
黑曲霉高产β-葡萄糖苷酶菌株的诱变、筛选及发酵条件优化
2020-11-08 00:40孙智郭金玲龚大春古永红文晓敏刘君子肖玲玲
科学导报·学术 2020年87期
孙智 郭金玲 龚大春 古永红 文晓敏 刘君子 肖玲玲


【摘 要】文章主要是以黑曲霉TJ02为出发菌对其的孢子进行了硫酸二乙酯化学的诱变,同时对该菌株的发酵培养基进行了优化,然后以玉米芯为主要碳源,对该菌株中的β-葡萄糖苷酶的温度以及稳定性展开了测试。
【关键词】黑曲霉;β-葡萄糖苷酶;诱变
纤维素酶是一种多组分良好的酶系统。根据其诱导反应的程度,它具有重要的功能。内切葡聚糖酶(英文缩写EG)、外切葡聚糖酶和β-葡萄糖苷三种主要类型。酶(缩写为BG)协同作用促进纤维素的降解而不产生长纤维聚糖和醛糖。这些糖最终被水解成糖。此外,β-糖苷酶将醋酸时间内迅速释放的纤维结构半乳糖降解为肝糖原,促进其清除。抑制酶组分是水解过程的最后一步,也是受外界因素制约的最关键因素。纤维素酶的产生菌是木霉菌。所产酶系具有较高的斜葡萄糖寡糖和外源葡萄糖苷酶活性,但β-葡萄糖苷酶数量相对不足,生物降解效率和质量较低。米曲霉具有独特的高β-糖苷酶活性。在酶体系中加入该酶还可以大大提高富含纤维素的核桃木复合材的速度和效率。本研究通过硫酸二乙酯的化学诱变,获得了一株遗传基因相对稳定的菌株。优化了黑青霉β-糖苷酶高产突变株DES-7,并对其培养的碳源和氮源进行了优化。确定了β-葡萄糖苷酶的最适pH和最适室内温度,并测定了其pH稳定性和热稳定性。
1测定基本情况
1.1黑曲霉致死率的测定
用氯酸盐人工诱导tj02黑曲霉。随着渐进突变育种时间的延长,细胞质的死亡率会增加。硫酸乙酯多倍体时间为10min时,孢子囊死亡率仅为23.2%;诱变育种时间延长至30min时,孢子囊死亡率提高到85.3%;化学诱变时间为40min时,卵死亡率为95.8%;50 min和60min时,孢子囊死亡率为95.8%,死亡率在60%以上。根据死亡率选定硫酸二乙酯,后续治疗时间为30min[1]。
1.2高β-葡萄糖苷酶活菌株的诱变及筛选
用硫酸二乙酯处理的孢子囊悬浮液包被在PDA板上。随机选择25个突变株并将其疫苗接种在新的PDA平板设备上以进行重新分离和纯化。通过刮除诱变环获得的点突变菌株的孢子囊疫苗在次级发酵琼脂培养基中进行二次发酵和连续培养。确定β-葡萄糖苷酶以进行初步筛选。突变体des-7和des-11的酶活性大大提高。
1.3遗传稳定性
通过这种初步筛选获得的突变菌株DES-7和DES-11被连续培养了8代,并确定了亲本的遗传稳定性。DES-7的β-葡萄糖苷酶活性比突变体DES-11的稳定。与原始菌株TJ02相比,β-葡萄糖苷酶活性提高了30%以上[2]。
1.4碳源对β-葡萄糖苷酶产量的影响
通过使用麸皮,芝麻杆,玉米芯(Wangetal,2011),木糖残渣,油炸的麦秆,微晶纤维素等,将一系列最便宜易得的原料用作研究的主要原料,检测其对β-葡萄糖苷酶对菌株的影响。当玉米芯用作碳源时,分离株发酵液中的中间蛋白和β-葡萄糖苷酶的生物活性最大,前者为0.48g/L和13.78IU/mL。紧接着进行蒸汽喷射的小麦,稻秆和麸皮,其次是玉米。核心是唯一的碳源。研究了相同高浓度对细菌产生的β-葡萄糖苷酶的影响。以玉米芯为唯一底物,其浓度比对噬菌体产生的β-葡萄糖苷酶的失活有重大影响。当玉米芯的浓度为50g/L时,β-葡萄糖苷酶活性达到39IU/ml10。902菌株的生产力将随着玉米芯浓度比的增加而增加。这可能与丝状真菌的高密度菌丝有关[3]。如果底物缠结的浓度太高,则二次发酵液相对较厚,这不利于分离物的产生。与小麦核心(50g/L)作为碳源相比,使用了天然的初始发酵培养基(玉米芯核心麸为发酵底物时β-葡萄糖苷酶活作为28iu/ml),肠球菌的容量规模也很差。
1.5氮源对β-葡萄糖苷酶产量的影响
在恒温、氮气浓度不变的前提下,以(NH4)2SO4、(NH4)2CO、NH4COOCH3、NH4Cl、NaNO3、干酵母粉、小麦充分搅拌和玉米蛋蛋白胨为两种不同类型,8种成分成为研究芽孢杆菌对β-葡萄糖苷酶产量的影响、(NH4)2SO4和复合肥在无机物氮源中对细菌酶的产生比三种组分更为不利;考虑到市场因素,(NH4)2SO4是最有利的无机复合氮源;发酵粉是最好的无机相氮源,初始琼脂培养基中的氮源为(NH4)2SO4与酵母粉混合,发酵的最佳效果高于多种组分,说明(初始液体培养基中的(NH4)2SO4和酵母发酵粉效果。
1.6β-葡萄糖苷酶表观酶学性质研究
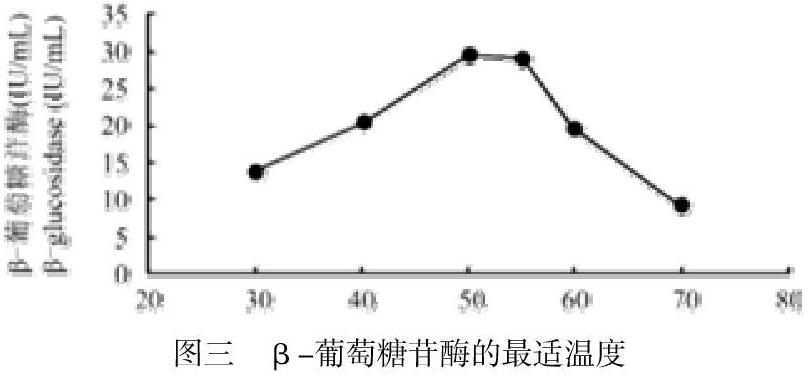
1.6.1 β-葡萄糖苷酶的最适温度
为了更好地确定β-葡萄糖苷酶的最佳温度,在不同摄氏度的基本条件下测量了β-葡萄糖苷酶的酶活性。当温度降至55°C时,酶的活性可以达到最高。观察β-葡萄糖苷酶活性的变化曲线:当总温度低于55℃时,酶活性随室内温度的升高而增加;当总温度为60℃时,酶活性降低。当摄氏温度超过70°C时,酶活性急剧上升,降至最佳总温度的30%。
1.6.2 β-葡萄糖苷酶的最适pH
在pH为2.0-9.0的相同缓冲空间溶液中,准确地测定了β-过氧化物酶的活性物质与pH的关系。由该菌株产生的β-葡萄糖苷的活性随pH而显着变化,并且其酶活性pH4.0左右是最高的,表明菌株所产的β-葡萄糖苷酶是一种弱酸性的β-葡萄糖苷酶,它的最佳pH值约为4.8。
1.6.3 β-葡萄糖苷酶的热稳定性
测定β-葡萄糖苷酶的热稳定性,并在37℃、50℃和65℃下測定β-葡糖苷酶的活性。β-葡萄糖苷酶的热稳定性在37℃时最好,24hβ-葡萄糖苷酶的活性为95%。温度到50摄氏度℃是β-葡萄糖苷酶的热稳定性最高,且24小时内β-葡萄糖苷酶的活性为90%。温度为65时℃β-葡糖苷酶的稳定性较差,β-葡糖苷酶的全活性在6小时内降至0。
1.6.4 β-葡萄糖苷酶的pH稳定性
在不同pH条件下于50℃保温2h后,测定β-糖苷酶的相对酶活性。在50℃加热2h后,β-糖苷酶在pH 4.0至6.0具有良好的稳定性,酶活性为大于85%。在pH 8.0的条件下,在50℃下保温2小时后,酶活性仅为原始酶的50%。
2讨论
β-葡萄糖苷酶也称为葡糖苷酶水解酶,也称为龙胆二糖酶,长纤维二糖酶和苦杏仁苷酶。它属于蛋白水解酶的类别。来自相同来源的芥子油苷酶的分子量和范围从数十kDa到数百kDa。葡糖苷酶最早是在1837年由Liebig和Wohler在苦腰果中发现的。作为纤维酶的组成部分,芥子油苷的主要基本功能是使非真实的还原性糖苷键与糖链末端复合,并根据实际情况释放葡萄糖和配体。另外,β-葡萄糖苷酶可以弱化对硝基苯半乳糖不耐受性和脱氢肋骨内酯,这在生产实践中起着重要作用[4]。β-葡萄糖苷酶是纤维素酶系统中最重要的酶。当乙酸纤维蛋白酶富含纤维素时,它的重复使用和葡萄糖苷酶的缺乏将导致大量纤维素二糖积聚。乳糖的大量积累实际上将对酶促反应形成非常强的即时反馈抑制作用。纤维素酶水解并显着增强果胶酶。系统中葡萄糖苷酶的活性对于提高果胶水解产物的速度和效率以及增加葡萄糖的年产量最为重要。因此,对葡萄糖苷酶的研究仍具有重要的理论和实用性。
黑曲霉tj02氯酸盐的多种化学诱变方法导致了β-葡萄糖苷酶突变体des-7,并具有较高的稳定性。与原始菌株相比,des-7的酶生产能力大大提高了至少30%。在工业部门,菌株的生产能力是关键点之一,而因素也与产品生产的成本直接相关。分子遗传学技术的整体发展取决于基因工程技术。分离株转化的非凡成就取得了重大突破。因此,基因工程仅限于具有详细遗传基团信息和清晰代谢功能和途径的菌株,而遗传背景不明确的菌株是没有办法进行定向改造。相反,传统的诱变育种方法如紫外线诱变育种、理化诱变育种等,不需要受到遗传遗传学经济背景的限制。对硫酸二乙酯进行了化学和物理诱变育种,获得了持续高产的芽孢杆菌,这直接证明了传统的化学诱变方法更有效。菌株转化是利用单一重要因子实验结果对突变株DES-7的碳源和氮源进行充分优化的方法。当氮源为甜玉米芯,氮源为酵母粉和氯化铵时,β-葡萄糖苷酶活性为39IU/mL,比初始菌株提高了85.7%。确定了β-葡萄糖苷酶的最佳室内温度、最适pH值、热稳定性和pH稳定性。证明了β-糖苷酶在55℃和pH 5.0时酶活性最高,在37℃和50℃下保持良好的高可控性,在pH 4.0~6.0时稳定性较好。木质膳食纤维的蛋白质水解一般在50℃下进行,本研究获得的突变菌株半乳糖在50℃下能保持较高的稳定性,可加入到膳食纤维酶系中用于木质粗纤维的水解[5]。
3结语
β-葡萄糖苷酶已在我国许多行业中被广泛应用。它具有巨大的商业形式,潜力无限。发酵总体水平低一直是β-葡萄糖苷酶工業化为能生产的主要原因。当前对于提高β-葡萄糖苷酶对天然发酵过程进行了整体优化,并对菌株进行了改良。
参考文献:
[1]蒋辉,王吉龙. 取力型变速器:,2014.
[2]周红,张阳,宋育阳,et al. 不同启动子及锚定蛋白对酿酒酵母表面展示β-葡萄糖苷酶活性的影响[J]. 农业生物技术学报,2020(10).
[3]罗秀针,郑金华,林燕燕,等. 1株灰黄霉素产生菌的诱变选育及其发酵条件优化[J]. 福建农业科技,2020(6):12-16.
[4]周红,张阳,宋育阳,et al. 不同启动子及锚定蛋白对酿酒酵母表面展示β-葡萄糖苷酶活性的影响[J]. 农业生物技术学报,2020(10):1849-1861.
[5]喻帆,方佳双,应灵萍,等. 细菌叶绿素a高产菌株的选育和发酵工艺优化[J]. 中国抗生素杂志,2020(6):567-572.
