
1-脱氧野尻霉素对肥胖小鼠肝细胞线粒体合成与自噬能力的改善作用
2020-10-29 06:17李思远宁俊丽丁晓雯黄先智
食品科学 2020年19期
李思远,宁俊丽,丁晓雯,黄先智
(1.西南大学食品科学学院,重庆 400716;2.西南大学 家蚕基因组生物学国家重点实验室,重庆 400716)
肥胖是一种由多种因素导致的全球性慢性疾病,常起因于能量摄入与消耗失衡,给机体带来炎症、氧化应激、线粒体功能障碍等不利影响,同时也是糖尿病、心血管疾病等的主要风险因素,已被世界卫生组织认定为影响人类健康的五大危险因素之一[1-4]。
线粒体在能量代谢过程中发挥十分重要的作用,可通过脂肪酸β氧化促进机体脂代谢以维持能量平衡[5]。在正常情况下机体内健康线粒体的数量通过新线粒体的合成及衰老、损伤线粒体的自噬作用保持数量上的动态平衡,而在肥胖条件下,这种平衡被打破,线粒体的合成及自噬能力均受到抑制[6-8],导致新线粒体合成减少,衰老、损伤线粒体增加,脂肪酸氧化能力下降。Flamment等[9]的研究表明,高脂饮食导致小鼠肝脏线粒体生物合成关键调控因子过氧化物酶体增殖物激活受体γ共激活剂1α(peroxisome proliferator activated receptor γ coactivator-1α,
PGC-1α)mRNA表达下调,抑制线粒体合成。Ha等[10]发现肥胖小鼠肝脏中PGC-1α及其下游基因核呼吸因子1(nuclear respiratory factor 1,Nrf1)mRNA表达下调,同时,线粒体中脂肪酸β氧化关键酶——肉碱脂酰转移酶1(carnitine palmitoyltransferase 1,CPT1)的mRNA表达也显著下调,认为肥胖影响机体对线粒体的合成能力,同时降低线粒体对脂肪酸的代谢能力。此外,肥胖还通过影响线粒体自噬关键蛋白unc-51样激酶1(unc-51 like kinase 1,ULK1)抑制衰老损伤线粒体的自噬[11]。鉴于线粒体在脂代谢方面的重要作用,提高肥胖个体的线粒体生物合成与自噬能力对于肥胖的改善甚至治疗具有重要意义。
1-脱氧野尻霉素(1-deoxynojirimycin,DNJ)是一种主要从植物桑中分离得到的哌啶类生物碱,目前研究已经发现其具有降糖、降脂、抗肿瘤、抗病毒等生理活性[12-14]。在脂代谢方面,本团队前期的研究发现DNJ降脂机理主要为调节脂代谢相关酶活性,抑制脂肪酸合成,促进脂肪酸β氧化[15]。肝脏在脂代谢过程中扮演重要角色,线粒体是脂肪酸β氧化的重要场所,本实验以高脂膳食诱导的肥胖小鼠为实验材料,研究DNJ对肥胖小鼠肝脏细胞线粒体生物合成与自噬的影响,进一步探究其减肥机制,为后续更深入研究DNJ对肥胖小鼠线粒体的影响以及DNJ相关产品的开发提供依据。
1 材料与方法
1.1 动物、材料与试剂
140 只4 周龄清洁级昆明种小鼠,雌雄各半,生产许可证号为SCXK(渝)2018-003,购于重庆恩斯维尔生物科技有限公司。
基础饲料购于重庆恩斯维尔生物科技有限公司;高脂饲料为实验室自行配制[16-17],含基础饲料78.8%(质量分数,下同)、猪油10%、蛋黄粉10%、胆固醇1%、胆酸钠0.2%。
DNJ(纯度≥98%) 南京道斯夫生物技术有限公司;脂联素测定试剂盒、成纤维细胞生长因子21(fibroblast growth factor 21,FGF21)测定试剂盒、肿瘤坏死因子α(tumor necrosis factor α,TNF-α)测定试剂盒厦门慧嘉生物技术公司;白细胞介素-6(interleukin 6,IL-6)测定试剂盒、诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)测定试剂盒、PGC-1α蛋白水平测定试剂盒、Nrf1蛋白水平测定试剂盒、ULK1蛋白水平测定试剂盒、CPT1活性测定试剂盒 上海优选生物技术有限公司;二喹啉甲酸法总蛋白测定试剂盒 南京建成生物工程研究所;总RNA提取试剂盒 北京百泰克生物技术公司;反转录试剂盒、实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qPCR)SYBR Green染料试剂盒 上海翊圣生物科技公司。
1.2 仪器与设备
5810R型台式冷冻高速离心机 德国Eppendorf公司;EPOCH2型全波长酶标仪 美国基因公司;QTOWER 3G qPCR仪 德国耶拿分析仪器公司;FM200型匀浆机 上海弗鲁克公司。
1.3 方法
1.3.1 DNJ灌胃液的配制
根据实验室前期的研究结果,将DNJ用生理盐水溶解,配制成质量浓度分别为0.2、0.4、0.8 mg/mL的DNJ溶液,4 ℃保存备用。
1.3.2 肥胖小鼠模型的建立
140 只4 周龄昆明种小鼠(雌雄各半),适应性喂养1 周后,随机挑选20 只(雌雄各10 只)作为对照组,继续饲喂基础饲料;其余120 只作为造模组饲喂高脂饲料6 周,根据下式计算肥胖度。肥胖度大于20%认为造模成功[18]。

1.3.3 动物分组及饲养
随机选择造模成功的肥胖小鼠80 只(雌雄各半)分为4 组(每组20 只,雌雄各半),分别为肥胖对照组(饲喂高脂饲料,灌胃生理盐水)和DNJ高、中、低剂量组(饲喂高脂饲料,同时分别以8.0、4.0、2.0 mg/(kgmb·d)剂量灌胃DNJ),以1.3.2节中的对照组作为正常对照组(饲喂基础饲料,灌胃生理盐水)。各组每天灌胃1 次,持续灌胃45 d。实验期间小鼠自由摄食、饮水,每5 d称1 次体质量。控制室温(22±2)℃,相对湿度(50±15)%,12 h轮换照明(8∶00~20∶00)。
1.3.4 样本采集及指标测定
各组小鼠灌胃到第45天,禁食不禁水12 h,称量体质量后眼球取血于真空采血管中,室温放置1 h后,于4 ℃、3 000 r/min离心10 min,分离上清液即为血清[19],于-80 ℃保存备用。血清脂联素、FGF21、TNF-α、IL-6、iNOS水平严格按照试剂盒说明书操作步骤测定。
取肝脏称质量后迅速置于预冷的生理盐水中漂洗去除表面血污,取出,吸干表面水分,称取部分肝脏,按照1∶9(m/V)加入磷酸盐缓冲液(pH 7.4)于冰上进行匀浆,于4 ℃、3 000 r/min离心15 min,取上清液,得质量分数10%肝匀浆,-80 ℃保存备用。按照试剂盒说明书的方法测定肝脏组织总蛋白、PGC-1α、Nrf1、ULK1蛋白水平以及CPT1活力。
1.3.5 肝脏总RNA提取及反转录
按照试剂盒方法提取肝组织RNA,测定A260nm/A280nm检测其纯度,并通过琼脂糖凝胶电泳观察条带检测其质量,之后按照试剂盒方法反转录为cDNA。
1.3.6 qPCR扩增
反应体系按照SYBR Green染料说明书操作配制,扩增程序:预变性95 ℃,5 min;变性95 ℃、10 s,退火55~60 ℃、20 s,延伸72 ℃、20 s,40 个循环。扩增结束后根据熔解曲线评估引物特异性,根据扩增Ct值计算2-ΔΔCt得到mRNA相对表达量。引物序列见表1。基因 正向引物(5’-3’) 反向引物(5’-3’)
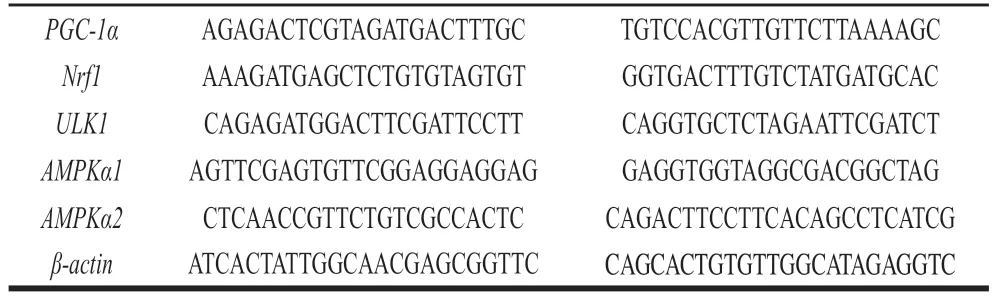
表1 qPCR引物序列Table 1 Primer sequences used for quantitative real-time polymerase chain reaction
1.4 数据统计分析
2 结果与分析
2.1 DNJ对肥胖小鼠体质量的影响
体质量的增加是肥胖的外在体现,机体摄入的能量过多时,多余能量便以脂肪的形式储存,导致体质量的增加,控制体质量的增加是防止或者缓解肥胖的重要方式。

表2 DNJ对肥胖小鼠体质量的影响(n=10)Table 2 Effect of DNJ on body mass in obese mice (n= 10)
由表2可知,DNJ各剂量组雌、雄鼠初始体质量与肥胖对照组雌、雄鼠初始体质量之间无显著差异(P>0.05),且均高出正常对照组体质量20%(P<0.05)以上,表明肥胖模型造模成功。灌胃结束后,肥胖对照组雌、雄鼠体质量显著高于正常对照组(P<0.05),而经过低、中、高剂量DNJ干预的雌、雄鼠体质量虽然仍显著高于正常对照组(P<0.05),但同时分别低于肥胖对照组体质量9.27%、9.08%、10.99%(P<0.05)和7.20%、6.34%、10.19%(P<0.05)。表明在实验剂量范围的DNJ能够缓解高脂饮食引起的肥胖小鼠体质量增加,维持体质量稳定,但在实验的时间范围内并不能使之恢复到正常水平。
2.2 DNJ对肥胖小鼠血清脂联素及FGF21的影响
脂联素是一种由脂肪组织分泌的细胞因子,具有促进线粒体合成、改善脂代谢的功能[20-21];FGF21主要在肝脏表达,其主要作用是促进糖、脂分解代谢,维持能量平衡,同时还有研究发现其能通过调节PGC-1α促进线粒体合成[22-23]。

表3 DNJ对肥胖小鼠脂联素及FGF21质量浓度的影响(n=10)Table 3 Effect of DNJ on adiponectin and FGF21 level in obese mice (n= 10)
如表3所示,与正常对照组比较,肥胖对照组雌、雄小鼠脂联素水平分别下降33.01%、32.35%(P<0.01),表明肥胖导致脂联素分泌下降,影响机体脂代谢。与肥胖对照组相比,中、高剂量DNJ的干预分别使雌鼠脂联素水平上升22.58%、43.19%(P<0.01),具有明显的量效关系且高剂量组脂联素水平恢复到正常水平(P>0.05);中、高剂量组雄鼠脂联素水平分别上升25.87%、29.58%(P<0.01),均未恢复至正常水平(P<0.05);低剂量DNJ对雌、雄鼠脂联素水平均无显著影响。
与正常对照组比较,肥胖对照组雌鼠的FGF21水平下降35.02%(P<0.01),表明肥胖同样导致雌鼠FGF21的分泌下降。与肥胖对照组比较,低、中、高剂量DNJ干预使雌鼠FGF21水平分别上升16.04%(P<0.05)、26.28%(P<0.01)、37.03%(P<0.01),均未恢复至正常水平(P<0.05);与此不同的是,雄鼠肥胖对照组的FGF21水平相比正常对照组上升32.98%(P<0.01),可能发生FGF21抵抗现象,与肥胖对照组相比,中、高剂量DNJ干预使雄鼠FGF21水平分别下降10.94%(P<0.05)、21.07%(P<0.01),且高剂量组恢复至正常水平(P>0.05),而低剂量DNJ对雄鼠FGF21的影响不显著(P>0.05)。
2.3 DNJ对肥胖小鼠血清TNF-α、IL-6、iNOS的影响
肥胖通常会导致TNF-α、IL-6、iNOS等炎性因子水平升高,使机体处于慢性低度炎症状态,进一步引发糖尿病、心血管疾病甚至癌症[24-26];另有研究表明,炎性因子水平的升高能够抑制线粒体的生物合成[27-28]。


图1 DNJ对肥胖小鼠血清炎性因子质量浓度的影响Fig. 1 Effects of DNJ on serum inflammatory factors level in obese mice
如图1 A 所示,相比正常对照组,肥胖对照组雌、雄鼠血清TNF-α水平分别上升53.03%、79.79%(P<0.01);相比肥胖对照组,低、中、高剂量DNJ使雌鼠TNF-α水平下降18.73%、18.29%(P<0.05)和29.06%(P<0.01),其中高剂量组恢复至正常水平(P>0.05),各剂量之间量效关系不显著;低、中、高剂量DNJ使雄鼠TNF-α水平下降17.90%、16.32%、30.84%(P<0.01),其中高剂量DNJ效果较好。
如图1 B 所示,与正常对照组相比,肥胖对照组雌、雄鼠血清IL-6水平分别上升40.87%、38.29%(P<0.01);与肥胖对照组相比,低、中、高剂量的DNJ使雌鼠IL-6水平分别下降11.50%、18.12%、25.72%(P<0.01),高剂量组恢复至正常组水平(P>0.05);中、高剂量DNJ使雄鼠中IL-6水平分别下降16.80%(P<0.05)、37.80%(P<0.01),具有明显的量效关系,且均恢复至正常组水平(P>0.05),但低剂量DNJ对雄鼠IL-6无显著影响。
如图1C所示,与正常对照组比较,肥胖对照组雌、雄鼠血清iNos质量浓度分别上升71.49%、61.44%(P<0.01);与肥胖对照组相比,中、高剂量DNJ使雌鼠iNOS水平分别下降21.10%、28.32%(P<0.01);低、中、高剂量D N J 能使雄鼠i N O S 水平分别下降16.08%、21.59%、29.01%(P<0.01),但均未恢复至正常水平(P<0.05)。
以上结果表明,肥胖可使机体处于炎症状态,而一定剂量DNJ的干预可以改善肥胖所导致的炎症程度。
2.4 DNJ对肥胖小鼠肝细胞线粒体合成与自噬能力的影响
肝脏脂代谢的异常涉及诸多疾病,如肥胖、非酒精性脂肪肝及糖尿病等,线粒体是重要的能量代谢场所,对于脂肪酸的代谢至关重要[29],因此,维持肝脏健康线粒体的数目对于肥胖及其并发症的改善具有重要意义。
2.4.1 DNJ对肥胖小鼠肝细胞线粒体生物合成的影响
PGC-1α是一种重要的线粒体生物合成调控因子,可促进另一种转录因子Nrf1的表达,调控线粒体DNA复制,促进线粒体生物合成[30]。
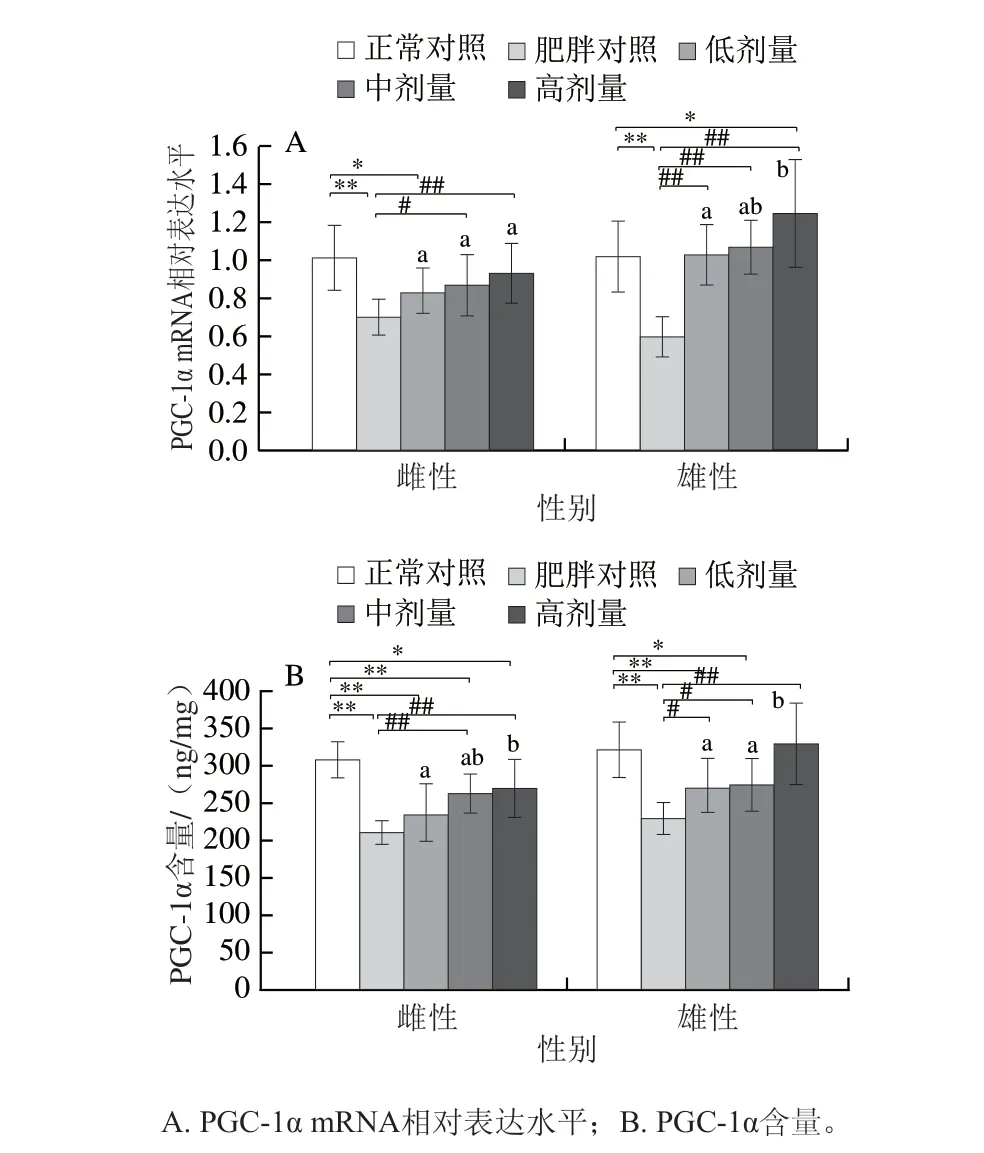
图2 DNJ对肥胖小鼠肝脏PGC-1α mRNA及蛋白表达的影响Fig. 2 Effects of DNJ on peroxisome proliferator activated receptor γ coactivator-1α mRNA and protein expression levels in liver of obese mice
如图2A、B所示,与正常对照组相比,肥胖对照组雌、雄鼠PGC-1α mRNA相对表达分别下调30.97%、41.16%(P<0.01),蛋白水平分别下降31.71%、28.77%(P<0.01)。相比肥胖对照组,中、高剂量的DNJ能够使雌鼠PGC-1α mRNA相对表达上调24.07%(P<0.05)、33.37%(P<0.01),PGC-1α蛋白水平上升24.82%、28.27%(P<0.01),低剂量组的PGC-1α mRNA相对表达及蛋白水平无显著变化;低、中、高剂量DNJ使雄鼠的PGC-1α mRNA相对表达分别上调71.95%、78.54%、108.76%(P<0.01),其中低、中剂量恢复至正常(P>0.05),高剂量组高于正常对照组(P<0.05),蛋白水平分别上升18.02%、19.80%(P<0.05)和43.99%(P<0.01),高剂量组PGC-1α蛋白水平恢复至正常水平。DNJ对雌雄小鼠血清PGC-1α mRNA相对表达与蛋白水平的变化趋势具有一致性。

图3 DNJ对肥胖小鼠肝脏Nrf1 mRNA及蛋白表达的影响Fig. 3 Effects of DNJ on Nrf1 mRNA and protein expression levels in liver of obese mice
如图3A、B所示,与正常对照组相比,肥胖对照组雌、雄鼠Nrf1 mRNA相对表达分别下调43.70%、50.18%(P<0.01),蛋白水平下降41.78%、29.49%(P<0.01)。相比肥胖对照组,低、中、高剂量DNJ使雌鼠Nrf1 mRNA相对表达分别上调45.55%、45.21%、70.00%(P<0.01),高剂量组mRNA相对表达恢复至正常水平,Nrf1蛋白水平分别上升27.42%、36.15%、50.25%(P<0.01),均未恢复至正常;中、高剂量DNJ使雄鼠Nrf1 mRNA相对表达分别上调54.13%、67.91%(P<0.01),Nrf1蛋白水平分别上升20.31%(P<0.05)、40.26%(P<0.01),低剂量组mRNA相对表达及蛋白水平均无显著变化,高剂量组Nrf1蛋白水平恢复到正常水平。DNJ对雌雄小鼠肝细胞Nrf1的mRNA相对表达与蛋白水平变化的影响具有一致性。以上结果表明,肥胖可使线粒体生物合成关键蛋白的表达下降,DNJ的干预可改善这一不利影响,促进线粒体合成,甚至可恢复到正常的合成水平。
2.4.2 DNJ对肥胖小鼠肝细胞线粒体自噬能力的影响
线粒体自噬是对衰老损伤线粒体的选择性清除,对维持正常的能量代谢、防止细胞损伤至关重要[31]。
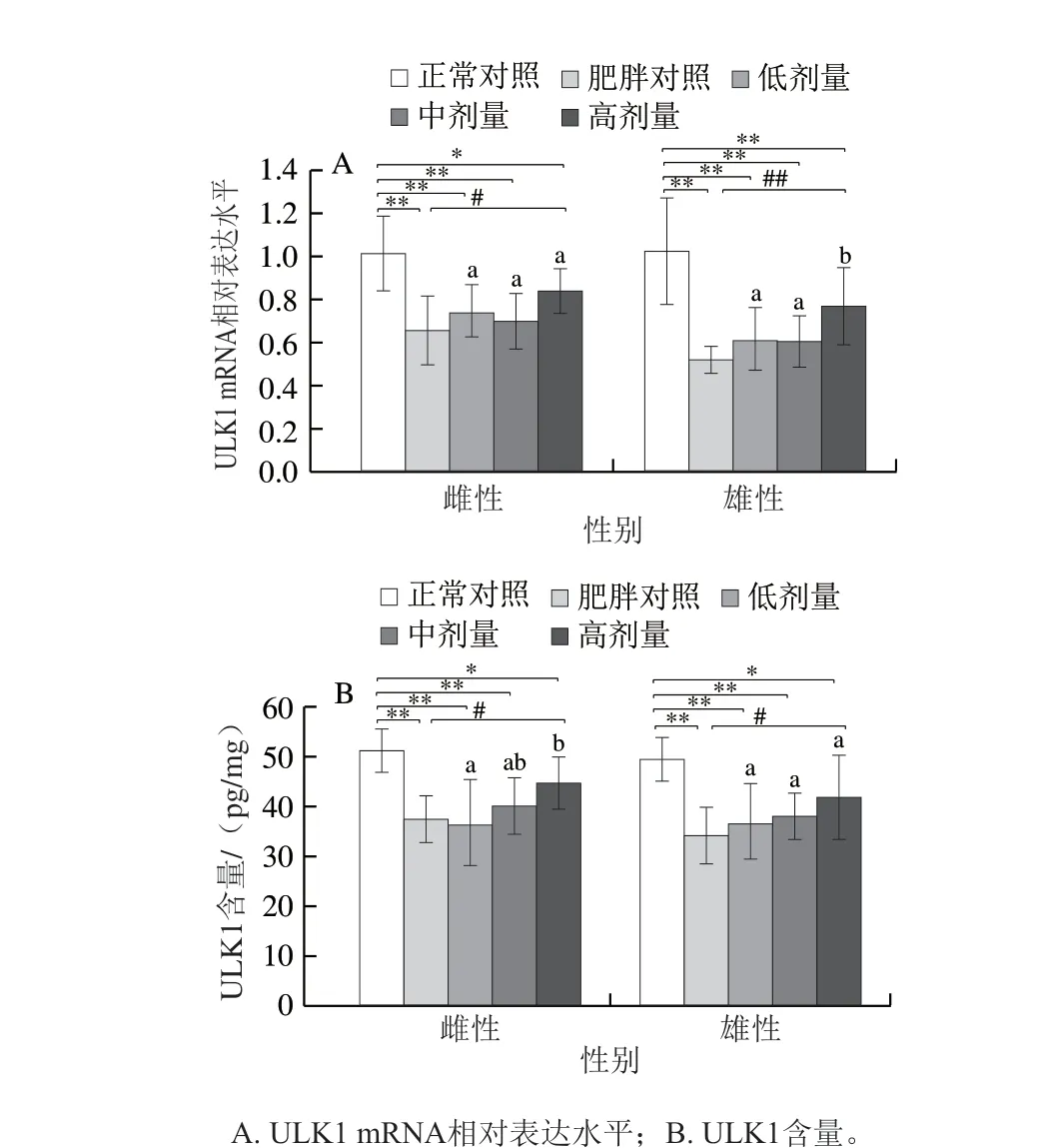
图4 DNJ对肥胖小鼠肝脏ULK1 mRNA及蛋白表达的影响Fig. 4 Effects of DNJ on unc-51 like kinase 1 mRNA and protein expression levels in liver of obese mice
如图4所示,与正常对照组相比,肥胖对照组雌、雄鼠ULK1的mRNA相对表达分别下调35.16%、49.29%(P<0.01),蛋白水平下降26.78%、30.92%(P<0.01),表明肥胖抑制了小鼠线粒体的自噬。相比肥胖对照组,高剂量DNJ能使雌鼠ULK1 mRNA相对表达上调27.71%(P<0.05),蛋白水平上升19.24%(P<0.05),低、中剂量的DNJ对雌鼠ULK1的mRNA相对表达及蛋白水平均无显著影响;高剂量DNJ能使雄鼠mRNA相对表达上调47.59%(P<0.01),蛋白水平上升22.76%(P<0.05),同样,低、中剂量的DNJ对雄鼠ULK1的mRNA相对表达及蛋白水平均无显著影响。表明较高剂量的DNJ干预对肝细胞线粒体的自噬能力有一定的恢复作用,但不能使之恢复到正常水平。
2.4.3 DNJ对肥胖小鼠肝细胞AMPK mRNA表达的影响
腺苷酸活化蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)是细胞内重要的能量传感器,涉及糖、脂代谢等诸多方面,对线粒体生物合成与自噬均具有调控作用[32],其由3 种亚基组成,其中α亚基具有两种亚型(AMPKα1与AMPKα2),起催化作用[33]。
如图5所示,与正常对照组相比,肥胖导致雌、雄鼠AMPKα1 mRNA表达分别下调48.79%、49.88%(P<0.01),AMPKα2 mRNA表达分别下调48.69%、48.23(P<0.01);相比肥胖对照组,中、高剂量的DNJ能使雌鼠AMPKα1 mRNA表达分别上调37.04%、60.33%(P<0.01),使雄鼠AMPKα1 mRNA分别上调49.04%、50.68%(P<0.01);但各剂量DNJ均未对雌、雄鼠AMPKα2 mRNA表达产生显著影响。
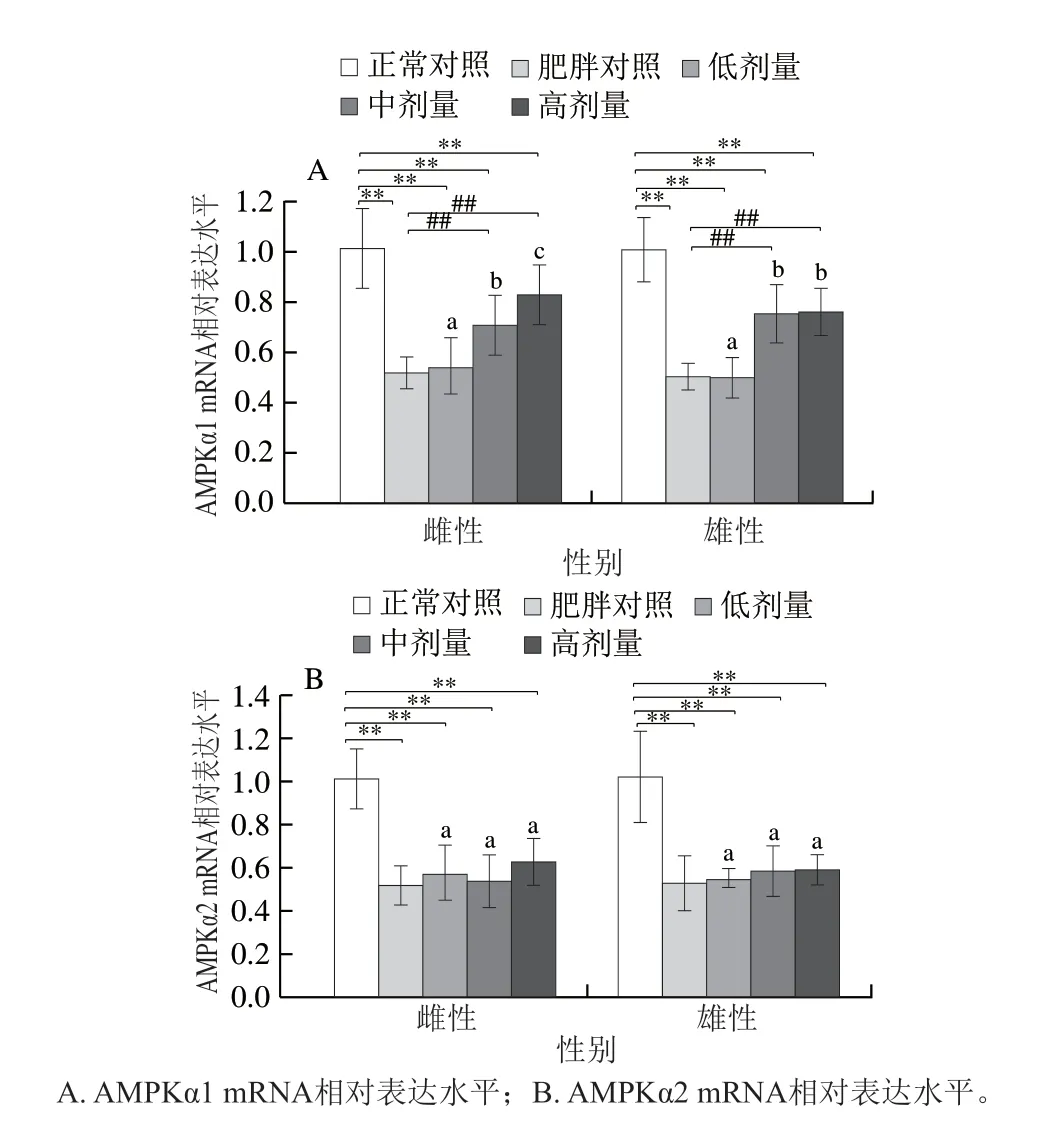
图5 DNJ对肥胖小鼠肝脏AMPKα1及AMPKα2 mRNA表达的影响Fig. 5 Effects of DNJ on AMPKα1 and AMPKα2 mRNA expression in liver of obese mice
2.5 DNJ对肥胖小鼠肝细胞CPT1活性的影响
CPT1是脂肪酸β氧化过程中的限速酶,位于线粒体外膜,其主要作用是转运脂酰CoA进入线粒体[34]。

图6 DNJ对肥胖小鼠肝脏CPT1活力的影响Fig. 6 Effect of DNJ on carnitine palmitoyltransferase 1 activity in liver of obese mice
如图6所示,与正常对照组相比,肥胖对照组雌、雄鼠CPT1活力分别下降44.76%、35.52%(P<0.01),表明肥胖个体脂肪酸分解代谢下降。相比肥胖对照组,中、高剂量的DNJ使雌鼠CPT1活力分别上升49.77%、58.05%(P<0.01),使雄鼠的CPT1活力上升21.39%(P<0.05)、49.60%(P<0.01),其中高剂量组雄鼠CPT1活力恢复至正常水平(P>0.05)。以上结果表明,一定剂量的DNJ干预能够促进肥胖小鼠脂肪酸分解,达到降脂的目的。
3 讨 论
肥胖是一种能量失衡导致的慢性疾病,同时也是心血管疾病、2型糖尿病、癌症等疾病的主要风险因素[35]。已有大量研究证实了DNJ在促进脂代谢、缓解肥胖方面的作用,如李有贵等[36]以50 mg/kg mb剂量桑叶DNJ灌胃高脂血症模型小鼠90 d,结果表明小鼠血清TC、TG水平逐渐恢复至正常水平。Tsuduki等[37]以高脂饲料饲喂大鼠,同时以1 mg/kg mb剂量灌胃DNJ 4 周,结果表明DNJ能够通过激活脂肪酸β氧化促进脂代谢,降低机体脂肪积蓄。在后续的研究中,Tsuduki等[38]通过进一步研究发现DNJ通过改善肥胖小鼠脂联素水平达到促进脂代谢的目的。本研究同样显示DNJ对肥胖小鼠脂联素具有改善作用,考虑到FGF21-脂联素轴在脂代谢中发挥重要作用,FGF21能够促进脂联素合成及分泌[39],因此对FGF21水平进行测定但在不同性别小鼠中观察到不同的结果,在雌性肥胖对照组小鼠中表现为FGF21水平下降,可能是合成及分泌不足导致,而在雄性肥胖对照组小鼠中则表现为FGF21水平显著升高,这与Chukijrungroat等[40]研究结果相一致,推测为FGF21抵抗现象,DNJ干预后,雌鼠FGF21合成分泌增加,雄鼠FGF21抵抗现象得到改善,造成这种性别差异的具体原因有待进一步研究。AMPK是体内重要的能量传感器,对能量代谢的平衡至关重要,FGF21与脂联素均可通过AMPK途径调控PGC-1α表达,促进线粒体合成[41-42]。肥胖个体线粒体通过能量代谢产生大量的ROS会对线粒体本身造成损伤,导致线粒体功能紊乱,对这些损伤线粒体的清除是十分有必要的,AMPK除促进线粒体合成外,还可通过磷酸化激活ULK1,促进损伤线粒体自噬,维持健康线粒体数量[43-44]。在本研究中,肥胖对照组PGC-1α、Nrf1以及ULK1表达显著下降,45 d的DNJ干预可以改善这一状况同时上调AMPKα1表达,表明DNJ可能通过改善FGF21与脂联素水平调节AMPK,进一步实现对肥胖小鼠线粒体生物合成与自噬的改善作用,但DNJ对AMPKα2的表达却未有显著影响。AMPK可磷酸化脂肪酸合成限速酶乙酰辅酶A羧化酶1(acetyl-CoA carboxylase 1,ACC1),使之活性下降,抑制脂肪酸合成,同时还可通过磷酸化ACC2提高CPT1活性,促进脂肪酸氧化[45],本研究的结果表明,肥胖小鼠经DNJ灌胃后,CPT1活性上升,可能与DNJ上调AMPKα1表达有关。
肥胖通常伴随着TNF-α、IL-6、iNOS等炎性因子水平显著升高,使机体处于慢性低度炎症状态,慢性低度炎症与胰岛素抵抗、糖尿病、心血管疾病的发展密切相关[46],还有研究发现这些炎性因子不仅会下调PGC-1α表达,抑制线粒体合成[27-28],还会导致FGF21活性功能下降,抑制脂联素分泌[47-48],在本研究中,肥胖对照组血清TNF-α、IL-6、iNOS水平显著升高,经过DNJ干预后,炎性因子水平均得到不同程度改善,与Zheng Junping等[49]研究结果相一致,表明DNJ具有抗炎作用。
本实验同时观察到DNJ对不同性别肥胖小鼠的改善作用具有性别差异。对于线粒体生物合成的转录因子PGC-1α和Nrf1,经高剂量DNJ灌胃45 d后,雄鼠PGC-1α和Nrf1蛋白水平均已恢复至正常水平,同剂量灌胃的雌鼠却仍与正常对照组存在显著差异,暗示DNJ对雄性肥胖小鼠线粒体合成的促进作用优于雌鼠;在对肥胖小鼠炎症的改善作用中,中剂量的DNJ便使雄鼠IL-6恢复至正常水平,雌鼠则需要高剂量,另一方面,高剂量DNJ使雌鼠TNF-α水平恢复至正常,而雄鼠则仍显著高于正常对照组。造成这些差异的原因有待进一步研究,这些差异也暗示了当使用DNJ时,应考虑性别差异予以剂量上的调整。此外,DNJ对线粒体合成与自噬的其他影响途径还需要更深入的研究。
DNJ可能通过降低肥胖小鼠炎症程度,改善FGF21与脂联素水平,进一步上调AMPKα1表达,促进肥胖小鼠肝脏线粒体生物合成与自噬,同时使线粒体脂肪酸氧化限速酶CPT1活性上升,促进脂肪酸分解,达到降脂的目的。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
做人与处世(2020年10期)2020-06-29
科技创新与品牌(2018年5期)2018-07-24
安徽医科大学学报(2015年9期)2015-12-16
医学研究杂志(2015年6期)2015-07-01
郑州大学学报(医学版)(2015年2期)2015-02-27
中国药业(2014年24期)2014-05-26
中国药业(2014年21期)2014-05-26
