
象山港蓝点马鲛8种同工酶特征分析
2020-11-06 09:19:48郑保霄郑春静娄正家薛良义
海洋学研究 2020年2期
郑保霄,郑春静,张 玉,娄正家,李 娟,薛良义*
(1.宁波大学 海洋学院,浙江 宁波 315211;2.宁波市海洋与渔业研究院,浙江 宁波 315010)
0 引言
同工酶是指具有相同催化作用而分子结构不同的酶,普遍存在于动植物中[1-2]。同一物种的不同个体或同一个体的不同组织都可能具有不同的同工酶分布。通过分析同工酶的表型而获知其基因型是生化遗传分析的手段之一,可用于遗传多态性座位的研究[3]。目前已有对多种鱼类同工酶从不同角度的研究报道,在组织特异性方面,金春华 等[4]检测了多种同工酶在大弹涂鱼(Boleophthalmuspectinirostris)肝脏中的表达,发现三梨醇脱氢酶(SDH)和葡萄糖6磷酸脱氢酶(G6PDH)仅在肝脏中表达;徐钢春 等[5]分析了刀鲚(Coilianasus)不同组织的乳酸脱氢酶(LDH)的表达,发现眼睛是刀鲚LDH表达较为典型的组织;张涛 等[6]研究了双须叶须鱼(Ptychobarbusdipogon)5种组织中苹果酸脱氢酶(MDH)和LDH同工酶的表达特征,结果表明2种酶具有组织特异性。在性别特异性方面,庞秋香 等[7]研究了泰山赤鳞鱼(Varicorhinusmacrolepis)苹果酸酶(MEP)的表型差异,发现不同性别同一组织和同一性别不同组织之间都存在明显差异,但在同一性别相同组织的不同个体之间不存在差异。在环境影响方面,曹永长 等[8]利用电泳法分析了鲮鱼的酯酶(EST),发现EST酶带在不同温度下发生了变化,推测鲮鱼体内的脂肪代谢在温度适应过程中发生了变化;黄福勇 等[9]研究了凫溪香鱼(Plecoglossusaltivelis)的15个同工酶,发现过氧化氢酶(CAT)、超氧化物歧化酶(SOD)和过氧化物酶(POD)3种同工酶可以结合来评价生理生化、环境因子对香鱼机体保护系统的影响;LENRTOVet al[10]分析了环境污染对超氧化物歧化酶(SOD)的影响,在来自环境严重污染地区的鱼类肝脏中发现了新的SOD亚型;KUZU et al[11]研究了重金属对鱼鳃中的碳酸酐酶Ⅰ和Ⅱ(CAⅠ和CAⅡ)同工酶活性的影响,结果表明Cu2+,Ag+,Cd2+和Ni2+离子对其均有抑制作用。在种质鉴定方面,万正义 等[12]通过分析长江四大家鱼肌肉组织的LDH同工酶来进行种质鉴定;陈玮彤 等[13]通过检测渤海湾天津沿岸花鲈(Lateolabraxmaculatus)肝脏和肌肉组织中的7种同工酶来分析其遗传多样性。
蓝点马鲛(Scomberomorusniphonius),又名鲅鱼,隶属于鲈形目、鲭科、马鲛属,属暖水性中上层鱼类,是东海、渤海和黄海地区主要的经济鱼类资源之一。象山港位于浙江省中北部沿海,港内营养盐含量较高,饵料生物繁富,是蓝点马鲛在东海的自然繁殖场[14]。近年来因过度捕捞导致象山港海域的马鲛鱼资源出现衰退,因此研究和保护蓝点马鲛种质资源显得尤为重要。近年来,国内外学者对蓝点马鲛鱼的研究多集中于洄游路线分析[15]、重要功能基因[16-17]、微卫星位点[18]以及线粒体基因组等方面[19]。同工酶是在蛋白质水平上研究物种遗传多样性,具有稳定的特征,目前仍然是我国进行生物种质鉴定的标准方法之一。本研究采用垂直聚丙烯酰胺凝胶电泳,对象山港水域蓝点马鲛9种组织中的8种同工酶进行分析,为蓝点马鲛的遗传多样性和种质鉴定提供参考,为该渔业资源的保护和开发提供科学依据。
1 材料与方法
1.1 样品采集与处理
实验所用的蓝点马鲛鱼于2017年4月中旬取自浙江省宁波市象山港,为近海即时捕捞的性腺成熟的活鱼,雌雄各3条,全长(623.5±55.5) mm。活体解剖,每尾鱼取心脏、鳃、肠、肌肉、胃、肝、眼、性腺和肾脏9种组织,分别装于已编号的2 mL冻存管中,迅速投入液氮保存,带回实验室-80 ℃保存至分析。取待分析的样品各0.1 g,加入4倍量(v/w)的Tris-HCl缓冲液(0.01 mol/L,pH=6.8左右),冰上匀浆后离心(4 ℃,13 000 r/min,30 min),取上清液用于电泳。
1.2 电泳、染色及成像
采用聚丙烯酰胺梯度凝胶电泳(Gradient Polyacrylamide Gel Electrophoresis,GPGE)对8种同工酶进行分析。凝胶制备方法参考文献[20],组织化学染色方法参照文献[21],染色液配制见表1。用上述配置的染色液分别染色,步骤如下:电泳完成后,将凝胶取出,放入染色液中,LDH、MDH、葡萄糖脱氢酶(GDH)、G6PDH、EST、SDH和MEP的显色需要在37 ℃避光20~30 min;SOD的显色需要曝光3~5 min,在37 ℃温育至有灰白色条带出现。染色后的凝胶放入7%冰乙酸中固定脱色。
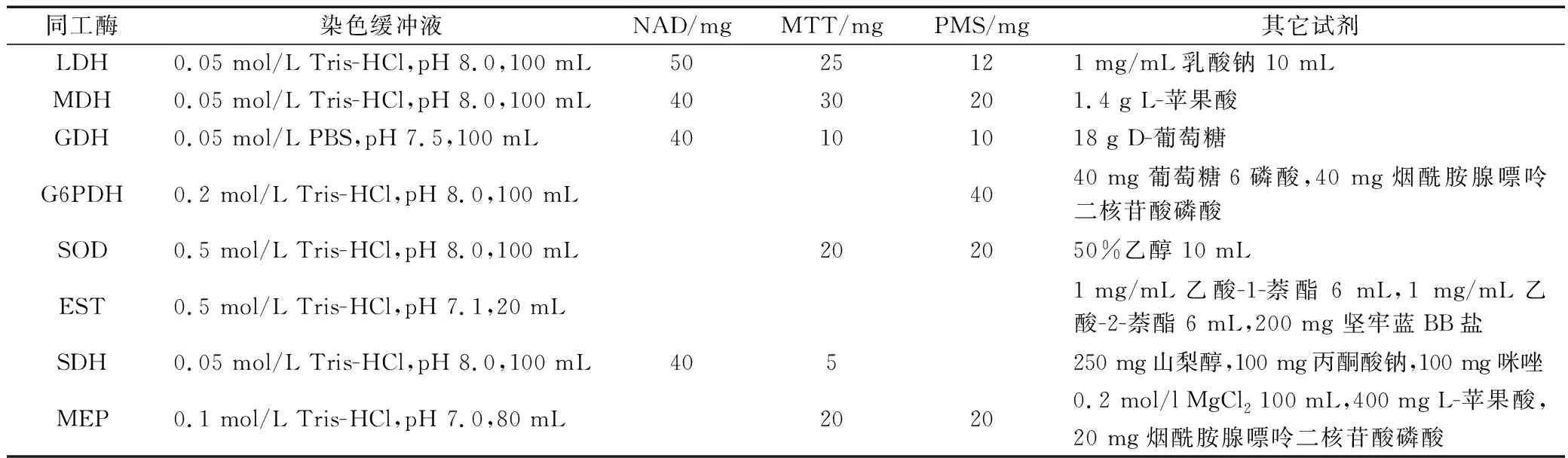
表1 同工酶染色液配方Tab.1 The formulation of isoenzyme staining solution
脱色后将凝胶板进行拍照,并根据电泳图谱结合相对迁移率和表达活性程度绘制酶谱模式图,模式图中的条带颜色深浅与电泳酶带活性强弱相对应。
1.3 电泳结果记录
同工酶的缩写、基因座位和等位基因的命名采用SHAKLEE et al[22]推荐的方法,以同工酶缩写名称的大写代表酶蛋白。
2 结果
实验共检测了乳酸脱氢酶(LDH)、苹果酸脱氢酶(MDH)、葡萄糖脱氢酶(GDH)、葡萄糖6磷酸脱氢酶(G6PDH)、超氧化物歧化酶(SOD)、酯酶(EST)、山梨醇脱氢酶(SDH)、苹果酸酶(MEP)等8种同工酶在各器官组织中的表达模式,检测结果见表2。
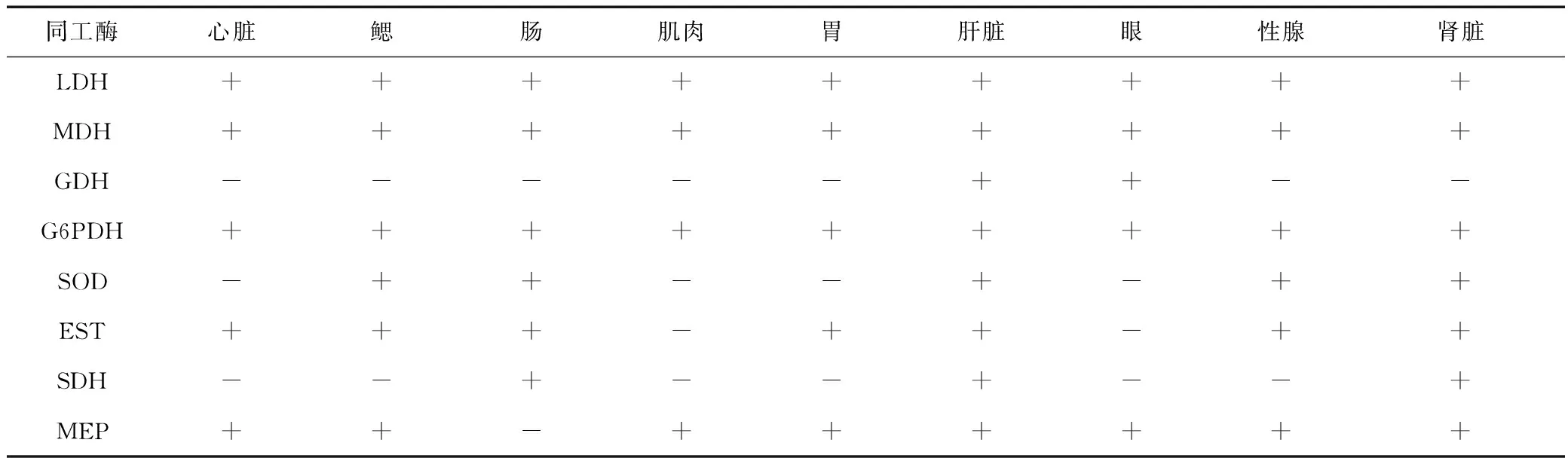
表2 同工酶在蓝点马鲛9种组织中的表达Tab.2 The expression of isoenzymes in nine tissues of Scomberomorus niphonius
2.1 各组织中的酶谱特征
2.1.1 乳酸脱氢酶(LDH E .C.1.1.1.27)
在蓝点马鲛9种组织中共检测到7种LDH酶带(图1a)。LDH1仅在肌肉组织中表达,其他组织中均不表达;LDH2在9种组织中都表达,在肌肉和肝脏中活性较弱;LDH3~LDH7只在眼睛中表达。
2.1.2 苹果酸脱氢酶(MDH E .C.1.1.1.37)
在蓝点马鲛9种组织中共检测到7种MDH酶带(图1b)。MDH1和MDH2仅分别在肠和性腺中表达;MDH3在心脏和肌肉中表达;MDH4在肠、胃、肝脏、眼和肾脏五个组织中表达,其在肠和眼睛中活性较弱;MDH5在除肠和眼睛外的其余7个组织中均有表达;MDH6在心脏、鳃、肠、肌肉和性腺中表达;MDH7在心脏、肌肉和性腺中表达,活性较弱。
2.1.3 葡萄糖脱氢酶(GDH E.C.1.4.1.2)
由图1中a可知,2种酱油在40~70天内总氮呈总体上升趋势,且pH值为5.4的酱油中含有更高的总氮值。pH 5.4条件下发酵的酱油总氮值由40天时的1.44 g/dL逐渐升高,至发酵55天时达到约1.53 g/dL,此后总氮含量逐渐趋于稳定。而pH 6.5条件下发酵的酱油总氮则由40天时的约1.4 g/dL逐渐升高至约1.5 g/dL,略低于pH 5.4条件下发酵的酱油。
在蓝点马鲛9种组织中共检测到3种GDH酶带(图1c)。GDH1仅在眼中检测到,活性较弱;GDH2和GDH3仅在肝脏中检测到,其余组织中均未检测到相关酶带。
2.1.4 葡萄糖6磷酸脱氢酶(G6PDH E.C.1.1.1.49)
在蓝点马鲛9种组织中共检测到3种G6PDH酶带(图1d)。G6PDH1在除鳃外的其他组织均有表达;G6PDH2在鳃、肝脏和性腺中表达,G6PDH3仅在肠和肾脏中表达。G6PDH1和G6PDH2均在肝脏中的活性最强。
2.1.5 超氧化物歧化酶(SOD E.C.1.15.1.1)
在蓝点马鲛9种组织中共检测到1种SOD酶带(图1e),在鳃、肠、肝脏、性腺和肾脏表达,在肝脏中活性最强。
2.1.6 酯酶(EST E .C.3.1.1.1)
在蓝点马鲛9种组织中共检测到5种EST酶带(图1f)。EST1在除肌肉和眼睛外其余7种组织中均有表达,在鳃和肠中的活性较弱;EST2仅在性腺中有表达;EST3和EST4的组织特异性相似,在心脏、鳃、肠、胃、肝脏和肾脏中均有表达;EST5仅在肝脏和肾脏中表达,且活性较弱。
2.1.7 山梨醇脱氢酶(SDH E.C.1.1.1.14)
在蓝点马鲛9种组织中共检测到3种SDH酶带(图1g)。SDH1和SDH2仅在肝脏中表达,且活性较强;SDH3仅在肠和肾脏中表达,在肾脏中活性较强。
在蓝点马鲛9种组织中共检测到5种MEP酶带(图1h)。MEP1在心脏、肌肉、胃、肝脏、性腺和肾脏等组织中均有表达;MEP2仅在鳃中表达;MEP3仅在肝脏中表达;MEP4在心脏和性腺中表达;MEP5在心脏、肝脏、眼和性腺中有表达,但活性都较弱。
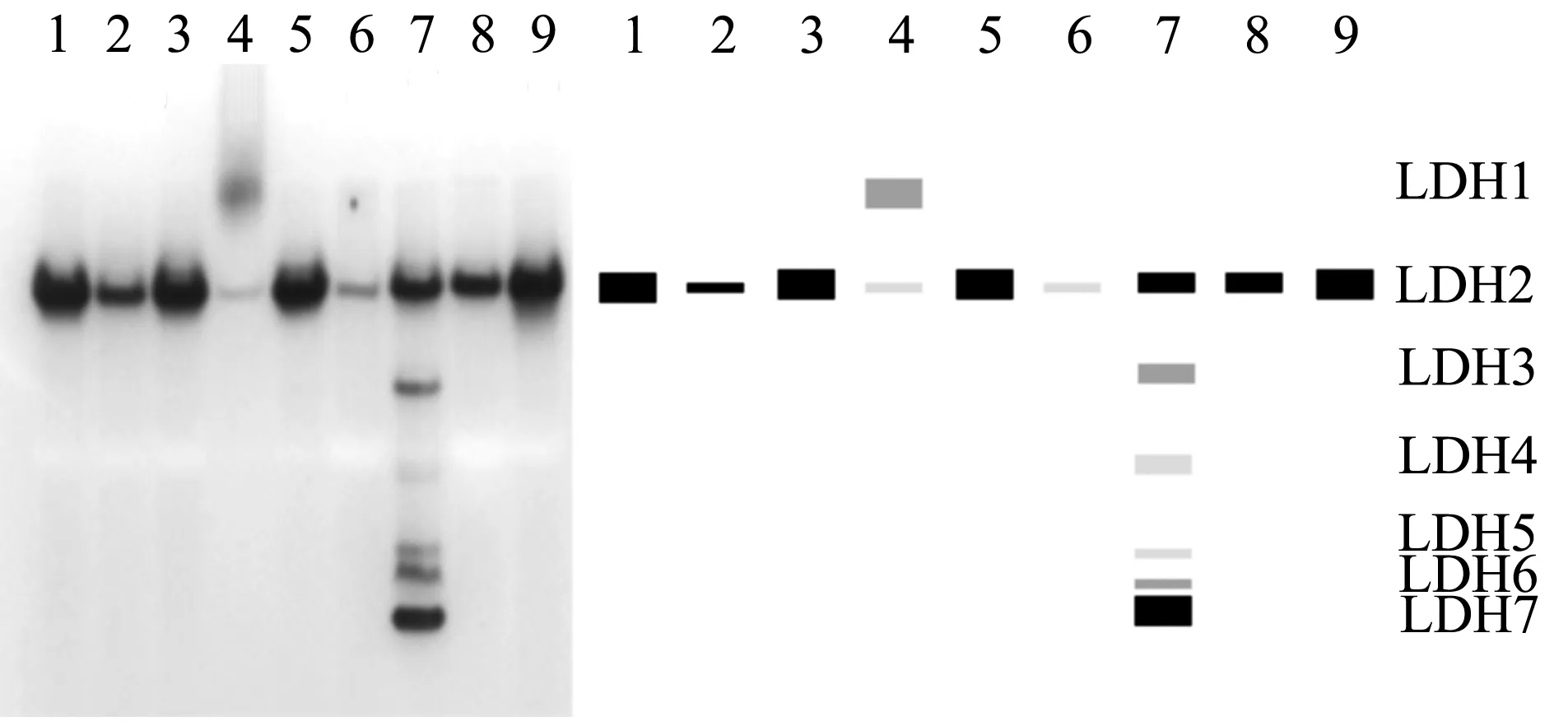
2.2 蓝点马鲛雌雄个体组织的同工酶谱比较
比较雌雄个体LDH同工酶在9种组织中的表达模式(图2),发现LDH同工酶的表达在雌雄个体间没有差别。其他同工酶在雌雄个体中的表达类似。
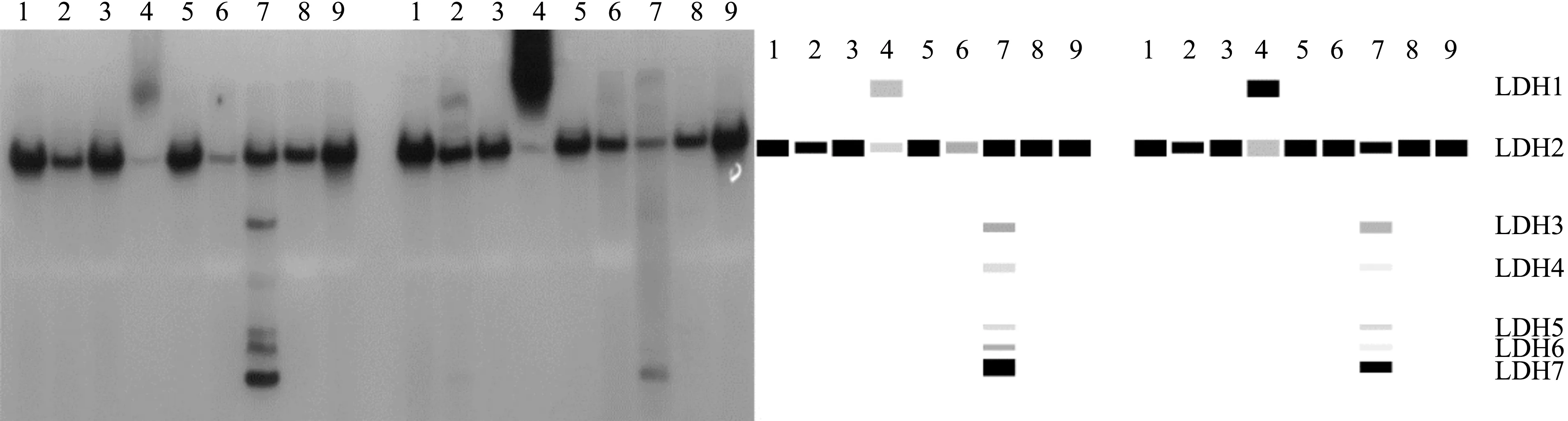
图2 蓝点马鲛LDH同工酶雌雄对比电泳图谱(左雌右雄)Fig.2 Electropherogram comparision of Scomberomorus niphonius LDH isozymes between male and female(female on the left and male on the right)(图谱上方数字分别表示:1—心脏;2—鳃;3—肠;4—肌肉;5—胃;6—肝脏;7—眼睛;8—性腺;9—肾脏。)(The numbers at the top of the graph indicate: 1—heart; 2—gill; 3—intestine; 4—muscle; 5—stomach; 6—liver; 7—eye; 8—gonad; 9—kidney.)
3 讨论
3.1 蓝点马鲛同工酶表达具有组织特异性
通过对蓝点马鲛9种组织中8种同工酶进行研究发现,同工酶表达的组织特异性主要表现在两个方面,一是在同一组织中不同的酶活性不同。在肾脏中未检测到GDH,其余7种酶均能检测到,但活性差别较大。在鳃和性腺中未检测到GDH和SDH,而其余6种酶均能检测到。在肝脏中8种同工酶均能检测到,LDH和SOD的活性不高,而其余6种同工酶的活性很高。在对卵形鲳鲹(Trachinotusovatus)的研究中发现,在肝脏中,EST、MDH和ME均能检测到,并且活性很高,而LDH的活性很低[23]。在对鲈鱼的研究中也有类似的发现[3]。马鲛鱼、卵形鲳鲹和鲈鱼都属于鲈形目,说明在鲈形目鱼类中,同一组织中存在的酶系统的进化是相对保守的。在肝脏中检测到多种的同工酶,可能是因为肝脏是重要的物质代谢场所,行使复杂的生理功能。二是同一种酶在不同组织中的表达存在差异。例如LDH,MDH和G6DPH在9种组织中都可以检测到,但表达强度相差很大。LDH在肌肉和肝脏中的活性较弱;在大多数组织中只有1个条带,但在眼睛中检测出多个条带,表明LDH同工酶在眼中存在多个结构形态。MDH在眼睛中的表达与其他组织相比较弱,在心脏,肾脏和肝脏等组织中活性较强。G6DPH在肝脏中的活性较强,在其余8种组织中的活性都很弱。同工酶在不同组织中表达存在差异的情况也存在于其它鱼类,如鲈鱼[3]、大弹涂鱼[4]等。
3.2 蓝点马鲛同工酶表达的特异性与功能的关系
LDH为糖酵解酶,能催化丙酮酸生成乳酸[24],由A、B和C 3个基因位点编码,LDH-A编码的酶多在厌氧组织中表达,LDH-B编码的酶多在含氧组织中表达[25-26],LDH-C基因存在组织特异性[27]。LDH是由A、B两个亚基随机组合成的四聚体,不同结构形式在电泳时形成各种不同的条带。LDH在蓝点马鲛9种组织中共检测出7条酶带,LDH1只在肌肉中检测到,可能是由LDH-A基因编码形成的,催化丙酮酸转化为乳酸,参与无氧代谢。LDH2在9种组织中均有表达,LDH3~LDH7只在眼睛中检测到,并且LDH7的活性很高,推测LDH7是由LDH-C基因编码形成,LDH3~LDH6为杂合带。
MDH为二聚体酶,主要参与三羧酸循环过程。MDH分为上清液型(s-MDH)和线粒体型(m-MDH),分别由2个等位基因控制,m-MDH主要使苹果酸脱氢,s-MDH主要使胞液中的草酰乙酸还原而穿入线粒体[28-29]。电泳条件下s-MDH的泳动速度比m-MDH快[6]。在本实验中MDH在蓝点马鲛9种组织中共检测出7条酶带,MDH1~MDH4为线粒体型,MDH5~MDH7为上清液型。MDH在心脏、肝脏和肾脏等多种组织含量丰富,其中m-MDH在肌肉、心脏、胃、肝脏、性腺和肾脏中活性较强,这与肌肉等组织在三羧酸循环过程中把苹果酸转为草酰乙酸所需要的大量能量密切相关,而糖原能通过酵解和有氧分解迅速释放大量能量[4]。
GDH属于短链脱氢酶,它具有4个相同亚基,主要存在于动物的肝脏[30-31],在高等动物脊椎动物中GDH只在肝脏中表达[30],能够催化D-葡萄糖成为D-葡萄糖酸内酯,同时还原NAD(P)+为NAD(P)H。在本实验中,GDH虽然在蓝点马鲛的肝脏和眼组织中检测到,但主要在肝脏中表达,在眼中活性较弱。
G6DPH为二聚体,是磷酸戊糖途径的关键酶,协助葡萄糖进行新陈代谢,催化NADP还原生成NADPH[32]。在蓝点马鲛9种组织中共检测到3种G6PDH酶带,其中肝脏的G6DPH活性最强,其次是肾脏,在其他组织中的活性较弱,这可能与肝脏和肾脏具有旺盛的糖代谢功能相关。在动物骨骼肌中基本缺乏磷酸戊糖途径,而在脂肪组织、肾上腺皮质中,大部分葡萄糖是通过此途径分解的,肝脏中也可通过此途径进行糖代谢[32],因此在肝脏、肾脏以及脂肪等组织中可能含有较多的G6DPH。
SOD为二聚体酶,是一种重要的防御酶,其功能主要是清除机体的超氧自由基,维持机体正常的生理功能[33]。在蓝点马鲛9种组织中只检测到1种SOD酶带,与其他鱼类中所报道的多条酶带有所差异,如在松江鲈鱼(Trachidermusfasciatus)7种组织中检测到3种酶带[34],这可能与同工酶在不同物种中的差异性表达有关。SOD酶在肝脏组织中的活性最强,可能与其在肝脏中参与各种生物分子旺盛代谢产生的氧自由基的清除有关。
EST为单体或二聚体酶,大多数为单体,通常由2个以上的等位基因控制,其多态现象非常普遍,条带多且复杂。EST是水解酶类,能催化脂类化合物水解,维持细胞正常的生理代谢[24]。在蓝点马鲛9种组织中共检测到5种EST酶带,其在肝脏和肾脏中的活性最强,具有明显的组织特异性,这可能与鱼类肝脏和肾脏中代谢旺盛,具有代谢废物和毒素解毒功能有关。
SDH能可逆地催化D-山梨醇氧化为D-果糖,在山梨醇代谢过程中发生重要作用,主要分布于肝脏、肾、脑、心和脾等组织中且含量极少,血清中此酶活力很低,如酶活力升高,强烈提示肝损害[35]。在蓝点马鲛9种组织中共检测到5种SDH酶带,仅在肠、肝脏和肾脏中有活性,其中在肾脏中活性最强,在肝脏中活性较弱,在肠中极弱,在其余组织中则不表达,说明在肾脏中SDH可能参与了较多的山梨醇代谢过程。
MEP是催化苹果酸生成丙酮酸的酶,它在胞液内催化苹果酸脱羧,其产生的NADPH可用于脂肪酸的合成,将糖酵解和柠檬酸循环联系起来[36],因此在机体代谢旺盛的器官和组织中含量较高。在蓝点马鲛9种组织中共检测到5种MEP酶带,在心脏、肝脏和肾脏中活性最高,这可能与这些组织物质代谢旺盛有关。MEP在鳃和眼中的含量极低,说明在这2种组织中,可能不需要大量的糖代谢和脂肪酸合成。
3.3 不同性别马鲛鱼的同工酶谱比较
在本研究中,不同性别的蓝点马鲛8种同工酶在9种组织中的表达酶谱没有差别。孔晓瑜 等[37]对鲈形目鳜亚科9种鱼类的LDH同工酶进行了比较研究,发现LDH同工酶表型具有明显的种间差异,但同种的个体及性别之间没有明显差异。李凯彬 等[38]在研究鳉形目的RR-B 系剑尾鱼(Xiphophorushelleri)时发现,不同性别的剑尾鱼其LDH酶谱和GDH酶谱是一致的。熊全沫 等[39]对鲤形目中国胭脂鱼(Myxocyprinusasiaticus)的5种酶进行研究,其酶谱没有性别的差异,这些研究与我们的结果一致。而庞秋香 等[7]对鲤形目的泰山赤鳞鱼的研究表明,MEP的表型在不同性别间存在明显差异。庞秋香 等[40]对文昌鱼目的文昌鱼(Branchiostomabelcheritsingtauense)研究发现MEP、MDH和酸性磷酸酶(EAP)表型在雌性和雄性个体之间存在差异,认为这3种酶的合成可能是受性别连锁基因控制或是受激素和不同的生理状态控制。同工酶都是基因表达的产物,其表达调控受多种因素影响,在本研究中的8种同工酶在蓝点马鲛中的表达可能不受性别连锁基因的控制。
4 结论
通过垂直聚丙烯酰胺凝胶技术,对象山港蓝点马鲛9种组织中8种同工酶的酶谱分析表明,MDH、EST和MEP三种同工酶在组织中表达具有多态性,GDH和SDH只在特定的组织中表达,而雌雄个体组织间同工酶的表达没有差别。同工酶表达的组织特异性主要表现在两个方面:一是在同一组织中不同的酶活性不同,二是同一种酶在不同组织中的表达有差异。这几种同工酶的差异性表达可为蓝点马鲛的种质鉴定和遗传学分析等方面提供理论依据。
猜你喜欢
遗传(2023年8期)2023-08-25 06:59:00
数学大王·中高年级(2022年12期)2022-12-11 04:11:25
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
心肺血管病杂志(2018年11期)2018-12-18 01:51:40
中国医药生物技术(2015年4期)2015-12-26 08:26:36
天津造纸(2014年3期)2014-08-15 00:42:04
河南医学研究(2014年3期)2014-02-27 14:51:59
茶叶通讯(2014年2期)2014-02-27 07:55:39
小朋友·快乐手工(2014年2期)2014-02-25 12:22:25
食品科学(2013年23期)2013-03-11 18:30:10
