
加勒比松3个变种种源试验初报
2020-10-09 09:11:00邓乐平赵奋成郭文冰黄炬峰谭志强吴惠姗
广东农业科学 2020年8期
邓乐平,黄 婷,赵奋成,郭文冰,黄炬峰,谭志强,吴惠姗,王 哲
(1.台山市红岭种子园,广东 台山 529223;2.广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广东 广州 510520)
【研究意义】木材是世界公认的四大原材料之一,随着我国经济的快速发展,木材需求量日益增长,实际消耗远远超过目前我国森林资源结构所决定的产量。据统计,2016年我国原木的进口量增加到4 910.3万m3,其中针叶树原木的数量占总量的50%以上[1]。随着国家天然林保护工程的实施,木材供给进一步减少。我国南方是商品林发展和木材生产的重点区域,在南方集体林区大力推进速生丰产林、工业原料林的发展是解决当前木材供需矛盾的重要途径之一。【前人研究进展】湿加松是以湿地松(Pinus elliottiiEngelmann)为母本、加勒比松(P.caribaeaMorelet)为父本的杂交后代,由于其拥有双亲生长快、适应性强、产脂量高、干形通直的特性,成为我国华南地区重要的人工造林树种[2]。广东省林业科学研究院与台山红岭种子园是我国最大的湿加松研究、生产基地[3],自1991年开始合作进行湿地松和加勒比松杂交育种研究,选育出了一批优良的杂种家系并迅速在广东及周边省区推广应用。【本研究切入点】湿地松原产于美国东南部,其适应气候环境为亚热带气候,在我国南到广东、广西,北至湖北、安徽,生长表现良好[4-7]。加勒比松属热带树种,20世纪60年代我国先后引入多批加勒比松,其中包含了大量的遗传变异。然而由于加勒比松耐寒性差、造林保存率低等原因,对加勒比松改良的热潮迅速降温,大量优异的加勒比松种质资源未得到良好保存,基因资源流失严重,目前仅在海南、广东和广西南部等热带和亚热带南部地区有零星分布[8-13]。【拟解决的关键问题】选育的湿加松良种遗传其父本耐寒性不足的缺点,偏北地区冬季的低温会伤害新生枝条导致生长受阻,限制了湿加松进一步往偏北地区推广种植。本试验从加勒比松原产地引入种源,旨在拓宽当前加勒比松遗传基础,筛选耐寒能力强的亲本,指导杂交配合,创制抗旱湿加松新种质,扩大湿加松在我国推广种植范围。
1 材料与方法
1.1 试验地概况
造林地位于台山红岭种子园(22°10′N,112°49′E),属南亚热带海洋性气候,年均温21.8 ℃,年降水量1 940 mm,平均海拔30 m,土壤为花岗岩发育的酸性砖红壤,pH值5~5.5,土层深厚。自然条件适宜加勒比松、湿地松生长。
1.2 试验材料
参试材料共有4个树种(变种)的22个种源/种批(表1),其中,湿地松(PEE)种源/种批5个、古巴加勒比松(PCC)种源/种批3个、巴哈马加勒比松(PCB)种源/种批2个、洪都拉斯加勒比松(PCH)种源/种批12个,提供种子的国家有洪都拉斯、古巴、澳大利亚、巴西、美国、加拿大、中国等。
1.3 试验方法
试验苗木于2014年9月育苗,2015年9月种植造林。试验采用完全随机区组设计,23个重复,根据试验地地形,其中19个重复为5株单行小区,3个重复为18株3行小区,1个重复为15株3行小区,株行距3 m×3 m。
1.4 数据收集与统计分析
2016年对1年生测定林株高H进行每木调查,2019年对4 年生测定林树高H和胸径D进行每木调查。利用树高、胸径计算单株材积V,计算公式为:

表1 湿地松、加勒比松种源/种批的基本概况Table 1 Basic profile of the provenances or batches of slash pine and Caribbean pine

式中,π为圆周率,f为形数,取f=0.5。采用SAS软件的GLM过程进行方差分析[14]及多重比较。以产自台山的PEE为对照计算生长量增益(Gi),计算公式为:
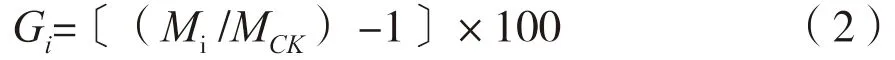
式中,Mi为被评价的参试材料生长量平均值,MCK为对照的生长量平均值。
由于古巴加勒比松和巴哈马加勒比松种源/种批数量相对较少,仅对拥有12个种源/种批的洪都拉斯加勒比松数据估算生长性状的遗传参数,利用统计软件ASReml估算洪都拉斯加勒比松各个试验因子的方差分量,根据方差分量计算不同年份各性状的遗传力及标准误[15-16]。数据分析的线性模型如下:
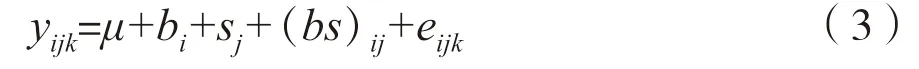
式中,yijk为单株观测值,µ为群体平均值,bi为区组效应,sj为种源效应,(bs)ij为区组与种源互作效应,eijk为误差效应。在模型试运行过程中,区组为固定效应,其他因素为随机效应。
种源遗传力计算公式为:
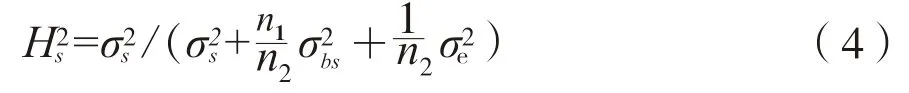

式中,rg、rp分别为遗传相关系数和表型相关系数,covg(x,y)、covp(x,y)分别为遗传协方差,分别为性状x和性状y的遗传方差,分别为性状x和性状y的表型方差。
使用R 3.4.3软件对4年生洪都拉斯加勒比松的平均树高、胸径、单株材积与相对应产地的纬度、经度、海拔3个因子进行简单相关分析,分析时排除12、18号两个南半球种源。
2 结果与分析
2.1 不同年度保存率分析
不同树种(变种)、不同种源/种批在1年生和4年生时苗木的保存率见表2。在1年生时,古巴加勒比松的保存率最低(82.64%),洪都拉斯加勒比松的保存率最高(93.20%);到4年生时,不同树种(变种)的保存率变化较大,其中古巴加勒比松的保存率依然最低(44.76%),而湿地松的保存率最高(74.95%)。在种源水平,洪都拉斯加勒比松5号种源1年生时100%存活,巴哈马加勒比松16号种源1年生保存率达99.13%,湿地松23号种源1年生保存率只有56.52%,该种源的苗木数量较少(23棵)有可能造成较大的误差。1年生和4年生保存率相差最大的是古巴加勒比松的3个种源,均超过36%,而湿地松24号种源前后保存率相差只有2%,同时也是4年生时保存率最高的种源(95.04%),而产自我国的古巴加勒比松(13号)保存率最低、只有38.18%,产自我国的巴哈马加勒比松 4年生保存率也远低于巴西种源。
2.2 种源试验林生长分析
利用单株数据作方差分析,由结果(表3)可知,1年生树高和4年生树高、胸径、单株材积在区组间、树种(变种)间、树种(变种)内种源间的差异均达到极显著水平,说明需要作进一步的比较和筛选。
4个树种(变种)的生长量多重比较结果见图1,以1年生洪都拉斯加勒比松的平均树高最高,巴哈马加勒比松和湿地松次之,古巴加勒比松最小(图1A);到4年生时,以巴哈马加勒比松平均树高最高,显著超过洪都拉斯加勒比松,其余3个树种(变种)平均树高差异不显著(图1B);4年生洪都拉斯加勒比松胸径最大,巴哈马加勒比松稍小,差异不显著,这2个变种的胸径均显著大于湿地松和古巴加勒比松(图1C)。4个树种(变种)4年生单株材积分成两个梯队,巴哈马加勒比松和洪都拉斯加勒比松在第一梯队,湿地松和古巴加勒比松在第二梯队(图1D)。

图1 不同树种/变种生长量年度变化Fig.1 Growth changes of different tree species/variants in different years

表4 种源试验林生长及增益情况Table 4 Growth and gain of provenance test forest
由表4可知,1年生平均树高最大的是1号种源;4年生时,树高生长以巴哈马加勒比松的16号种源最大,同时也是单株材积生长最优的种源,与对照(湿地松)相比,单株材积增益高达75.73%。其中,在洪都拉斯加勒比松变种内的12个种源间,1、2号种源的单株材积明显小于对照,5、7、8、9号种源单株材积与对照相当,3、4、6、10、12、18号种源单株材积增益较高,其中10、12、18号种源单株材积显著高于对照;古巴加勒比松的3个种源单株材积增益均为负值;古巴加勒比松13号种源和巴哈马加勒比松14号种源单株材积增益分别为-29.03%和-28.17,这两个种源产地均为我国湛江市遂溪县;湿地松23、25号种源单株材积小于对照,但未达到显著水平。树高性状在1年和4年各个种源的排名变化较大,两个年份的相关系数为0.3298,标准误较大、为0.2053,达显著水平。
2.3 种源试验林遗传参数估算
洪都拉斯加勒比松12个种源的数据用于估算生长性状的遗传参数,结果见表5,在种植后1年树高的遗传力达到0.9822,同时标准误较小;种植后4年各个性状的广义遗传力也均高于0.8。
所有种源的遗传相关和表型相关分析结果见表6,树高、胸径、单株材积3个性状之间的遗传与表型均呈极显著正相关,相关系数在0.730~0.984之间;遗传相关系数稍大于表型相关系数,树高与胸径的相关性相对最小,且遗传相关系数的标准误相对较大;胸径与单株材积的相关系数超过0.970,表明可通过胸径性状间接选择单株材积优秀的个体或种源。

表5 洪都拉斯加勒比松生长性状的广义遗传力Table 5 Broad heritability of growth traits of Honduran Caribbean pine
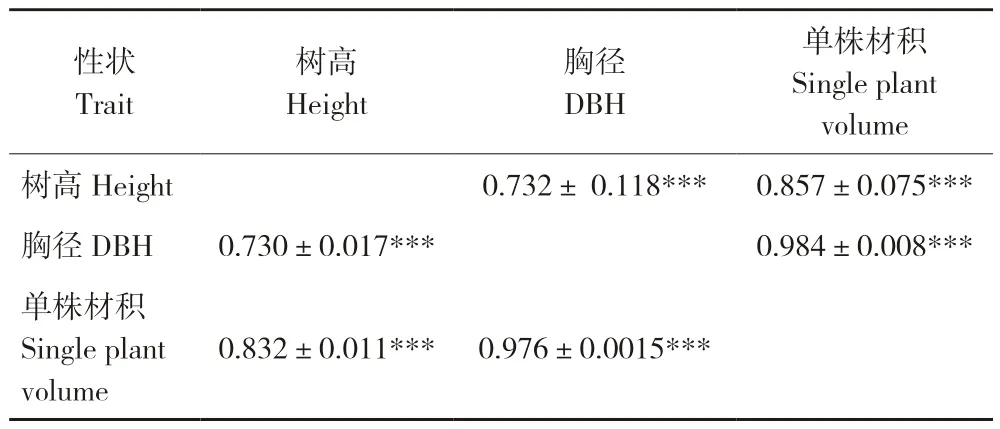
表6 性状间的遗传相关与表型相关系数Table 6 Genetic correlation and phenotypic correlation coefficient among different traits
洪都拉斯加勒比松生长性状与地理因子相关分析结果(表7)表明,生长量与纬度和经度呈负相关,与海拔呈正相关,但都未达到显著水平。

表7 生长性状与地理因子相关分析Table 7 Correlation analysis of growth traits and geographic factors
3 讨论
本试验分析了加勒比松部分种源种子的发芽率,其中洪都拉斯加勒比松平均发芽率为54.34%,古巴加勒比松平均发芽率只有38%,与王观明等[8]种源试验的发芽率相当。韩超等[17]统计加勒比松种子浸泡24 h后除去漂浮在水面上不饱和的种子,播种后发芽率达到70.8%,与同期播种的思茅松发芽率接近,但加勒比松种子浸泡时浮在水面上的比例明显更大,表明加勒比松种子发育不良现象相对较严重。
已有多个加勒比松种源试验、子代测定试验研究结果显示,在加勒比松3个变种中,洪都拉斯加勒比松的多年生保存率相对最低[11-13,18],这主要是由于洪都拉斯加勒比松的抗性较差,面对主要病虫害松稍螟和枯叶病时受害最严重,古巴加勒比松和巴哈马加勒比松抗病虫害能力相对较强,保存率较高。在本试验中尚未观察到严重的病虫害,各个树种的保存率体现了该树种对台山环境的适应性。4个树种(变种)中4年平均保存率最低的是古巴加勒比松,只有44.76%,且3个古巴加勒比松种源的保存率非常接近,表明台山的某些因素限制了古巴加勒比松的生长。洪都拉斯加勒比松和巴哈马加勒比松的平均保存率都约为66%,但在两个变种内种源间保存率差异较大。在洪都拉斯加勒比松内,2号种源的保存率最低,其发芽率也是所有种源里最低的,表明该种源种子质量有待提高。
本试验共设置23个区组,方差分析结果显示在区组间各生长性状的差异达到极显著水平。在不同树种(变种)间,各生长性状的差异同样都达到极显著水平。不同年度的树高数据显示,1年生时巴哈马加勒比松在前期生长小于湿地松和洪都拉斯加勒比松,到4年生时成为最高的树种。在广西合浦、广东湛江、海南琼海的加勒比松种源试验中,连续多年的调查结果显示出一样的生长趋势,巴哈马加勒比松树高超过其他变种是在种植后3年[10-12]。在海南岛东部及西北部的种源试验结果显示加勒比松3个变种在不同立地条件下适应程度有明显差异,在立地条件较差的海南岛东部,洪都拉斯加勒比松的生长量最大,而在立地条件较好的西北部,巴哈马加勒比松和古巴加勒比松生长量都超过洪都拉斯加勒比松,尤其是树高,与地点的交互作用极显著,表明加勒比松控制树高的基因在不同立地条件下遗传性不稳定[10]。本研究只设置了一个试验地点,3个生长量性状在4年时的遗传力均超过0.8,各生长性状显示了较稳定的遗传,并且各个性状之间也表现出高度的遗传相关和表型相关,这表明在台山进行加勒比松种源试验并筛选优良种源的可靠性较高。不同种源的洪都拉斯加勒比松生长差异较大,其中产自澳大利亚的洪都拉斯加勒比松(12号种源)生长表现最好,其次是6、10号种源材料。所有种源中,产自巴西的巴哈马加勒比松16号种源生长表现最好,从国外引进的种子生产国里,巴西的纬度与台山最接近,两地气候更接近,可能是巴西种源材料在台山生长良好的因素之一。对洪都拉斯10个种源进行地理因子与生长量简单相关分析,结果显示纬度与生长量有中低等的相关系数,却并没有达到显著水平。
在加勒比松里,古巴加勒比松和巴哈马加勒比松分别有一个种批来自我国湛江市遂溪县,这两个种批4年保存率较低,均未达到改变种的平均水平,生长量也都未达到该变种的平均水平。而巴西种源的巴哈马加勒比松的材积则达到我国种子园种子的2.4倍。在我国加勒比松引进热潮期,为了解决加勒比松种子的长期供应不足的问题,广东省营建了11个种子园或母树林,总面积达117 hm2[19],然而这批种子园材料均选自我国首次引种的约3 000个个体[20-21],遗传基础很窄。随着时间推移,加勒比松改良热潮退去,广东省加勒比松种子园面积减少到58 hm2,种子园内保存的无性系数量也进一步减少,过窄的遗传基础可能导致了遂溪县生产的加勒比松种子品质下降,因此,进一步引种拓宽我国加勒比松遗传基础非常重要。
4 结论
本研究的3个加勒比松变种中,洪都拉斯加勒比松的种源数量较多,变异程度高,部分种源在台山生长表现优秀。来自巴西的巴哈马加勒比松材积增益是所有种源中最高的,而古巴加勒比松不适宜种植在台山,两个美国种源的湿地松在台山无明显增益。由于国内加勒比松遗传基础较窄,生产的加勒比松子代表现出一定的衰退,本次引种试验可有效拓宽我国加勒比松的遗传基础。在未来加勒比松引种中,应重点考虑原产的洪都拉斯加勒比松,以及巴西产的巴哈马加勒比松,定向筛选抗寒加勒比松,还应当尽量选择高纬度的种源。
猜你喜欢
现代经济信息(2023年17期)2023-09-04 10:42:13
美食(2022年2期)2022-04-20 14:34:19
中国果业信息(2020年8期)2020-12-16 11:28:26
少儿美术(快乐历史地理)(2019年5期)2019-09-10 08:34:36
小天使·一年级语数英综合(2019年8期)2019-08-27 02:23:00
世界热带农业信息(2018年8期)2018-03-27 01:10:38
世界热带农业信息(2016年1期)2016-04-13 12:37:20
发明与创新(2015年17期)2015-02-27 10:39:04
小资CHIC!ELEGANCE(2015年1期)2015-01-23 14:41:51
海外英语(2006年6期)2006-07-19 07:25:40
