
一种利用组织培养水平转移遗传物质的新方法应用
2020-11-09 09:26
现代园艺 2020年20期
(长治职业技术学院,山西 长治 046000)
传统的嫁接是一种营养繁殖方式。前人已经证明嫁接可以导致砧木和接穗之间的水平基因转移[1]。然而,尚未报道通过嫁接在作物育种中进行水平基因转移的应用。通过组织培养方法(GTCM)创建了一种新的类似移植方法,以促进不同材料之间细胞质遗传物质的水平交换。
胞间连丝是一种细胞间通道,在细胞间可以交换各种分子,包括核酸、蛋白质、离子、水甚至病毒[2]。作物杂种优势利用的主要方法是CMS 系统,遗传雄性不育系统(GMS),自交不亲和系统,雄性不育系的基因工程,化学杂交剂等。CMS 系统是不育系的细胞质和核相互作用的结果。在这种类型的CMS 中,想要实现保持系,传统的方法是人工合成保持系。整个过程需要杂交,测试和自交多代,大面积种植[3]。
由于砧木和接穗之间的遗传物质的交换不会在远距离的情况下发生,通过组织培养法(GTCM)创造了一种新的类似嫁接法,可广泛应用于不同种质之间的细胞质遗传物质的交换。即使在不同的物种之间也可以使用这种改进的方法。步骤如下,首先,在无菌环境中将2 种类型的嫩茎切成2 种不同的大小。其次,来自2种类型种质的愈伤组织接合在一起生长并形成整体,其中一些细胞质遗传物质相互交换和转移。第三,相互作用的愈伤组织在不同的培养基中分化,再生和生长芽和根。最后,分子鉴定和表型确认进一步验证了阳性细胞系。
1 材料和方法
物料A:供体材料,细胞质雄性不育系Ogu89,半冬甘蓝型油菜不育系;B:受体材料,隐性正常细胞质育性系,中双11 号是2008 年在我国发布的优良半冬甘蓝型油菜品种;通过从Ogu89 到中双11 的细胞质遗传物质的水平转移开发隐性CMS 保持系。
通过组织培养方法在不同材料之间水平转移遗传物质[4]。详细过程(7 个步骤)描述如下:
1.1 材料准备
将来自A 供体材料(Ogu89)和B 受体材料(中双11)的种子浸泡约24h,挑出腐败种子浸入95%乙醇中1min,用无菌水清洗种子2~3 次,在1%氯化汞中连续洗涤15min,再用无菌水洗涤3~4 次。最后,在MS 培养基上播种。
MS 培养基配方:4.4g/LMS 粉末+20g/L 蔗糖+2.6g/L植物凝胶。
1.2 采用组织培养法和外植体去分化材料相似接枝
当幼苗的下胚轴伸长约10cm 时,将材料A(Ogu89)切成约5mm 的片段,将B(中双11)切成10mm的片段。然后,将2 种类型的切割段与切口对接接触,置于去分化介质上。
去分化培养基配方:4.4g/L MS 粉+1 mg/L 2,4-D+1 mg/L 6-BA+30 g/L 蔗糖+2.6g/L 植物凝胶。
1.3 去分化后的外植体再生
2 周后,分化的愈伤组织出现在诱导再生培养基中。将2 种类型的外植体转移到诱导再生培养基上。
再生培养基:4.4g/L MS 粉+4mg/L 6-BA+2 mg/L ZT+5mg/L AgNO3+30 g/L 蔗糖+2.6 g/L 植物凝胶。
1.4 外植体分化
2 周后,在切割段的切割位置出现分化的绿色和白色致密愈伤组织。2 种类型的片段一起转移到分化芽的培养基中,切割段的愈伤融合并一起生长。
分化芽培养基:4.4g/LMS 粉+3mg/L 6-BA+2mg/L ZT+30g/L 蔗糖+2.6 g/L 植物凝胶。
1.5 观察外植体和茎分化的生长
每2 周更换1 次培养基。观察分化培养基中愈伤组织的变化。愈伤分化成叶片相似的组织。如果没有出现类似叶子的组织,则仍然需要将培养的外植体置于分化芽培养基中培养。
分化茎培养基:4.4g/LMS 粉+0.005mg/L6-BA+30g/L蔗糖+2.6g/L 植物凝胶。
1.6 小苗驯化
当外植体在分化茎干培养基中,生长成具有4~5片叶子和3~4 根纤维根小的整合植物时,打开培养盆使幼苗适应外部环境。2~3d 后,取出植物,清洗根部周围的培养基,并将其移植到带有营养土壤的盆容器中。在充足的阳光下自然生长并定期施肥[5]。
1.7 用分子标记和表型测试筛选并确认阳性细胞系
对参与细胞质雄性不育系的Ogu89 和中双11,采用蔗糖密度梯度从黄化幼苗中提取线粒体DNA,在25℃黑暗中生长4d,提取总DNA 以对可育基因进行分子检测。Ogu 细胞质雄性不育系具有465bp 的简单序列重复(SSR)标记,其与调节Ogu 不育的Orf138 基因紧密连结。通过使用特异性引物检测,使用mtDNA 特异性引物扩增PCR。试验需要具有Ogu89 CMS 的核遗传物质和中双11 隐性生育系的正常细胞质的品系。
2 结果与分析
2.1 隐性CMS 保持系的开发
在MS 培养基上分别播种了80 粒Ogu89(不育系)和中双11(优良品系)种子。Ogu89 的77 粒种子和中双11 粒的50 粒种子长成了幼苗。采用组织培养法进行相似移植后,分化后得到156 个外植体。分化和再生过程后,成功转移了105 个外植体(67.3%)。最后,实现了约30 棵幼苗。
2.2 筛选并确认阳性植株
在Murashige 和Skoog(MS)培养基上分别播种了80 粒Ogu89(不育系)和中双11(优良品系)种子。Ogu89 的77 粒种子和中双11 粒的50 粒种子长成了幼苗。采用组织培养法进行相似移植后。去分化后得到156 个外植体。分化和再生过程后,成功转移了105 个外植体(67.3%)。最后,实现了约30 棵幼苗。
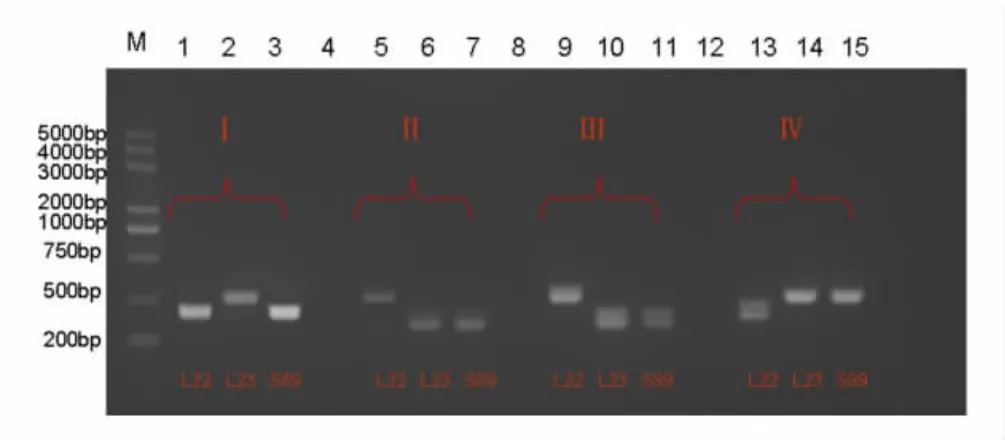
图1 使用分子标记检测转移的细胞质不育基因
Ogu 细胞质雄性不育系具有465bp 的简单序列重复(SSR)标记,与调节Ogu 不育的Orf138 基因紧密联系。通过Orf138 基因的特异性引物扩增的PCR 产物中出现了465bp 的条带(如图1)。显示了测试结果,然后验证了成功转换的真实性。需要的是具有Ogu89 CMS核遗传物质和中双11 隐性生育系正常细胞质的品系。结合表型性状和基因型鉴定的结果,最终获得了3 株具有正常生育力和农艺性状的植物,这些植物与雄性不育系相似。因此,1/10 的幼苗(3/30 幼苗)和1.9%外植体(3/156 外植体)可以从CMS 系转移到受体。
3 结论与讨论
在隐性CMS 系统中,大多数种质可以恢复不育系的繁殖力。传统方法则需要多次回交。然而,遗传物质在不育系和携带正常细胞质可育基因的恢复系之间的水平转移,可从后代中选择保持系,通过这种改良的组织培养方法获得,节省了大量种植,缩短繁殖时间,降低繁殖成本。
本研究中的技术(GTCM)可以快速实现CMS 基因从相关野生物种到栽培品种的低水平转移。将转化质体的靶基因从一个种质快速水平转移到另一个种质,由于质体转化具有独特的优点,它受到越来越多的关注。该GCTM 技术可以使具有转基因质体和非转基因品系的愈伤组织一起生长以形成整体,并分化成正常植物。经过分子和表型确认后,筛选成功转化的植物。
在试验过程中,会遇到以下问题:(1)组织培养中的操作必须小心和标准。应避免污染。否则可能导致整个试验失败。确保使用的所有设备都无菌。为了达到预期的结果,重复组织培养过程非常必要。
(2)影响培养细胞生长和芽再生的因素很多,如抗生素。根据现有方案,胚胎发生愈伤组织来自下胚轴胚发生培养物。胚胎发生愈伤组织在MS 培养基中培养,胚萌发和植物再生。MS 培养基由补充有6.8mM 谷氨酰胺,3.8mM 天冬酰胺,3%(w/v)蔗糖,0.3%植物凝胶和2μM6-BA,2μM2,4D 的无机盐组成。
总之,GTCM 技术使能够快速开发隐性CMS 的保持系,将不育基因从相关的野生物种转移到栽培品种,甚至在具有更远遗传距离和杂交不相容性的物种之间转移,并将转化质体的快速靶向基因,从一个种质转移到另一个是在低成本和高效率的条件下。
猜你喜欢
山东林业科技(2022年3期)2022-07-14
课外生活·趣知识(2022年2期)2022-02-08
烟台果树(2021年3期)2021-11-29
北方果树(2020年2期)2020-04-09
高考·中(2019年6期)2019-09-10
生物学教学(2018年5期)2018-11-29
课程教育研究·学法教法研究(2017年10期)2017-05-31
考试周刊(2016年81期)2016-10-24
科学中国人(2016年30期)2016-07-14
中学生物学(2016年6期)2016-07-04
