figure 5 Comparison of light response of electron transfer reaction in the leaves of G.straminea from different sites
2.7 不同种源麻花艽引种驯化后,海拔与光合特征参数的相关性分析
为了解移栽后,原产地海拔对光合特征参数的影响及个光合参数的关系,我们对不同种源地麻花艽的光合相关参数进行了分析.如表3所示,引种地海拔与叶绿素a、叶绿素b、总叶绿素含量、光合速率均呈正相关关系,而与胞间CO2浓度,蒸腾速率和气孔导度呈负相关关系.其中与光合速率的正相关性接近达到显著水平.同样,叶绿素a和总叶绿素含量与Pn呈正相关关系.这与水稻中的研究结果一致[28].同样,在光合荧光参数中,非光化学猝灭系数NPQ与麻花艽种源地海拔具有正相关性,相关系数为0.461.而最大电子传递速率ETRm与种源地海拔则呈现出极显著的相关关系,相关系数为0.943(P<0.05).因此,根据叶绿素、光合速率特征,结合引种驯化荧光特征,可以看出,不同种源麻花艽中,达日、碌曲和河南类群,特别是达日类群,具有优良的适应性和光合潜力,在引种栽培上具有很好的利用价值.
3 讨论
麻花艽是青藏高原的特有物种[17],也是传统中藏药材“秦艽”的代表种[19-20].因其资源储量大,品质好而在秦艽市场上占有相当的比例[18].目前,麻花艽的市场供应仍主要依赖于野生资源.由于长期的野外采挖,不仅破坏高原脆弱的生态系统,也易于造成物种的濒危和物种多样性的损失[18,29].尽管,目前麻花艽的人工栽培已经取得一定进展[22-23],但优良种质筛选研究仍极为缺乏,对异地移植后其生理生态适应性更缺乏了解.因此,研究移栽麻花艽的驯化适应性,对于高山药用植物栽培、种质选育及多样性保护等均具有重要意义.

表3 麻花艽原产地海拔与光合特征参数相关分析
已有研究表明,高山植物在极端环境条件下,其形态解剖和生理生化特征都发生明显的改变[1-3].作为植物生存最重要的储能和形成有机物质的光合作用,其光合器官也对高山胁迫环境存在明显的响应和适应[4-7].有研究结果表明,光合作用总叶绿素含量随着海拔的升高而降低,而类胡萝卜素含量随海拔而升高[3].本研究发现异地移栽后,总叶绿素含量和类胡萝卜素含量在不同种源麻花艽类群间依然有差异,高海拔种源达日与碌曲驯化类群均具有较高总叶绿素含量和类胡萝卜素含量,其中叶绿素b的变化趋势更为明显.有研究认为,光强和光质都能对叶绿素合成产生影响[30],而温度对叶绿素合成也具有重要调节作用,低温降低叶绿素的含量[3].高海拔地区的类群,其叶绿素和色素合成在原产地可能受到低温条件的限制,因此,合成效率可能较低[4];而植物生长季8月份栽培区域内水热同期,使得叶绿素和胡萝卜素合成在高海拔类群中更为活跃.叶绿素a和b的比值尽管在不同类群中存在差异,但整体具有一定程度的降低,这与野外的测定结果有所不同[3-4].对荒漠环境下的云杉研究也发现,强光下叶绿素含量的降低可能是一种驯化适应[7],因此,叶绿素在不同类群的差异,也可能是对移栽地区环境的驯化适应.而类胡萝卜素含量的增加,则与其对高海拔强光的适应有可能关[3].高海拔来源的植物长期受到强太阳辐射,特别是UV-B辐射的影响,类胡萝卜素不仅可以在光反应中传递电子,而且作为非抗氧化物质来清除过多的自由基,因此高含量的胡萝卜素含量和Car/Chl能更有利于植物光合器官的保护[31].因此,异地移栽后,麻花艽类群的叶绿素合成的改变可能存在着一定的驯化适应,但高海拔植物具有高含量胡萝卜素可能是因为更多受到原产地环境的影响.
夏明忠[9]比较了不同来源水稻的光合特征,认为高光强下高海拔水稻具有较高的光合速率.Westbeek等[32]比较了高山、亚高山和低海拔起源的7种植物,发现高海拔地区的植物光合速率明显大于低海拔地区而羧化能力也一样,这与本研究的结果一致.本研究发现,异地移栽后,麻花艽类群的光合速率与引种地海拔具有显著正相关性(P<0.05),而胞间CO2浓度则呈现降低的趋势,说明引种移栽后,高海拔来源的麻花艽仍然具有较高的CO2同化能力.Rubsico酶在光合作用中具有重要作用,其遗传和动力学特征可能已经在长期的低温、低气压等高山环境下发生了动力学性质改变,羧化能力提高[3].但也有研究表明,高海拔植物的光合速率并不高,可能是低气压导致的结果[33].应该看到,麻花艽类群的光合速率尽管随海拔有所增高,但类群间并没有达到显著水平,这可能与综合因素限制有关[4].同样,有研究认为,随着海拔升高,气孔导度增加[3];但本研究结果显示,不同种源的麻花艽类群,气孔导度随着海拔的升高具有降低的趋势.气孔导度受光质和温度影响,高温降低气孔导度[34].高海拔植物的光合最适温度降低[3].因此,异地移栽后,不同种源麻花艽类群的气孔导度降低,可能也与栽培地区夏季的温度较高有关.高海拔类群气孔导度的降低,导致胞间CO2降低,构成光合速率的限制因素,使得高海拔类群净光合速率并未显著高于低海拔的互助和同仁类群,但其趋势仍然表现出随海拔增高的趋势.这也与前期矮嵩草、高山稻的研究结果一致[4,7],说明起源地可能仍然是影响麻花艽光合潜力的主要因素.
叶绿素荧光技术是分析光合机构PSⅡ光化学活性的有力工具[11].Fv′/Fm′反映PSⅡ原初光能捕获效率,而PSⅡ非光化学猝灭系数NPQ则反映光合机构通过非辐射能量耗散过剩激发能的能力,调节PSⅡ光合量子效率[33].本研究发现,不同类群麻花艽的Fv′/Fm′在稳态光强860 μmol/(m2·s)下均无显著差异,而NPQ随海拔升高有增强趋势.师生波等[7]对云杉的研究发现,高光强下,其NPQ在移栽地区无差异,存在趋同适应性.但不同种源麻花艽NPQ值的差异可能更多与其起源地有关,其中高海拔达日、碌曲和河南类群的NPQ值均高于低海拔同仁和互助类群,是否存在趋同适应尚待进一步观察.在稳态光强下,麻花艽NPQ平均值约为1.5~2.5,均低于3.0,呈现了一定的耐阴性[3],这也与麻花艽类群在高原的分布生境一致.
叶绿素荧光参数曲线可反映植物光合功能对环境变化的快速响应与应答.本研究结果显示,光化学猝灭系数qP的光响应曲线随光强呈现降低的趋势.qP反映了电子传递过程中质体醌的QA的氧化还原状态,光环境中QA接受电子,形成还原态NADH,可将同化力转化为有机物的化学能,qP的降低意味着强光对PSⅡ反应中心产生了抑制作用.高海拔类群的qP降低程度明显低于低海拔类群的,说明高海拔起源的麻花艽类群发生光抑制的程度弱于低海拔的,能更好的适应高原的强太阳辐射.这可能与非光化学猝灭系数NPQ增强有关.不同海拔麻花艽非光化学猝灭系数NPQ随光强的增加其变化趋势较为一致,高海拔类群升高较明显.强光下NPQ较高,表明此时植物类囊体膜上的PSⅡ反应中心多呈失活状态,失活的反应中心与光捕获天线色素一起共同参与过剩激发能的耗散过程,可保护功能反应中心免遭多余光能的伤害[35-36];同样,高海拔类群具有较高类胡萝卜素含量,能有助于抵御强光产生的过量自由基对于反应中心的伤害.随着海拔升高,太阳辐射增强,原产地高海拔的麻花艽类群可通过非光化学能量耗散的增强保护PSⅡ反应中心,是对高海拔强光辐射的一种适应.
ETR描述PSⅡ反应中心相对的电子传递速率.一般认为,低于200 μmol/(m2·s)光强下,光合电子传递与光强之间呈直线相关关系[34];而本研究发现,光强小于600 μmol/(m2·s)的中低光下,麻花艽的ETR与光强之间也呈显著正相关关系.其中高海拔的α值显著大于低海拔的互助和同仁类群,表明高海拔麻花艽类群具有更高的光反应效率.而光强大于600 μmol/(m2·s)后ETR增加缓慢且在高光强下略有降低的趋势,这可能与反应中心发生了光抑制,导致ETR降低有关[37].最小饱和光强EK值表明,不同种源麻花艽的数值在410~658 μmol/(m2·s),并且达日、碌曲和河南类群具有更高的最小光饱和值,说明其光抑制的发生需要更高的光照强度.ETR随光强的变化趋势一致,且来源于高海拔的麻花艽类群具有较高的ETR数值,同样高海拔类群的ETRm显著高于低海拔的互助和同仁类群.这表明,高海拔来源的麻花艽其光反应中心的电子传递效率也显著高于低海拔种源地区.同样,光响应曲线也显示,高海拔类群具有更高的Fv′/Fm′和NPQ值,并且无论在适宜光照强度还是高光强下,其ETR都随海拔而明显增强.
4 结论
总上所述,异地移栽驯化过程中,不同海拔麻花艽类群的光合气体代谢参数以及叶绿素荧光分析数据均显示,叶绿素含量、光合速率、非光化学猝灭系数和最大电子传递速率均与海拔有正相关性.移栽驯化1 a后,原产地海拔依然对麻花艽光合特性有重要影响,这也与前人的光合特征研究结果一致[8-9].因此,高海拔的达日、河南和碌曲驯化类群具有更高光合潜力和对强光适应性.近期研究也表明,NPQ在植物应对强光环境的协同进化过程中起着重要作用[38].高原地形复杂,生境千差万别,强光、低气压、低温等胁迫因素可能不仅塑造着植物的外形,而且对光合代谢过程也产生有深刻影响[3].高海拔植物在生长过程中遭受更高的太阳辐射和紫外辐射[3,6,19],在合成光合产物的同时,必须耗散掉过剩的激发能,保护光反应中心.因此,高海拔起源的麻花艽类群可能通过较高的光化学反应和非光化学能量耗散,以及高的电子传递速率来适应强太阳光辐射,维持光合机构的有效运行;此外,较高含量的胡萝卜素和叶绿素b含量也是高海拔麻花艽类群保护光合器官的有效策略.
猜你喜欢
作文小学高年级(2022年6期)2022-07-01 09:41:30
云南化工(2021年9期)2021-12-21 07:44:10
阅读与作文(小学高年级版)(2021年8期)2021-09-12 16:55:18
World Journal of Integrated Traditional and Western Medicine(2020年12期)2020-04-20 03:01:36
读友·少年文学(清雅版)(2018年1期)2018-09-11 01:40:34
人大建设(2018年3期)2018-06-06 03:11:01
人大建设(2017年10期)2018-01-23 03:10:15
阅读与作文(小学高年级版)(2017年7期)2017-08-04 08:51:09
人大建设(2017年12期)2017-03-20 05:22:25
影像科学与光化学(2014年5期)2014-03-11 16:03:23
