
三峡水库蓄水后期香溪河库湾浮游甲壳动物群落结构
2020-10-15 09:44:46蔡庆华
水生生物学报 2020年5期
陈 星 叶 麟 谭 路 蔡庆华
(1. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京, 100049)
三峡水库是世界瞩目的大型水利工程, 2003年6月三峡水库首次成库(目标水位135 m), 2006年10月完成二期蓄水(目标水位156 m), 2008年10月完成三期试验性蓄水(目标水位175 m), 至2010年10月三峡水库达到175 m设计水位, 逐渐形成了水域面积达1084 km2的巨型水库, 三峡水库已成为我国最重要的淡水资源储存库之一。河流筑坝蓄水改变了河流自然连通的状态, 使得景观破碎化, 随着蓄水时间的增加, 水库逐渐由河流生态系统演变为湖泊生态系统, 这将改变生态系统的过程, 进而影响水生生物群落和生态系统功能[1—3]。水库被看作是河流与湖泊之间的过度环境, 其综合了自然变化过程(季节、气候)与生态系统管理(蓄水、泄洪、灌溉), 因此水库兼具有湖泊和和河流的某些特征[4]。
香溪河库湾作为三峡水库湖北库区最大的一级支流, 自香溪河库湾成库以来, 对香溪河库湾的研究陆续展开, 这些研究主要集中于水体理化环境[5]、悬浮物[6]、沉积物[7]、浮游植物[8,9]、浮游轮虫[10]、底栖动物[11]和微塑料[12]等方面。对香溪河库湾浮游甲壳动物的研究则集中于蓄水初期[13,14], 然而自2010年10月三峡水库首次正式完成175 m蓄水以来, 库区水体滞留时间进一步增加, 大坝周期性的蓄水、泄洪使得库区水位波动幅度变大, 水文条件的改变不仅会影响库区水体理化环境, 还会影响浮游甲壳动物群落的演替[15]。
浮游甲壳动物在水生食物网中扮演着重要的角色。浮游甲壳动物主要以细菌、轮虫和浮游植物为食, 同时又是鱼类的天然饵料, 因此能够将能量从初级生产者传递至高营养级[16]。当水体理化环境及其他水生生物群落发生改变时, 浮游甲壳动物能够迅速响应, 其多样性也能够用于指示水体受干扰的程度[17]。因此对三峡水库蓄水后期香溪河库湾的浮游甲壳动物群落展开研究是十分有必要的。
对香溪河库湾浮游甲壳动物已有不少研究, 这些研究主要集中于蓄水初期(2002—2004年)[13,14],对三峡水库蓄水后期香溪河库湾浮游甲壳动物群落时空变化规律的认识还不够深入。本文通过分析2015—2017年香溪河库湾浮游甲壳动物群落结构, 揭示香溪河库湾浮游甲壳动物的时空变化规律,探究水体理化环境与浮游甲壳动物的关系, 为三峡水库香溪河库湾的长期监测和管理提供数据基础。
1 材料与方法
1.1 研究区域
香溪河库湾位于三峡大坝上游38 km, 流经宜昌市兴山县、秭归县, 是三峡水库湖北库区最大的一条支流, 流域面积为3099 km2, 自然落差1540 m,年平均流量为65.5 m3/s[18]。自三峡水库蓄水后, 香溪河库湾下游二十余公里的河段逐渐演变为湖泊型水体[19]。根据水库分区理论[20], 将香溪河库湾分为库首、库中、库尾, 每个区域各设置一个采样点,分别以XX02、XX05、XX08表示(图 1)。本研究从2015年1月至2017年12月逐月对香溪河库湾浮游甲壳动物、水样进行采集。
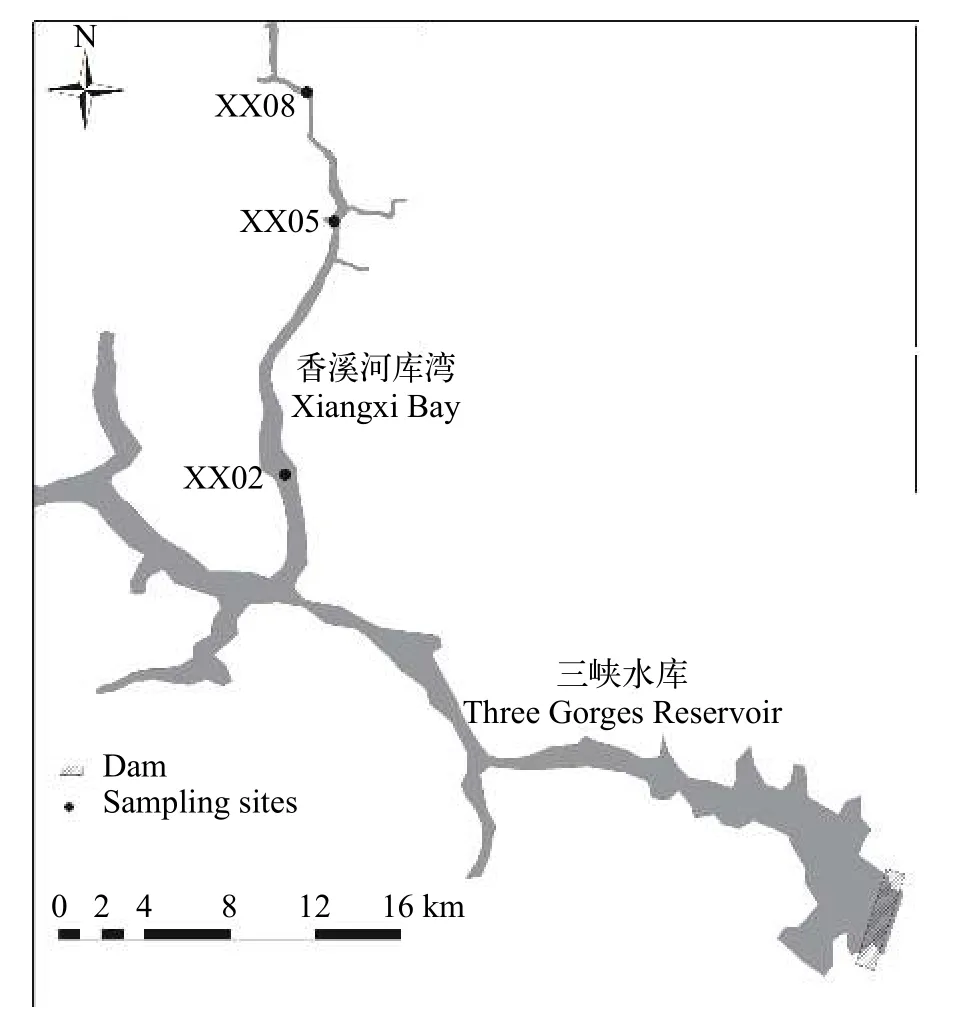
图1 采样点在三峡水库香溪河库湾中的位置Fig. 1 Location of sampling sites at Xiangxi Bay of the Three Gorges Reservoir
1.2 样品采集与分析
浮游甲壳动物定量样品采用5 L有机玻璃采水器采集表层水(0.5 m)共20 L, 经25 # 浮游生物网过滤后, 保存于100 mL的聚乙烯瓶中, 并加入4 %的福尔马林固定。将标本带回实验室静置48h后, 用吸管小心的吸取上清液, 将浓缩后的样品在解剖镜和显微镜下鉴定、计数。第五胸足是桡足类重要的分类学特征, 而剑水蚤科第五胸足褪化明显, 显微镜下往往难以观察。首先将0号昆虫针绑在细木棒末端制成解剖针, 将需解剖的标本小心挑出, 置于载玻片上, 滴若干甘油, 然后用解剖针从第四胸足开始解剖, 小心反复横拉, 连带第四胸足和第五胸足剖下进行观察, 必要时进行绘图。标本鉴定主要参考《中国动物志·节肢动物门·甲壳纲·淡水枝角类》[21]、《中国动物志·节肢动物门·甲壳纲·淡水桡足类》[22]、《长江流域的枝角类》[23]和《Freshwater Crustacean Zooplankton of Europe》[24]。浮游甲壳动物生物量依据体长-体重回归方程计算[25]。
现场用温深仪 XR-420(RBR公司)测量水深(Depth), 并用塞氏盘测量水体透明度(SD)。采用多参数水质仪YSI6600(美国)测量表层水水温(WT)、溶解氧(DO)、电导率(Cond)、浊度(Tur)和pH。用5 L采水器采集表层水, 取水样保存于清洁的塑料瓶中, 并加入适量的硫酸, 调节pH至2以下, 放于4℃冰箱保存。将水样带回实验室后使用连续自动分析仪SAN+++(荷兰SKALAR公司)测量总氮(TN)、硝氮(NO3-N)、氨氮(NH4-N)、总磷(TP)、磷酸盐(PO4-P)、可溶性硅(DSi)和溶解碳(DOC)。另取一部分已知体积的水样经Whatman GF/C滤膜过滤,滤膜保存于黑暗4℃的冰箱中, 采用分光光度计法测定叶绿素a(Chl.a)[26]。
1.3 数据分析
将环境因子、浮游甲壳动物密度和生物量lg(x+1)转化后, 采用方差分析水体理化指标、密度和生物量的时空差异, 组间多重比较采用TukeyHSD的方法进行。采用相似性分析检验浮游甲壳动物群落时空差异。
采用广义线性模型分析浮游甲壳动物密度和生物量与环境因子的关系。为了使AIC值最小, 每次自动对一个解释变量进行逐步逆向选择, 得到最简化模型。误差分布参数设定为高斯分布, 与此同时我们还检查了模型的膨胀因子(VIF), 并将VIF>4的因子予以剔除, 以确保最终用于分析的预测因子之间不存在多重共线性, 并通过计算偏差解释率确定关键的预测变量。最后, 我们对残差图进行了检查, 以确保没有偏离正态性和同方差。
传统的生物多样性指数(物种丰富度、香农维纳多样性指数和辛普森指数等)与采样强度和物种丰度分布密切相关, 不同采样强度的生物多样性指数往往缺乏可比性[27]。除此以外, 物种丰富度不包含任何关于物种丰度或相对丰度的信息, 稀有种和常见种具有相同的权重[28]。近年来Hill numbers越来越多的被用于描述分类、系统发育、功能多样性[29—32], 一些生态学家对Hill numbers进行了插值和外推, 提出了一种基于个体(丰度)和基于样本量(发生率)的自疏曲线, 以便于不同采样强度生物多样性数据的比较[28]。在本研究中每月仅对香溪河库湾3个样点浮游甲壳动物进行了采集, 可能会由于采样强度过低而低估了浮游甲壳动物生物多样性, 为了避免这种错误, 我们采用R软件“iNEXT”包构建了浮游甲壳动物自疏曲线模型, 比较浮游甲壳动物生物多样性时空差异[33]。
对物种-样点矩阵标准化处理后, 进行降趋势对应分析(Detrended Correspondence Analysis, DCA),各排序轴最大值均超过4, 说明单峰模型更适合排序, 因此采用典范对应分析(Canonical Correspondence Analysis, CCA)探索浮游甲壳动物群落与环境因子的关系[34]。采用前项选择和蒙特卡洛置换检验(置换次数999次)对解释变量进行筛选, 得到简约CCA排序图。以上所有过程均在R软件“vegan”包中完成[35]。
其他数据的可视化和统计分析过程均在R软件(3.51版本)中完成。
2 结果
2.1 环境因子
香溪河库湾水体理化指标在各年度平均值、标准差如表 1示。方差分析的结果表明, TN浓度在各样点间的差异不明显(P=0.44); NO3-N、TP、PO4-P浓度在各样点间差异明显(P<0.05), TukeyHSD组间多重比较表明NO3-N在XX08-XX02和XX08-XX05具有显著性差异(P<0.05); TP和PO4-P三个样点彼此间均存在显著性差异(P<0.05)。NO3-N整体上表现为由库尾向库首增加, TP和PO4-P整体上表现为由库尾向库首减少。各样点TN、NO3-N、TP和PO4-P年际变化如图 2所示, 各种营养盐浓度在每年3—4月呈下降趋势, 5—7月明显增加, 并呈现出周期性变化。方差分析的结果表明TN、NO3-N、TP、PO4-P和WT在不同年份间具有显著性差异(P<0.05), 其余环境因子在年际间不具有显著性差异。
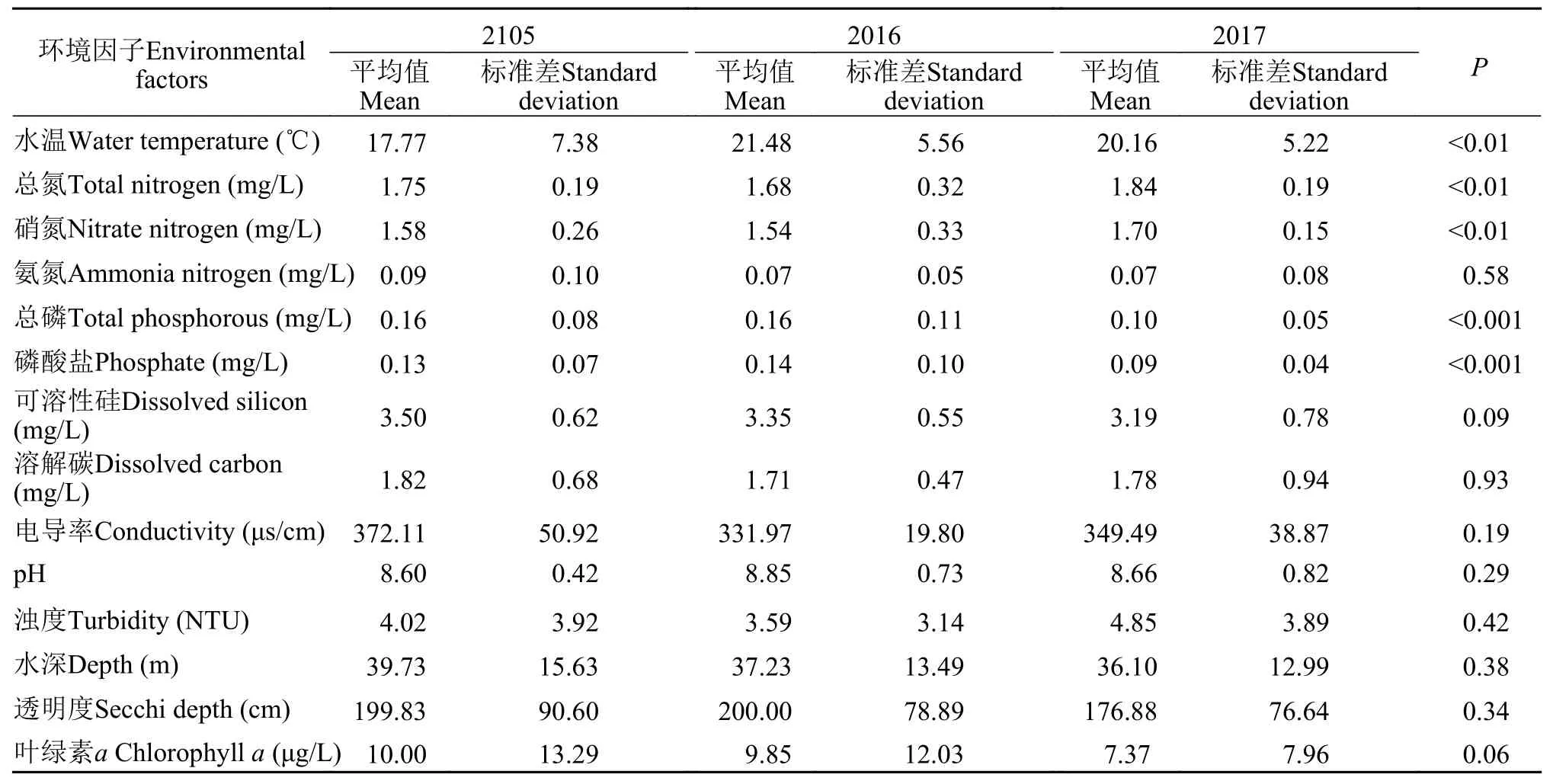
表1 三峡水库香溪河库湾水体理化指标的统计描述Tab. 1 Statistical description of physical and chemical indexes of Xiangxi Bay of the Three Gorges Reservoir
2.2 群落组成
共采集浮游甲壳动物23种, 其中枝角类10种,桡足类13种, 优势种类年际间具有一定差异(表2)。2015年出现频率较高的物种为僧帽溞(Daphnia cucullata, 30.6%)、简弧象鼻溞(Bosmina coregoni, 38.9%)、英勇剑水蚤(Cyclops strenuuss, 30.6%)、广布中剑水蚤(Mesocyclops leuckarti, 33.3%)和跨立小剑水蚤(Microcyclops varicans, 47.2%), 相对丰度分别为48.7%、13.0%、1.4%、1.7%和20.0%。2016年出现频率较高的物种为简弧象鼻溞(22.2%)、广布中剑水蚤(36.1%)和跨立小剑水蚤(36.1%), 相对丰度分别为10.8%、12.5%和14.9%。2017年出现频率较高的物种为短尾秀体溞(Diaphanosoma brachyurum, 25%)、蚤状溞(Daphnia pulex, 27.8%)、僧帽溞(30.6%)和跨立小剑水蚤(38.9%), 相对丰度分别为2.9%、11.8%、18.8%和31.0%。相似性分析表明浮游甲壳动物群落组成在年际间的差异不显著(P=0.31), 在香溪河库湾不同区域间的差异同样不显著(P=0.10)。
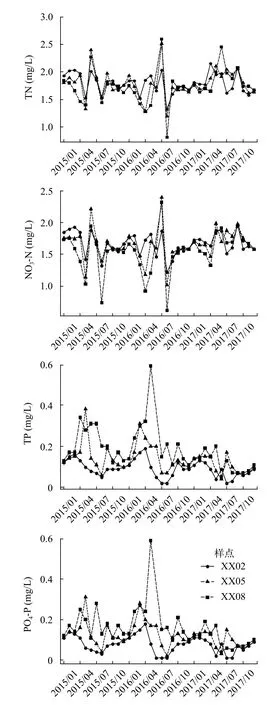
图2 三峡水库香溪河库湾TN、NO3-N、TP、PO4-P时空变化Fig. 2 Spatial and temporal variations of TN, NO3-N, TP and PO4-P in Xiangxi Bay of the Three Gorges Reservoir
浮游甲壳动物密度和生物量均呈现出明显季节性变化规律(图 3), 春季和夏季具有较高的密度和生物量, 秋季和冬季具有较低的密度和生物量。2015年6月浮游甲壳动物密度最大值为16.17 ind./L,2015年5月浮游甲壳动物生物量最大值为0.70 mg/L。每年1—2月均未能采集到浮游甲壳动物标本。群落随季节变化规律基本相同, 4月小型种简弧象鼻溞具有较大的密度, 随后5月和6月大型种类蚤状溞、僧帽溞占据优势, 同时具有较高的生物量, 跨立小剑水蚤除1月和2月外, 其他月份均有检到, 在6月具有较大的密度。广布中剑水蚤虽然在大部分月份均有检到, 但其密度和生物量较低。浮游甲壳动物密度和生物量在年际间呈下降趋势, 密度和生物量在年际间均具有显著性差异(P<0.05)。
2.3 生物多样性
基于个体的自疏曲线表明XX02浮游甲壳动物物种丰富度观测值低于外推值, 香农指数观测值则与外推值差别不大。其余区域物种丰富度和香农指数观测值与外推值差别不大(图 4)。XX02物种丰富度的实际观测值为19, 外推值为21, 样本覆盖率估计值为99.8%。XX05物种丰富度和香农指数均高于XX02、XX08, XX02和XX08的物种丰富度、香农指数具有一定的重叠。自疏曲线还表明我们对XX02的采样强度过低, 获取的个体数太少,可能低估了该区域的物种丰富度。
物种丰富度和香农指数季节差异如图 5所示。浮游甲壳动物物种丰富度在各季节从高到低依次为春季、夏季、秋季和冬季, 春季和夏季物种丰富度外推值具有较大的重叠, 冬季由于个体数偏少,模型并未能给出外推值。浮游甲壳动物香农指数在各季节从高到低依次为秋季、冬季、春季和夏季, 同样由于冬季个体数偏少, 模型也未能给出外推值。秋季物种丰富度和香农指数的观测值均低于外推值, 春季和夏季观测值与外推值差别不大。表明我们对秋季和冬季的采样强度过低, 可能会低估了浮游甲壳动物的物种丰富度和香农指数。
2.4 浮游甲壳动物群落与环境因子的关系
广义线性线性模型表明Chl.a、NH3-N和DOC对浮游甲壳动物密度具有显著影响, 解释变量对模型的解释率分别为18.9%、9.5%和0.3%, 因此Chl.a是影响浮游甲壳动物密度最关键的因子, 其次为NH4-N, 尽管DOC对模型解释率较低, 但其能显著减小模型AIC值。广义线性模型同样表明Chl.a、WT、NO3-N和PO4-P对浮游甲壳生物量具有显著影响, 解释变量对模型的解释率为27.6%、9.6%、2.4%和1.6%, Chl.a同样是影响浮游甲壳动物生物量最关键的因子, 其次为WT, PO4-P对生物量的影响不如前两者。
CCA排序轴前两轴的特征值分别为0.192、0.089, 对群落变异的解释率分别为10.78%和9.41%,蒙特卡洛置换检验结果表明温度(WT)、叶绿素(Chl.a)和水深(Depth)对浮游甲壳动物群落具有显著性影响(P<0.05)。WT、Chl.a和Depth与排序轴一的相关性系数分别为0.986、0.459和-0.539, 排序轴一代表了WT梯度。WT、Chl.a和Depth与排序轴二的相关性系数分别为-0.154、0.881和-0.140,排序轴二代表了Chl.a梯度。蚤状溞、僧帽溞、台湾温剑水蚤(Thermocyclops taihokuensis)和网纹溞属(Ceriodaphniasp.)等种类密度与叶绿素呈正相关, 短尾秀体溞、圆形盘肠溞(Chydornus sphaeri-cus)和点滴尖额溞(Alona guttata)密度与水深呈正相关, 广布中剑水蚤、锯缘真剑水蚤(Eucyclops serrulatus)、毛饰拟剑水蚤(Paracyclops fimbriatus)和跨立小剑水蚤密度与水温呈正相关(图 6)。
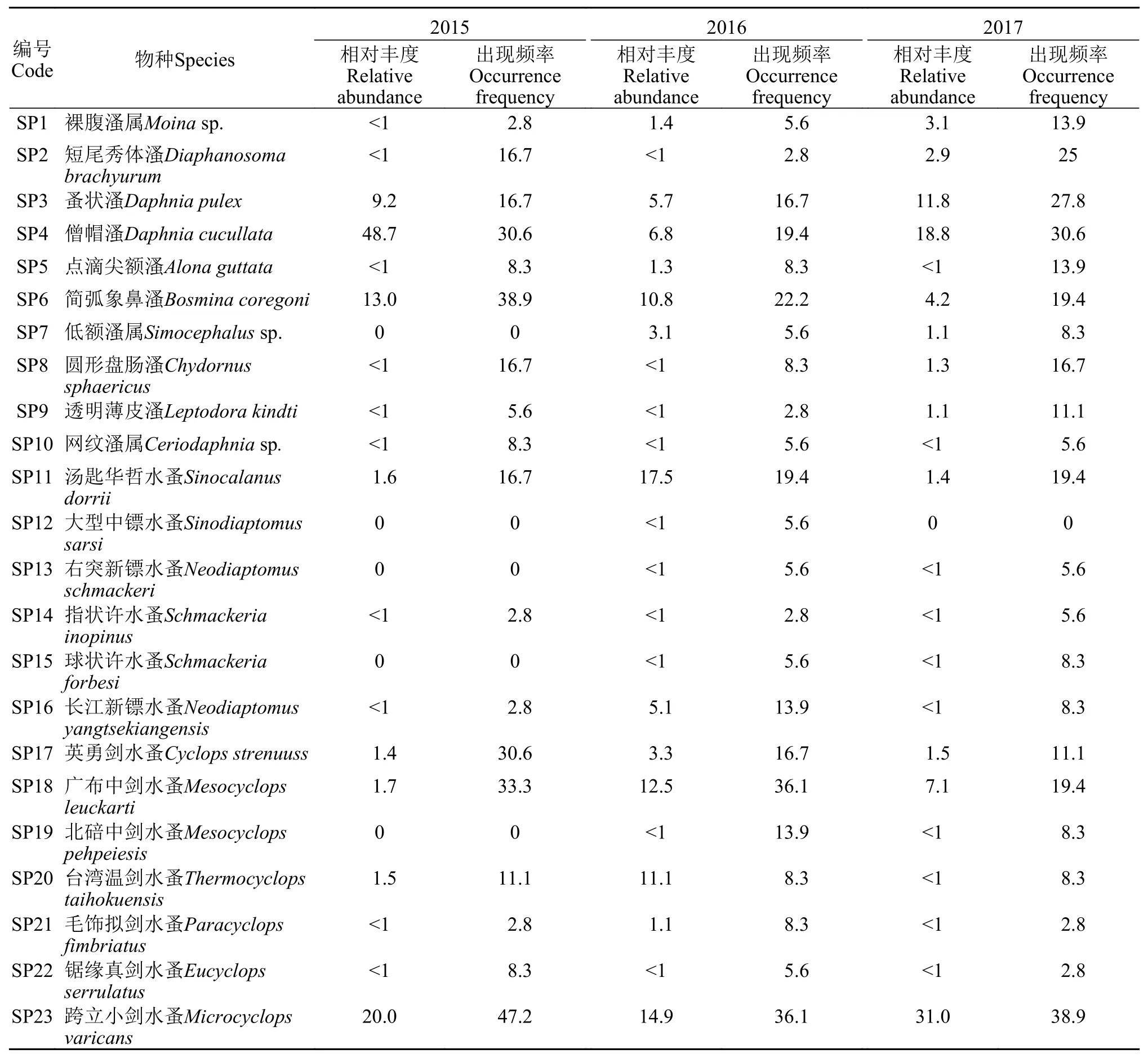
表2 三峡水库香溪河库湾浮游甲壳动物不同年际相对丰度、出现频率Tab. 2 The relative abundance and frequency of crustacean zooplankton in Xiangxi Bay of Three Gorges Reservoir in different years (%)
3 讨论
3.1 环境因子及浮游甲壳动物群落特征
TN、NO3-N、TP、PO4-P和WT在年际间具有显著性差异, 说明香溪河库湾水体部分理化因子仍然处于不稳定的状态。TN、NO3-N、TP和PO4-P具有明显的季节变化规律, 且具有一定周期变化。各营养盐呈现出纵向分布格局, 如TP和PO4-P浓度从库尾至库首逐渐降低, Seda对捷克峡谷型水库的研究同样发现类似的纵向梯度分布格局[36]。
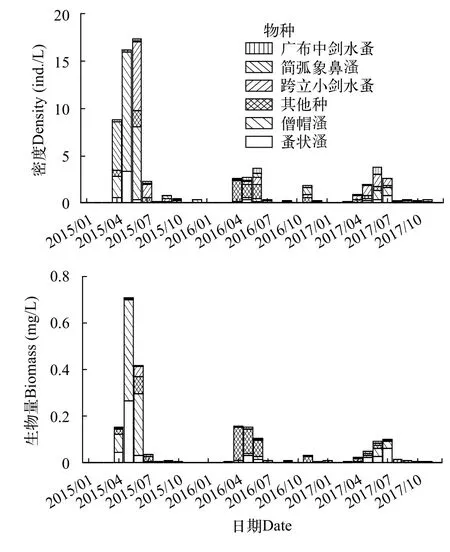
图3 三峡水库香溪河库湾浮游甲壳动物密度、生物量时间变化Fig. 3 Temporal variations of crustacean zooplankton density and biomass in Xiangxi Bay of the Three Gorges Reservoir
共检出浮游甲壳动物动物23种, 其中枝角类10种, 桡足类13种, 相比于蓄水初期仅鉴定18种, 浮游甲壳动物动物物种数有一定增加, 物种组成也有一定的差异[13,14]。以往的研究表明河流筑坝改变了水体连通状态, 使得浮游甲壳动物生物多样性降低[37], 这与本研究的结果相反。可能是由于随着库龄的增加, 水动力条件较筑坝初期逐渐趋于稳定,使浮游甲壳动物多样性增加。出现频率较高的物种包括简弧象鼻溞、广布中剑水蚤、跨立小剑水蚤、短尾秀体溞、蚤状溞、僧帽溞和英勇剑水蚤,均为长江中下游地区常见的种类[21—23,38]。香溪河浮游甲壳动物密度最大值仅为16.17 ind./L, 生物量最大值仅为0.70 mg/L, 均处于较低的水平, 以往的研究同样发现浮游甲壳动物在大型深水水库的密度和生物量处于较低的水平[39]。
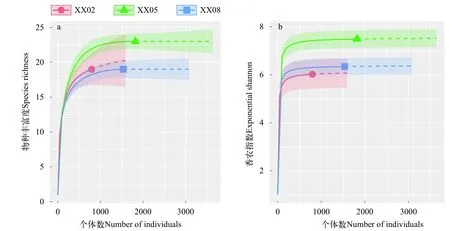
图4 香溪河库湾不同区域基于个体自疏曲线Fig. 4 Individual-based rarefactions curve for each areas in Xiangxi Bay
3.2 浮游甲壳动物多样性
由于样本量过少, 我们可能低估了库首(XX02)物种丰富度(图 5), 浮游甲壳动物生物多样性指数在库中(XX05)高于库首(XX02)和库尾(XX08)。Pinell-Alloul对浮游甲壳动物空间异质性有过详细的论述, 他认为嵌套和斑块是淡水和海洋浮游甲壳动物的共同特征, 当空间尺度超过1 km, 非生物因子对维持空间异质性占主导作用[40], 生境适当程度的异质性和不稳定性也有利于维持较高的物种丰富度[41]。香溪河库湾不同区域营养盐和叶绿素存在一定的差异, 而且库中相较库首和库尾受大坝蓄水回水顶托和泄洪影响也相对较小, 环境因子的差异和适当的水文波动可能是造成库中生物多样性高于库首和库尾原因。

图5 香溪河库湾不同季节基于个体自疏曲线Fig. 5 Individual-based rarefactions curve for each seasons in Xiangxi Bay
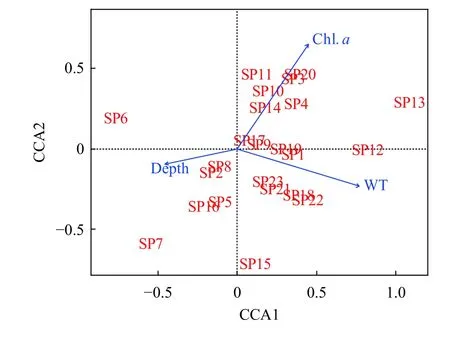
图6 浮游甲壳动物与环境因子的典范对应分析排序图Fig. 6 Ordination plot of canonical correspondence analysis for crustacean zooplankton with selected environmental variables
3.3 浮游甲壳动物群落与环境因子的关系
以往的研究表明水温对浮游甲壳动物群落具有重要影响, 如浮游动物生长、繁殖和产卵等生理学特征[42]。Beaver等[43]对美国西部102个水库的浮游甲壳动物进行了研究, 发现浮游动物群落组成、现存量和粒径大小与温度密切相关, Shurin等[44]的研究则表明较大温度的波动能够支持更高物种丰富度。香溪河库湾的研究同样反映了这一规律, 浮游甲壳动物在春季和夏季(5月和6月)具有较高的密度和生物量, 每年冬季(1月和2月)均未能采集到标本, 夏季则具有较高的物种丰富度。温度一定程度的升高还会使得浮游甲壳动物群落以大型种类为主[45]。4月小型种简弧象鼻溞具有较大的丰度, 随后5月和6月大型种类蚤状溞、僧帽溞占据优势, 广义线性模型同样表明生物量与温度呈正相关, 因此温度的波动可能是群落季节变化的驱动因子。
叶绿素浓度在某种程度上可以代表浮游甲壳动物食物来源。食物的可获得性以及对食物的竞争也是影响浮游甲壳动物行为(垂直分布)和群落组成的重要因子[46]。本研究发现Chl.a存在明显的季节变化, 春季和夏季浓度较高, 冬季浓度则较低, 这也使得香溪河库湾浮游甲壳动物存在明显的季节差异, 蚤状溞、僧帽溞和网纹溞属等大型种在夏季占据优势, 秋季圆形盘肠溞和点滴尖额溞等小型种在秋季和早春占据优势。大型枝角类通常对浮游植物具有较高的摄食压力, 小型枝角类多以水体中的细菌和悬浮物为主要食物来源。以往的经验往往认为产毒素的蓝藻通过毒害作用能够抑制大型枝角类对浮游植物的摄食, 某些极端情况还能造成浮游动物的死亡[47]。然而近年来一些生态学家通过一系列控制实验, 证明即使是高浓度的微囊藻毒素也并不妨碍大型枝角类对浮游植物的摄食[48]。春季和夏季浮游甲壳动物同样具有较大的密度和生物量, 广义线性模型也表明叶绿素浓度与密度和生物量呈正相关, 且是最重要的环境因子, 这说明食物资源的增多, 能够维持更高密度和生物量。尽管营养盐对浮游甲壳动物密度和生物量也具有一定影响, 但不如温度和叶绿素显著(表 3)。叶绿素浓度和温度的季节性变化可能是香溪河库湾浮游甲壳动物密度和生物量季节演替的重要因素。
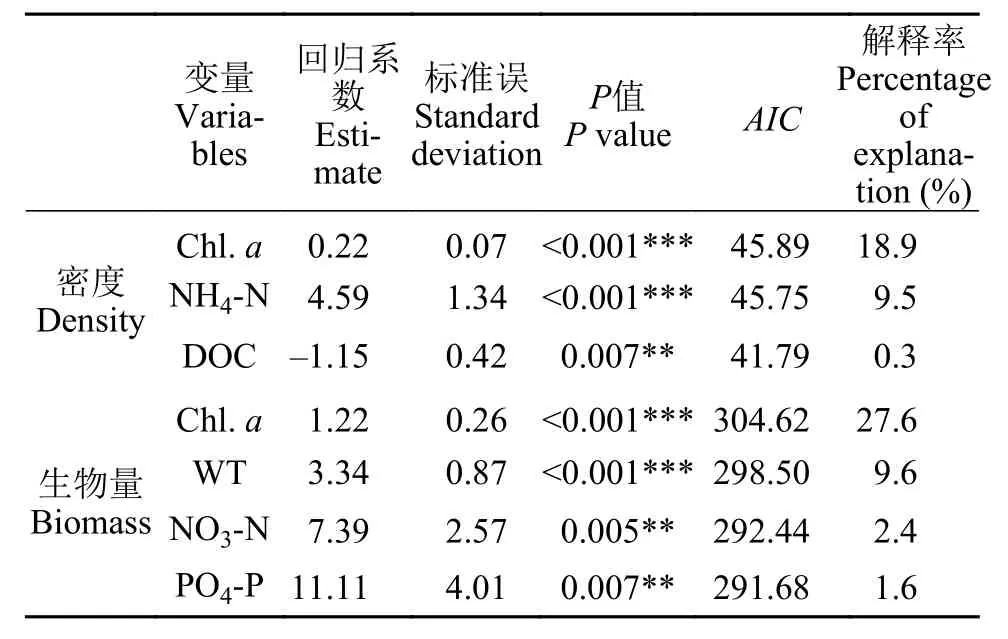
表3 广义线性模型的统计结果Tab. 3 Statistic results of general linear models
我们的研究还表明水深是影响浮游甲壳动物群落变异的环境因子。应该指出的是香溪河库湾水位呈现出季节周期性变化, 具体表现为春夏季水位较低, 秋冬季水位较高, 这主要是受大坝季节性蓄水、泄洪影响。以往的一些研究表明水文条件(水体滞留时间、流速和流量等)对浮游甲壳动物群落有重要影响[49], 如Burdis等[50]对密西西比河上游天然河流湖泊的研究发现水体滞留时间是影响浮游甲壳动物群落最重要的生态因子。由于缺乏具体的流量、流速等数据, 用水深评估水动力条件对浮游甲壳动物的影响显然是不够准确的。
4 结论与展望
三峡水库水环境与水生态研究的三峡水库蓄水后期, 香溪河库湾浮游甲壳动物大多为长江流域常见种类, 且水温、营养盐、密度和生物量均处于不稳定状态, 物种丰富度较蓄水初期有所增加, 生物多样性也呈现一定的纵向分布格局。值得注意的是CCA分析的结果表明环境因子对浮游甲壳动物变异解释率仅为20.19%, 表明环境因子并不能完全解释香溪河库湾浮游甲壳动物群落变异。因此通过方差分解等统计方法评估水动力条件和水体理化因子对浮游甲壳动物综合影响可能是未来一个值得研究的方向, 除此以外, 本文仅研究了香溪河库湾浮游甲壳动物3年的变化, 基于浮游甲壳动物密度和生物量仍然处于不稳定的状态, 香溪河库湾浮游甲壳动物长时间序列变化、群落稳定性及其维持机制和对极端洪水的响应都是未来值得研究的问题。
猜你喜欢
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
东方考古(2020年0期)2020-11-06 05:34:02
中国水利(2020年14期)2020-08-02 16:19:30
创新作文(小学版)(2016年10期)2016-11-11 05:45:20
创新作文(1-2年级)(2016年4期)2016-05-14 12:45:16
湖南农业(2015年7期)2015-03-20 01:48:37
湖泊科学(2014年4期)2014-02-27 10:30:32
中国工程咨询(2011年11期)2011-02-13 02:35:00
