
生命早期环境不可预测性对过度进食的影响:基于生命史理论*
2020-10-20 08:32罗一君牛更枫
心理学报 2020年10期
罗一君 牛更枫 陈 红
(1 西南大学心理学部;2 西南大学认知与人格教育部重点实验室,重庆 400715) (3 华中师范大学心理学院,武汉 430079)
1 引言
生命史理论(Life History Theory)认为,在不可预测的生存环境中,疾病或暴力可能使得长期投入付之一炬,个体无法从长期投入中获得回报,因而会更倾向于享受当下追求即时满足(Mittal &Griskevicius,2014)。食物可以被视为即时奖励,一方面它可以满足基本能量需求,另一方面食物可以帮助转移注意力并改善情绪。不仅如此,由于食物与香烟酒精等成瘾物质有着相似的多巴胺反应系统(Nieto,Wilson,Cupo,Roques,&Noble,2002),并能激活相似的奖赏脑区(Volkow,Wang,Fowler,Tomasi,&Baler,2011),有研究者甚至将食物视为成瘾物质(Ziauddeen &Fletcher,2013)。因此在面对美味食物刺激时,环境不可预测性高的个体会更倾向于放任进食欲望而忽视长远的健康风险。
过度进食(Overeating)是指个体摄入食物超过自身能量需求的一种进食行为(Prentice,2001),它不仅是肥胖和进食障碍的直接原因,而且也是抑郁、焦虑、自伤和物质滥用等内化和外化问题的重要风险因素(Striegel-Moore &Bulik,2007)。同时,过度进食还有着较高的发生率——在美国49%女性和 30%男性报告有过度进食和暴饮暴食行为(Lipson &Sonneville,2017);在中国,过度进食数量和比例都呈逐年上升的趋势(Tong et al.,2014)。基于这一背景,过度进食的影响因素得到了研究者的广泛关注。大量研究表明,生命早期生活在不可预测环境中的个体更可能出现过度进食行为(Miller et al.,2018;Maner,Dittmann,Meltzer,&McNulty,2017;Proffitt Leyva &Hill,2018),但其内部作用机制尚不清晰。有研究者指出,在生命史理论背景下研究相关行为时,应综合考虑外部环境不可预测性与内部身体状态的交互效应(Chang et al.,2019a)。Chang 等(2019a)发现,身体状况较差时,环境不可预测性对个体攻击行为、冒险行为和冲动性的预测效应比身体状况较好时更大。据此可以推断,内部状态较差(例如饥饿状态)时,个体可能有更大的即时满足倾向和不健康进食行为。而实验研究则发现,个体处于饱食状态时,压力组个体有更多过度进食行为(Hill,Prokosch,DelPriore,Griskevicius,&Kramer,2016)。本研究拟从生命史视角出发,探究生命早期环境不可预测性影响过度进食的机制以及饥饿状态的作用。
1.1 生命早期环境不可预测性与生命史策略
生命史理论认为在分配有限资源的过程中,个体必须在躯体投入(Somatic effort,指个体在维持和发展自身资本时的投入,如学习知识和技能)和繁衍投入(Reproductive effort,指个体在那些可以获得即时满足活动上的投入,如睡眠和进食)之间做出适应于当前环境的权衡决策(Mittal &Griskevicius,2014),而分配和权衡资源时的倾向被称为生命史策略。快生命史策略(Fast life history strategies)指有机体牺牲长期利益而着眼于当下即时满足的行为倾向,而慢生命史策略(Slow life history strategies)指牺牲眼前获得投入到长远利益的行为倾向。
生命史策略会受到生命早期环境的影响。研究发现,环境不可预测性(Environmental unpredictability)会促使个体采取快生命史策略(Ellis,Figueredo,Brumbach,&Schlomer,2009)。因为在这类环境中,危险和暴力的不可预测性可能让个体过早地失去生命,长期投入可能因为疾病或暴力付之一炬,牺牲眼前获得而关注长期投入的行为并不能有效利用资源;相反地,将未来收益折现到当前的行为倾向可以让此类环境中的个体将资源最大化利用。在人类社会中,环境不可预测性主要表现为居住环境和养育环境的不可预测性(Proffitt Leyva &Hill,2018)。研究者指出,生命早期环境不可预测性较高,例如,危险暴力的居住环境,父母的奖惩机制不统一,情绪不稳定等,都代表着威胁以及可获得的物质资源和情感资源的不可预测(Lu &Chang,2019)。这种居住和养育环境的不可预测性会使个体发展出较快的生命史策略。如Chang 等(2019b) 整合居住和养育环境的不可预测性以及家庭收入作为环境压力的指标,以来自不同文化背景下的1245 名青少年为被试,结果发现生命早期环境压力能够显著正向预测5 年半后的快生命史策略;Lu 和Chang(2019)招募了中国留守儿童和对照组,整合父母缺失、家庭环境混乱、负性生活事件和感知压力作为环境威胁和不可预测性的指标,结果发现它能显著正向预测18 个月后的快生命史策略。此外,回溯性研究也发现,个体回忆的生命早期环境不可预测经历越多,其在快生命史策略上的得分越高(Maner et al.,2017)。因此,生命早期环境不可预测性可能是快生命史策略的重要影响因素。
1.2 生命史策略与过度进食
由于快生命史策略是一种享受当下而忽视未来风险的认知和行为倾向。在食物资源有限且不可预测的环境中,当前过度进食以增加能量和脂肪储备的快策略倾向有助于应对食物匮乏的时期,使个体得以生存和繁衍(Ahlstrom,Dinh,Haselton,&Tomiyama,2017)。尽管在现代社会,食物匮乏很少出现,但由于快生命史策略与冲动性和即时满足高相关,而美味食物可以满足个体即时享乐的需求,因此拥有快生命史策略个体仍可能有着过度进食的倾向。相关横断和纵向研究都发现,快生命史策略不仅与个体的攻击行为以及吸烟酗酒等损害长期健康的问题行为正相关(Chang et al.,2019b;Lu&Chang,2019),还与问题性进食行为显著正相关(Maner et al.,2017;Salmon,Figueredo,&Woodburn,2009)。Maner 等(2017)通过对400 名成年被试的在线调查发现,快生命史策略与过度进食行为、身体质量指数显著正相关。Salmon 等(2009)发现快生命史策略能显著正向预测进食障碍症状。此外,纵向研究提示生命史策略可以作为生命早期环境不可预测性和一系列快策略相关问题行为(如攻击行为)的中介变量(Chang et al.,2019a,2019b;Lu &Chang,2019)。进食领域的研究也显示快生命史策略在生命早期环境不可预测性与过度进食中起中介作用(Maner et al.,2017)。基于上述证据,本研究假设快生命史策略能在生命早期环境不可预测性对过度进食的影响中起中介作用。
1.3 死亡威胁的作用
生命史策略对行为的影响也会受到当前环境的影响。研究者指出,个体行为不仅受根植于生命早期所形成的固定反应策略的影响,也会随着当前情境的改变而变化——当前不可预测的情境会激活并扩大个体在生命早期形成的针对不可预测情境的固定反应图式(即快生命史策略) (Proffitt Leyva &Hill,2018)。在不可预测的情境中,死亡威胁的不可预测性对个体行为的预测效力最强(Mittal&Griskevicius,2014)。在死亡威胁启动下,个体更倾向于表现出与快策略相关的反应倾向,例如高冲动性和高冒险性行为,并倾向于获得即时奖励(Griskevicius,Delton,Robertson,&Tybur,2011)。有研究发现,在死亡威胁启动时,童年压力组对异性身体吸引力的要求显著低于控制组(汪佳瑛,陈斌斌,2016),而身体吸引力通常是好基因的体现,该研究表明死亡威胁启动条件下与快策略相关的行为倾向——繁衍动机——被加强。就进食行为而言,在死亡威胁启动下,童年压力较高个体会更多地过度进食(Hill et al.,2016)。研究者指出,在死亡威胁情境下,快策略个体倾向于认为威胁是不可控的,因而会折现未来以保证当下的享乐行为(Mittal &Griskevicius,2014)。因此,感知死亡威胁可能会增强快生命史策略对过度进食的影响。此外,以往研究对于死亡威胁的高低区分多为实验室操纵,缺乏生态效度,本研究拟选取新冠病毒疾病爆发背景下高死亡威胁vs.控制组个体,探究感知死亡威胁在快生命史策略与过度进食关系中的调节作用。
1.4 社会支持的作用
即便个体行为很大程度受进化规律的影响,但作为社会性较高的人类依然有着其独特的适应方式。社会支持是指个体从家庭,朋友,同事或团体组织等社会关系中获得的物质和精神支持(姚若松,郭梦诗,叶浩生,2018)。社会支持会提高个体对自身的总体评价,使个体相信自己被关心和接受,从而减轻个体当前的压力并增加个体积极行为出现的频率(Dukes &Holahan,2003),进而促进个体的良好适应。国内外研究者一致认为,社会关系建构了情绪调节的外部资源,可以弱化或增强情绪调节对心理适应和行为的影响路径(Berkman &Syme,1979;Marroquín &Nolen-Hoeksema,2015;周宵,伍新春,曾旻,田雨馨,2016)。就进食行为而言,研究表明作为社会支持的一部分,良好友谊质量会促进积极身体意象的形成,进而增加积极的进食行为(陈曦梅,罗一君,陈红,2020b)。如前所述,快生命史策略是由环境不可预测性造成的一种将当前收益最大化不考虑未来风险的行为倾向,而社会支持往往伴随着信息支持和物质支持(Malecki &Demaray,2003),这在一定程度上减轻了当前环境带来的压力,进而缓解快生命史策略对过度进食的影响;另一方面,社会支持伴随的情感支持可以促使个体以怜悯和尊重的态度对待自身,正视身体的能量需求,进而使个体根据内部的饥饿感和生理线索来进食(陈曦梅 等,2020b),这在一定程度上也可以缓解快生命史策略对过度进食的影响。因此,社会支持可能作为快生命史策略影响过度进食的保护性因素。
此外,由于少有研究直接选取处于生命早期的个体探究生命早期环境不可预测性对过度进食的影响,也缺乏外部环境威胁和内部状态共同影响过度进食的证据。基于以上论述,本研究拟通过两个研究来系统探讨生命早期环境不可预测性对过度进食的影响及其内在机制。具体而言,研究1 选取正处于生命早期的青少年群体,操纵饥饿状态(饥饿vs.饱食),并假设生命早期环境不可预测性能预测个体的过度进食。在饱食状态下,如果个体选择的食物份量和实际进食量越多,则表示有越多的过度进食行为。此外,少有研究从生命史理论的视角解释生命早期环境不可预测性与过度进食行为的关系机制,并探究其中的威胁性因素和保护性因素,同时以往研究对于死亡威胁的操纵缺乏生态效度。因此,研究2 选取新冠病毒疾病爆发背景下,死亡威胁较高的武汉市社区居民和死亡威胁相对较低的其他市居民,采用问卷法令被试回忆生命早期(5~14 岁时)环境不可预测性,并探究其影响当前过度进食的机制。研究2 假设,生命早期环境不可预测性通过生命史策略的中介作用间接影响过度进食,且死亡威胁较高时,快生命史策略个体会有更多的过度进食行为,而在社会支持较高时,快生命史策略个体则不会有更多的过度进食行为,即死亡威胁会扩大环境不可预测性通过生命史策略间接影响过度进食的效应,而社会支持则能缓冲这一效应。此外,有研究发现环境恶劣性对个体进食行为有重要影响(Cheon &Hong,2017),因此,在两个研究中,环境恶劣性均作为协变量进行控制。
2 研究1:环境不可预测性对青少年过度进食的影响
2.1 被试
获得西南大学心理学部伦理委员会审批和同意后,研究者联系了武汉市一所公立初中学校,取得4 个班级的班主任同意后,在课余时间招募自愿参与者参与研究。经Gpower 计算,本研究需要招募n
≥ 89 的被试量以达到足够的统计检验力(Faul,Erdfelder,Lang,&Buchner,2007)。最后,研究招募了处于生命早期的初一和初二年级学生被试(n
=91,男生=48),年龄分布为12~14 岁(M
=13.59,SD
=0.61),身体质量指数分布为12.94~29.38 (M
=20.10,SD
=3.32)。2.2 研究过程
本研究采用饱食进食范式(Eating in the absence of hunger) (Hill et al.,2016;Cheon &Hong,2017),操纵了饥饿状态(饥饿vs.饱食),旨在探讨在饱食状态下,高环境不可预测性个体是否有更多的进食行为(即,过度进食)。主要程序如下:(1)实验要求被试在午餐时间无进食来到实验室,完成饥饿状态和情绪状态测量,随后被随机分配到饱食组和饥饿组;(2)在饱食条件下,被试(n
=43,男生=17)获得一份标准的中餐饭盒,其中包括米饭150 g、蔬菜100 g、鸡肉50 g 以及汤200 g,并被告知可以随意进食直到吃饱;在饥饿条件下,被试(n
=48,男生=31)获得一杯水,被告知可以随意饮取;(3)被试报告自己的饥饿状态作为操纵检验的指标;(4)被试在电脑上完成食物份量选择任务;(5)被试被告知主试需要两分钟时间准备材料,在等待时间里被试可以随意进食桌上两个碗里的零食(巧克力和葡萄干);(6)两分钟后,主试收走零食并计算被试进食的重量(g),实验结束。最后,饥饿组被试也获得一份标准饭盒,所有被试完成问卷测量后获得一只定制水杯。本研究中,自变量为环境不可预测性,调节变量为饥饿状态,因变量为被试选择的食物份量(包括高热量食物份量和低热量食物份量)以及实际进食量。2.3 研究材料
2.3.1 食物份量选择任务
本研究采用食物份量选择任务。在每个单独的trial,要求被试根据自己当前的状态选择想要进食的食物份量。指导语为:下面每一幕都会出现一种食物,请根据你当前的真实感受选择你想吃的食物份量。选择不吃请按0,选择一份请按1,选择两份请按2,选择三份请按3,选择四份请按4 (图1)。
2.3.2 高热量与低热量食物图片
实验材料制作方式参照以往研究(Sim,Lim,Forde,&Cheon,2018)。首先,要求30 名青少年(年龄=13.30 ±0.47 岁,男=11)对20 张高热量食物图片和20 张低热量食物图片的唤醒度、熟悉度、愉悦度、美味度、饱腹程度、健康度和热量高低进行1~7 点评分。由于低熟悉度,两张高热量和两张低热量图片被移除。正式实验材料保留18 张高热量与18 张低热量图片,两类图片在唤醒度和熟悉度上无显著差异,但高热量食物图片在愉悦度、美味度、饱腹程度和热量上显著高于低热量食物图片,而低热量食物图片在健康度上显著高于高热量食物图片。最后每种食物图片都用Photoshop 软件被做成0~4 份放在同一张图上。

图1 食物份量选择任务
2.3.3 研究工具
生命早期环境不可预测性 采用修订的环境不可预测性测量问卷,包含两个维度:居住环境不可预测性(3 题,例,我的家里总是杂乱无章) (Mittal,Griskevicius,Simpson,Sung,&Young,2015),以及父母行为和情绪不可预测性(6 题,例,父母在特定情境里的行为取决于他/她的情绪) (Ross &Hill,2000)。所有项目经正向计分,得分越高环境不可预测性程度越大。该量表在本研究有良好的结构效度,χ(25)=1.28,RMSEA=0.055,CFI=0.967,TLI=0.953,SRMR=0.056;内部一致性系数居住环境不可预测性α=0.72,父母行为和情绪不可预测性α=0.83,总内部一致性系数为α=0.81.
生命早期环境恶劣性 采用汪佳瑛等(2016)所使用的社会经济地位量表进行测量(例,我的家庭有足够多的钱来满足各种需求)。所有项目经反向计分,得分越高环境恶劣性程度越大。内部一致性系数α=0.76.
饥饿状态和情绪状态 饥饿状态采用一个题项测量“你现在饥饿的程度是多少?”被试在1~7 点量表(1=一点也不饿,7=非常饿)上选择符合自己真实感受的数字;情绪状态采用0~100 点VAS 量表测量10 种(抑郁的、满意的、难过的、恼怒的、快乐的、羞耻的、害怕的、轻松的、有压力的、内疚的)情绪状态。
过度进食度指标 本研究选用3 个指标作为过度进食度的指标,分别是饱食组被试选择的高热量食物份量、低热量食物份量和实际进食量(g) (Hill et al.,2016)。
2.4 结果
采用SPSS 20.0 进行数据分析。首先,采用多元回归分析检验饥饿组和饱食组的组间差异,发现两组在年龄,BMI 和情绪状态上组间差异均不显著(p
s >0.072);同时,饱食组(M
=4.23,SD
=1.34)与饥饿组(M
=3.93,SD
=1.33)的饥饿状态在基线测量时无显著差异,F
(1,89)=1.13,p
=0.290,但饱食组(M
=1.86,SD
=0.86)的饥饿状态得分在后测显著低于饥饿组(M
=4.48,SD
=0.74),F
(1,89)=242.26,p
<0.001,表明饥饿状态的操纵是有效的;其次,采用调节回归分析,在控制了年龄,性别,身体质量指数和环境恶劣性后,对环境不可预测性与饥饿状态在高热量食物份量、低热量食物份量和实际进食量三个变量上的交互作用进行分析。结果发现(见表1)。1)饥饿状态可以调节环境不可预测性对低热量食物份量选择(β
=0.89,SE
=0.03,t
=2.65,p
=0.010)和高热量食物份量选择(β
=0.73,SE
=0.03,t
=2.20,p
=0.030),而无法调节环境不可预测性对实际进食量的影响(β
=0.38,SE
=0.51,t
=1.14,p
=0.257)。简单效应分析显示,(a)在因变量为低热量食物份量时,对于饥饿组,高环境不可预测性个体选择了更少的低热量食物(β
=-0.39,SE
=0.02,t
=-2.86,p
=0.006),而对于饱食组,环境不可预测性无法预测低热量食物选择(β
=0.14,SE
=0.02,t
=0.90,p
=0.375) (图2a);(b)在因变量为高热量食物份量时,对于饥饿组,环境不可预测性无法预测高热量食物选择(β
=-0.12,SE
=0.02,t
=-0.79,p
=0.432),而对于饱食组,高环境不可预测性个体选择了更多的高热量食物(β
=0.38,SE
=0.02,t
=2.61,p
=0.013) (图2b)。
表1 饥饿状态(饥饿vs.饱食)对生命早期环境不可预测性与过度进食的调节作用

图2 简单效应分析:(a)饥饿状态对环境不可预测性影响低热量食物份量选择的调节作用;(b)饥饿状态对环境不可预测性影响高热量食物份量选择的调节作用;EU 指生命早期环境不可预测性;低EU=环境不可预测性得分为负一个标准差(-1 SD),高EU=环境不可预测性得分为正一个标准差(+1 SD)
2.5 讨论
研究1 选取了处于生命早期的青少年群体,探究了环境不可预测性与过度进食的关系,发现在饱食状态下,环境不可预测性越高,个体会选择越大份量的高热量食物,即有更大的过度进食行为倾向。这一结果与回溯性的研究证据相符(Hill et al.,2016;Proffitt Letva et al.,2018),他们认为个体在饱食状态下也会选择过度进食是因为生命早期环境不可预测性使个体形成了不可预测的认知图式——环境不可控且养育者不可靠,因而发展出即时享乐的行为倾向。因此,当个体面对高热量/不健康食物时,即便在饱食状态下,环境不可预测性较高的个体也倾向于忽视长远健康风险(肥胖及相关疾病等)而选择更大的食物份量。本研究一方面进一步验证了以往研究结果。另一方面,本研究还发现在饥饿状态时,环境不可预测性越高,个体选择的低热量/健康食物越少。进食的营养均衡是保持健康的重要条件之一,更少的低热量/健康食物意味着有更高的健康风险。这一结果在一定程度上符合Chang 等(2019a)的发现——外部环境威胁在个体状态较差时对问题行为影响更大。由于饥饿状态属于一种需求状态,在饥饿状态下,环境不可预测性较高个体的高冲动性和低自我控制性令其忽视健康饮食而选择了更少的低热量/健康食物。这一新发现为生命早期高环境不可预测性个体有着更大的健康风险的观点提供了证据。
但本研究并未发现饥饿状态与环境不可预测性对实际进食量影响的交互作用,可能的原因是研究中食物种类有限,被试本身对食物的喜好影响了研究结果(Hill et al.,2016)。Shapiro 等(2019)采用的饱食进食范式是在被试进食标准餐后,再提供较多的食物种类选择(自助餐厅进食)。未来研究可参考这一方式,扩大食物种类的选择以增加研究测量效度和生态效度。本研究表明环境不可预测性可以直接影响过度进食,后续研究还需要在这一基础上探究中介(如何影响)和调节(何时作用会放大;何时作用会缓冲)因素。
3 研究2:生命早期环境不可预测性对成年人过度进食的影响
已有研究提示,生命早期环境不可预测性对过度进食的影响是通过生命史策略的中介作用实现的。但上述研究并未直接测量生命史策略,也没有探索这一路径中可能的当前环境因素和保护性因素。在研究1 的基础上,研究2 进一步选取不同死亡威胁环境下的个体(高死亡威胁vs.控制组),旨在探索生命早期环境不可预测性与过度进食关系中的情境因素。同时,基于前言对于生命史策略和社会支持的综述,本研究同时探究生命史策略在环境不可预测性与过度进食关系中的中介作用和社会支持在这一中介关系中的调节作用。
3.1 被试
自新型冠状病毒疾病(Coronavirus Disease-19,COVID-19)爆发以来,截止到2020 年1 月30 日已经有数以千计的人们感染,这会诱发较高的死亡威胁。本研究设计在获得西南大学心理学部伦理委员会审批并同意后,于2020 年1 月23 日~2020 年2月4 日武汉市封城后的14 天内招募武汉市居民作为高死亡威胁组,同时招募湖北省外42 个受疫情影响相对较小城市的居民(北京,唐山,重庆,济南,贵阳等)作为控制组。由主试通过问卷填写者的网络IP 地址确认每位被试的当前所在地。被试统一在网络上阅读并同意知情同意书后,完成生命早期环境不可预测性、感知死亡威胁、消极情绪、生命史策略、社会支持和过度进食问卷测量。每位参与者通过支付宝转账获得5 元人民币报酬。同时,对于消极情绪得分较高者,为其推荐西南大学心理学部的免费心理援助服务。此外,为防止被试不认真填写,在题目中穿插测谎题项,询问其所留的支付宝倒数第四位是奇数或偶数,在数据分析时进行剔除。最终剔除了23 个测谎题项错误的数据后,保留了301 名武汉市居民数据作为高死亡威胁组,179名湖北省外42 个城市的居民数据作为控制组。经Gpower 计算,本研究需要招募n≥89 的被试量以达到足够的统计检验力(Faulet al.,2007),被试量符合要求。被试的年龄分布为在18 到60 岁之间(M
=27.24,SD
=9.77)。3.2 测量工具
3.2.1 感知死亡威胁
采用汪佳瑛和陈斌斌(2016)用以检验感知死亡威胁的4 道题项:(1)你感觉环境变得更加危险的程度是多少?(2)你感觉环境变得不安全的程度是多少?(3)你感觉环境变得更加不可预测的程度是多少?(4)你感觉环境变得更加不确定的程度是多少?所有问题都使用1 (非常不强烈)到7 (非常强烈)的7 点量表进行评分。在本研究中,内部一致性系数α
=0.94。3.2.2 生命早期环境不可预测性
测量项目与研究1 相同。但在指导语上限定被试回忆在生命早期(5~14 岁)时居住环境和父母情绪与行为不可预测性的问题。该量表在本研究有良好的结构效度,χ(25)=2.49,RMSEA=0.056,CFI=0.982,TLI=0.971,SRMR=0.030.内部一致性系数父母情绪与行为不可预测性α
=0.90,居住环境不可预测性α=
0.63,
总内部一致性系数为α=
0.89.
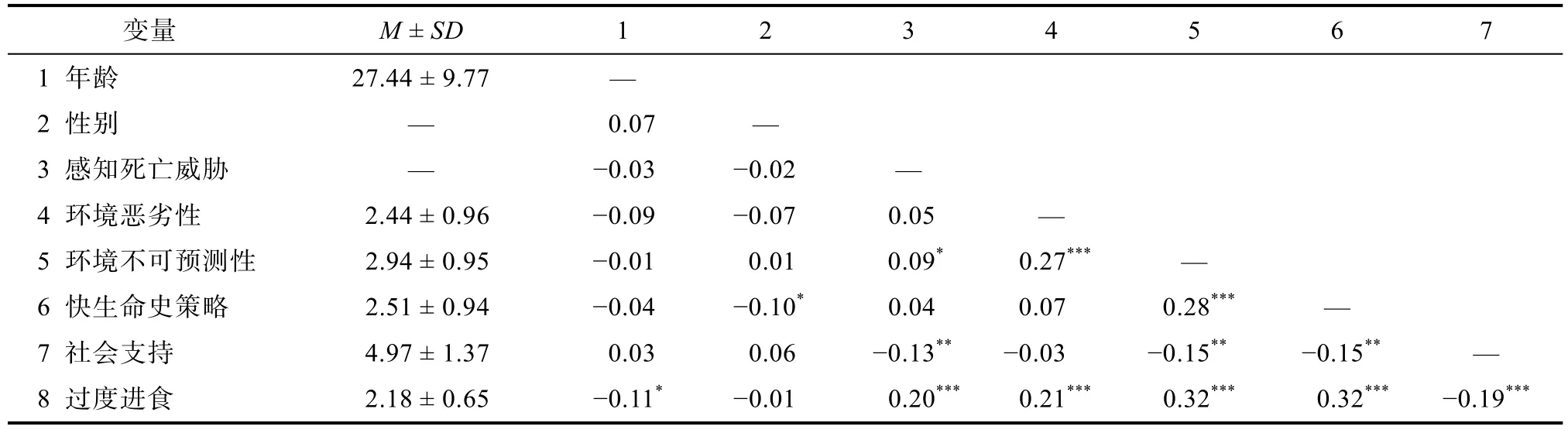
表2 描述性统计与相关分析(n=480)
3.2.3 生命早期环境恶劣性
从工业内部结构情况看,我市仍以重化工业为主,其发展演变相对滞后。2012年,工业行业大类处于产值前6名的分别是:化学原料和化学制品制造业、石油加工、石油和天然气开采、橡胶和塑料制品业、有色金属冶炼和压延加工业、纺织业。而各种金属制品和机械加工业的产值之和约1330亿元,只占到化学工业的55%。从工业化发展阶段的一般特征来看,我市已经进入到工业化的后期(第4阶段),现在仍以重化工业为主的产业结构,与以加工组装工业为主导产业的标准相比相对滞后,需要采取措施促进重工业向高加工度化和技术集约化转变。
测量项目与研究1 相同。但在指导语上限定被试回忆在生命早期(5~14 岁)时社会经济地位的问题。在本研究中内部一致性系数α
=0.86。3.2.4 生命史策略
采用Mini-K 量表(Figueredo et al.,2006)。原量表是根据20 项心理行为指标来测量个体在快慢策略连续谱系的相对位置(例,我常常能预知事情的结果),所有问题都使用1 (非常不符合)到5 (非常符合)的5 点量表进行评分。由于本研究的主要假设,本研究将所有题项反向计分后相加作为快策略的指标,得分越高表明策略越快。该量表在中国群体中有良好的应用(王燕,林镇超,侯博文,孙时进,2017)。本研究中内部一致性系数α
=0.91。3.2.5 社会支持
采用领悟社会支持量表(汪向东,王希林,马弘,1999,心理卫生评定测量手册,pp.131~132)。量表通过12 个项目来测量个体感知的社会支持[例,在我遇到问题时,有些人(亲戚,朋友,同学)会出现在我身旁]。所有问题都使用1 (非常不同意)到7(非常同意)的7 点量表进行评分。得分越高,表明社会支持越高。本研究中内部一致性系数α
=0.95。3.2.6 过度进食
采用Anglé 等(2009)三因素进食量表中的失控进食分量表进行测量。该分量表有9 个题目(例,即使刚吃完饭,但当我闻到或看到美味的食物时,我很难忍住不吃),所有问题限定的时间都为新冠病毒疾病爆发以来,被试在1 (非常不符合)到4 (非常符合)的4 点量表上进行基于真实感受和情况的评分。该量表在中国群体的研究中具有良好的信效度和适用性(Chen,Luo &Chen,2020a).在本研究中,内部一致性系数α
=0.92。3.3 结果
采用SPSS 20.0 进行描述性数据分析。
1)采用t
检验分析高死亡威胁组(M
=21.67,SD
=4.34)和控制组(M
=18.64,SD
=6.82)在感知死亡威胁得分的组间差异,两组在感知死亡威胁得分上差异显著,t
(1,478)=35.32,p
<0.001,d
=0.53。2)采用皮尔逊积差相关分析变量间的两两相关关系,发现生命早期环境不可预测性与快生命史策略和过度进食正相关;感知死亡威胁与过度进食正相关,而社会支持与过度进食负相关(见表2)。
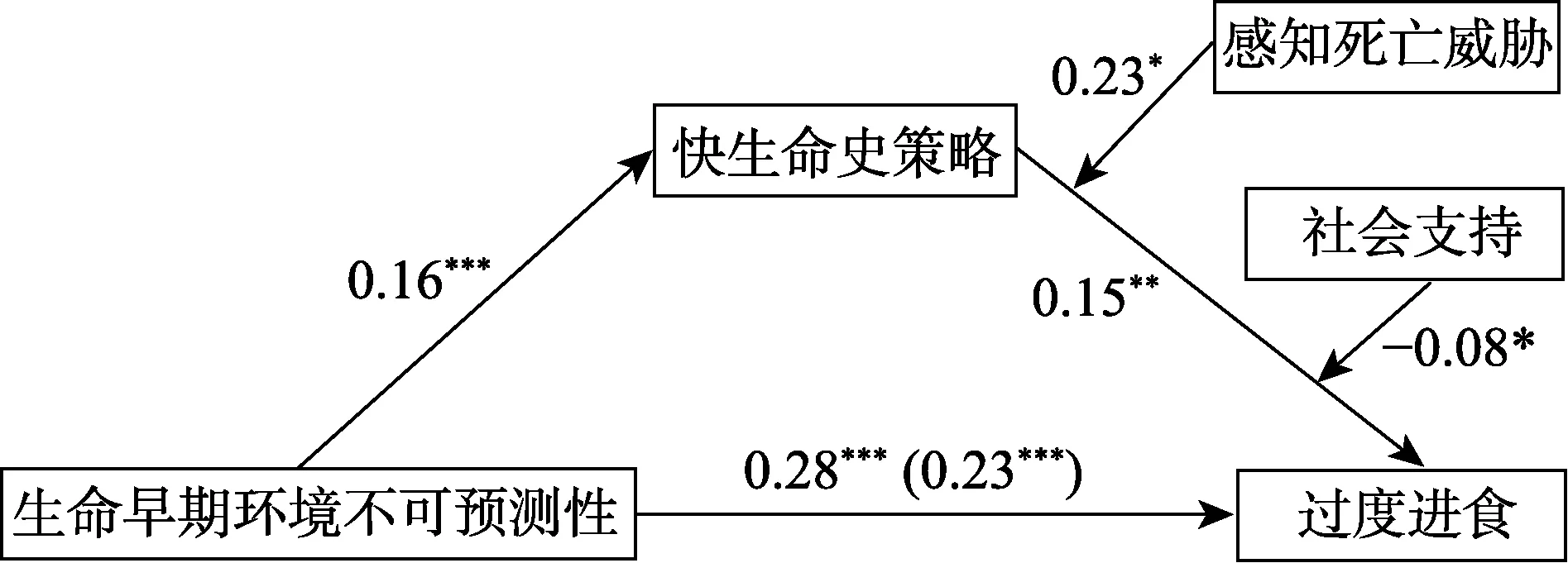
图3 有调节的中介模型(注:所有路径系数为标准化的路径系数)
3)采用SPSS 20.0 内process 插件(Model 16)(Hayes,2013)检验生命史策略是否在生命早期环境不可预测性对过度进食的影响中起中介作用,同时检验感知死亡威胁和社会支持在其中的调节作用,将年龄、性别和生命早期环境恶劣性作为协变量(图3)。结果表明快生命史策略在生命早期环境不可预测性和过度进食的关系中起中介作用,而感知死亡威胁和社会支持都可以调节该中介模型的后半段路径(见表3)。各调节变量在不同值时的中介效应不同(见表4)。简单效应分析显示,对于高死亡威胁组,快生命史策略正向预测过度进食(β
=0.34,SE
=0.06,t
=6.31,p
<0.001),对于低死亡威胁组,快生命史策略也可以正向预测过度进食(β
=0.16,SE
=0.06,t
=2.43,p
=0.016),但效应相对较小(图4a)。当社会支持较高时,快生命史策略无法影响过度进食(β=
0.12,t
=1.42,SE
=0.07,p
=0.159),而当社会支持较低时,快生命史策略正向预测过度进食 (β=
0.61,t
=8.15,SE
=0.06,p
<0.001) (图4b)。3.4 讨论
本研究在生命史视角下,通过两个研究揭示了生命早期环境不可预测性与过度进食的关系及其内在作用机制。研究结果一方面显示了生命早期环境不可预测性通过快生命史策略的中介作用影响过度进食;另一方面也进一步揭示了死亡威胁和社会支持的调节作用。在死亡威胁较高时,生命早期环境不可预测性通过快生命史策略的中介作用影响过度进食的效应会被放大,而社会支持较高时,这种效应可以被缓冲。研究结果对死亡威胁较高背景下过度进食的科学预防和干预具有重要的理论意义和实践价值,促进了心理学科在灾难中和灾难后更多地发挥作用。

表3 有调节的中介模型的回归分析

表4 不同条件下中介模型的间接效应量

图4 (a)死亡威胁对快生命史策略影响过度进食的调节作用;(b)社会支持对快生命史策略影响过度进食的调节作用;慢策略=生命史策略得分为负一个标准差(-1 SD),快策略=生命史策略得分为正一个标准差(+1 SD)
4 总讨论
本研究从进化视角出发,通过两种研究方法探索了生命早期环境不可预测性对过度进食的影响及其机制。研究1 以青少年为被试,证实了生命早期环境不可预测性对过度进食的直接影响;研究2采用问卷法选取新冠疫情背景下的被试进一步验证了研究结果,同时探究其作用机制。即生命早期环境不可预测性通过快生命史策略的中介作用对过度进食的影响在高死亡威胁下被扩大,而在高社会支持时可以被缓冲,研究结果具有较高的生态效度。
4.1 生命早期环境不可预测性对过度进食的直接影响
研究1 选取了处于生命早期的青少年群体,发现环境不可预测性影响饱食状态下个体的过度进食,研究2 回溯性地测量了新冠疫情背景下社区居民的生命早期环境不可预测性,也发现了其对过度进食的直接影响。研究结果具有一致性,都支持了生命史理论和以往童年经历较高环境不可预测性个体有更多过度进食的结论(Miller et al.,2018;Maner et al.,2017;Proffitt Leyva &Hill,2018)。生命史理论指出,由于总体资源有限,个体在各方面的投入必须要有所取舍;而不可预测性较高的环境代表着个体无法稳定地获得保证自身成长和发展所需的资源。因此,当面对即时奖赏(如食物)时,其生命史权衡的结果倾向于接受而不是拒绝它。除了过度进食之外,生命早期环境不可预测性还会导致个体的攻击行为,物质滥用等外化行为问题(Chang et al.,2019a,2019b;Lu &Chang,2019),提示了环境不可预测性对个体的问题行为具有跨领域的一致性。
4.2 生命史策略的中介作用与死亡威胁及社会支持的调节作用
研究2 还发现,生命早期环境不可预测性还会通过生命史策略的中介作用影响过度进食。国内外纵向研究表明环境不可预测性对快生命史策略的形成有重要的预测作用(Lu &Chang,2019;Chang et al.,2019b)。而近期研究则发现快生命史策略伴随的高冲动性倾向会增加过度进食(Cheon &Hong,2017)。过度进食与物质滥用和攻击行为一样遵循着生命早期环境不可预测性-快生命史策略-问题行为的作用模型,提示生命早期高环境不可预测性会促使个体对资源的权衡向快策略转变,即更加看重眼前的口腹之欲而忽视长远的健康风险,这会导致进食的数量和能量超过自身需求的行为,即过度进食。
同时,本研究还发现,高死亡威胁背景下个体快生命史策略对过度进食的预测更强,这支持了生命史理论和以往基于该理论的问题行为研究(Mittal&Griskevicius,2014),表明在高死亡威胁环境下,快生命史策略个体更可能将世界感知为不可预测和不安全的,因而会更多地选择眼前利益忽视长远风险。美味食物作为一种天然具有奖励性质的物品,可以为个体带来短暂的愉悦;但从长远角度来看,过度进食伴随着巨大的健康风险,极易导致肥胖和肥胖相关疾病。因此,在死亡威胁下,如何有效地减少过度进食行为,促进健康的生活方式是亟待探讨的问题。
此外,本研究还发现,社会支持能够有效缓冲这一效应,具体表现在低社会支持个体遵循着生命早期环境不可预测性-快生命史策略-过度进食这一作用机制,而高社会支持个体即便有较快的生命史策略,也可能不会过度进食。这与近期基于积极心理学的进食研究发现一致(陈曦梅 等,2020b),该研究提示良好的友谊质量可以促进个体基于自身饥饿线索进食,即“饿时吃,饱时停”。社会支持可以满足个体的社会性需要,使个体感受到被理解和关怀,进而肯定和关注自身的价值(Allgöwer,Wardle,&Steptoe,2001)。社会支持还会促进个体在压力情境时正视自己的内部感受,减少问题性进食行为(Wonderlich-Tierney &vander Wal,2010)。社会支持在生命早期环境不可预测性通过快生命史策略影响过度进食的路径中起缓冲作用,这一发现为较高死亡威胁情境下过度进食的干预提供了科学证据。
4.3 研究意义与实践启示
多篇元分析都发现,生命早期的压力环境会导致成年后的过度进食、肥胖相关的疾病和较短的生命周期(Danese &Tan,2014;Hemmingsson,2018;Hemmingsson,Johansson,&Reynisdottir,2014),但这一路径的内在机制缺乏进化角度的探讨。本研究基于生命史理论揭示心理本能对个体行为的巨大影响,从进化角度为该路径提供了合理解释。研究结果具有较大的理论和应用价值。一方面,研究深化了该理论对于进食领域的应用和解释;另一方面为过度进食和肥胖的预防干预提供了证据。
新冠背景下和灾后的心理援助可以从提升社会支持入手。研究显示,提升感知社会支持的干预可以提高个体的自我管理,进而提升健康积极的行为(McEwen,Pasvogel,Gallegos,&Barrera,2010)。以提升社会支持为核心的干预需要注意三方面,第一,心理援助的专业人员和同伴参与;第二,提供信息、认同和情感支持;第三,降低参与难度(Stewart,Simich,Shizha,Makumbe,&Makwarimba,2012)。在网络较为发达的现代,线上社会支持干预成为值得考虑的方式之一(White &Dorman,2001)。一方面它客观上克服了时间和空间的限制,降低了居民的参与难度;另一方面,参与者可以在任何地方以较为舒适的方式参与,杜绝了病毒在干预时传播的可能性。线上社会支持的研究发现,干预可以以社区形式开展,每周一次将同类心理困扰的参与者集中进行线上干预可以显著提高参与者的自我觉知,积极应对和感知社会支持,同时降低孤独感(Stewart,Barnfather,Magill-Evans,Ray,&Letourneau,2011)。因此,该方式不仅适用于受灾背景下,同时也有助于灾后居民心理和行为的积极提升。
4.4 研究的局限与未来展望
本研究还存在一些局限。首先,研究1 并未发现饥饿状态与环境不可预测性对实际进食量影响的交互效应。可能的解释是食物的种类有限,而被试本身对于食物的喜好影响了研究结果;未来研究可增加食物种类的选择。其次,研究1 中被试的饥饿和饱食状态是通过自我报告测量的。为提高研究的信度和效度,未来研究可参照Proffitt Leyva 和Hill (2018),将被试的生理指标(血糖)纳入测量。研究2 采用的是横向数据,无法建立因果联系。未来研究可以考虑采用追踪设计揭示生命早期不可预测性对过度进食的纵向机制和因果联系。
5 结论
(1)研究1 通过实验法发现生命早期环境不可预测性正向预测过度进食,表现在环境不可预测性可以显著正向预测青少年在饱食状态时的高热量食物份量选择。
(2)研究2 通过问卷法发现生命早期环境不可预测性通过生命史策略的中介作用间接影响过度进食,同时高死亡威胁会放大这一效应,而高社会支持则可以缓冲这一效应。
猜你喜欢
心理学报(2022年8期)2022-08-09
文萃报·周五版(2022年5期)2022-02-18
疯狂英语·新阅版(2022年1期)2022-01-26
奥秘(2020年3期)2020-04-01
作文评点报·高中版(2019年30期)2019-11-07
福建基础教育研究(2019年11期)2019-05-28
中小学心理健康教育(2017年34期)2017-12-19
环球时报(2017-11-22)2017-11-22
中学科技(2015年1期)2015-04-28
