
干旱胁迫对9个葡萄砧木品种生理指标的影响
2020-09-30 02:51,,,,,
经济林研究 2020年3期
, ,,, ,
(1.新疆农业大学 林学与园艺学院,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区玛纳斯县中信国安葡萄酒业有限公司,新疆 玛纳斯 832200)
葡萄VitisviniferaL.为葡萄科Vitaceae 葡萄属Vitis的一种落叶木质藤本果树[1],其果实酸甜爽口,营养丰富,在全球果树生产中占据重要地位,其栽植面积和产量曾长期均居世界水果栽植面积和产量的首位。新疆维吾尔自治区则是我国葡萄栽培面积最大的省区,因为气候条件及地形特点等因素的影响,其水资源极度缺乏,是我国水资源匮乏的干旱地区之一,干旱严重制约了新疆葡萄产业的健康发展。针对水资源匮乏地区的果树生产,国外相关研究多集中于筛选抗旱砧木方面。许多研究结果都表明,葡萄砧木对接穗的树体生长势、果实成熟期、果实产量与品质及抗性等均有较大影响,合理使用砧木不仅可以提高接穗的抗逆性,扩大优良品种的种植范围,而且能最大限度地展现其品种特性[2-4]。因此,研究并筛选抗旱型葡萄砧木对于促进新疆葡萄产业的发展具有重要现实意义。葡萄砧木抗旱能力的评价是一个非常重要的问题,目前国内外主要通过测定其生理指标来进行评价。郭修武等人[5]分别以‘无核白鸡心’和‘巨峰’葡萄为研究对象,在相同的干旱胁迫条件下,通过测定株高、叶片光合速率、水分利用效率、超氧化物歧化酶活性和脯氨酸含量,得出了各项指标都较高的‘巨峰’葡萄其抗旱能力强于‘无核白鸡心’葡萄的结论;薛加楠等人[6]在比较分析3 种不同地理起源葡萄的抗旱性时发现,叶片的脯氨酸、过氧化物酶、过氧化氢酶和超氧化物歧化酶等指标含量均最高的品种,其抗旱能力最好;邹原东等人[7]以6 个葡萄品种为试材进行了干旱胁迫试验,结果发现,在干旱胁迫条件下,可溶性蛋白、可溶性糖和脯氨酸含量的增加幅度均较大而丙二醛含量上升较缓慢的品种,其对干旱胁迫的耐受力较强;胡宏远等人[8]以赤霞珠为试验材料进行了不同程度的干旱胁迫试验,结果得出,干旱胁迫可使其丙二醛、脯氨酸和可溶性糖的含量均有增加,使其过氧化氢酶活性增强,而其可溶性蛋白含量呈现出先降后升的变化趋势,其过氧化物酶活性则呈先升后降的“抛物线”变化趋势,其超氧化物歧化酶活性却呈现出“双峰”变化趋势。目前,我国研究者对葡萄抗旱性的研究多集中于鲜食葡萄品种和酿酒葡萄品种,而针对葡萄专用抗旱砧木筛选的研究报道比较少。近年来,新疆引进了大量的葡萄专用砧木品种,但由于对其适应性的了解不够,使得各种砧木资源不能很好地发挥其作用,面对日趋紧张的水资源短缺现状,迫切需要从现有的葡萄砧木品种中筛选出抗旱能力强的砧木,更好地解决干旱区绿洲葡萄生产的问题。为给新疆优良抗旱砧木的筛选与推广提供科学依据,本研究以9个葡萄砧木品种为试材,在人工控水给予干旱胁迫的条件下进行了盆栽试验,对砧木抗旱性评价的相关生理指标进行了测定,并采用主成分分析法对各个砧木的抗旱能力进行了综合评价,现将研究结果分析报道如下。
1 材料与方法
1.1 试验材料
本研究所用的葡萄砧木主要来源于新疆维吾尔自治区玛纳斯河流域的中信国安葡萄酒业有限公司,砧木品种分别为山河3号、山河4号、河岸4号、河岸7号、河岸9号、河岸10号、1613、101、Ganzia,共计9 个葡萄砧木品种。
1.2 试验方法
1.2.1 试验设计
试验在新疆农业大学特色果树研究中心进行。于2018年12月采集各品种1年生休眠枝条,每个品种各剪取10 ~15 根枝条,剪取的枝条长为50 ~80 cm,枝条取回后沙藏。于2019年3月10日对9 个葡萄砧木品种进行扦插育苗试验,每个品种10 盆,每盆1 株,栽于口径30 cm、高40 cm的塑料盆中,扦插基质配比为营养土∶珍珠岩∶细砂=2∶1∶1,进行遮阴管理,待苗木恢复正常生长后,进行统一的水分管理。
2019年7月将已有的葡萄砧木扦插苗培养至每株长出7 ~8 片叶后,每个品种各选取叶片数、枝长和长势均基本一致、生长状态良好且无病虫害的扦插苗3盆,进行人工控水给予干旱胁迫处理。浇透水后第2 天进行第1 次取样,之后每间隔5 d采样1 次,直到第21 天顶叶出现萎蔫状时停止取样,取样次数共计5 次,取样时间为上午9:00—11:00 时,每次每盆采取健康的功能叶片3 片,每个砧木品种共采9 片叶,混合后将其分作3 份,作为3 个重复。并将叶片置于液氮中保存1 h,然后放在-25 ℃的冷藏冰箱中保存以备用。
1.2.2 生理指标的测定
采用饱和称重法[9]测定叶片相对含水量(relative water content,RWC),采用电导仪法[10]测定相对电导率(relative conductivity,REC),参照上海酶联生物公司相关试剂盒中说明的测定方法分别测定超氧化物歧化酶(superoxide dismutase,SOD)活性、过氧化物酶(peroxidase,POD)活性和脯氨酸(proline,Pro)含量、丙二醛(malondialdehyde,MDA)含量。
1.3 数据处理
本试验所有数据处理和表格制作均采用Excel 2010 和SPSS 19.0 完成。
2 结果与分析
2.1 干旱胁迫下不同砧木叶片相对含水量的变化
叶片相对含水量指的是组织含水量占饱和含水量的百分数,是评价植物叶片保水力的重要指标之一。不同时间的干旱胁迫下9 个葡萄砧木品种叶片相对含水量见表1。表1表明,在干旱胁迫处理期间,9 个葡萄砧木品种叶片相对含水量随着干旱胁迫时间的延长均呈现出明显的下降趋势。在浇透水后第2 天,所有测试葡萄砧木品种的叶片相对含水量都保持为91%~85%,没有显著性差异(P>0.05)。但从胁迫的第6 天开始,各砧木品种间叶片相对含水量开始出现显著性差异(P<0.05),至浇透水后第21 天,其差异达到最大,仅有河岸9号和山河4号这2 个品种的叶片相对含水量大于65%,其中山河4号的最高,为65.45%;叶片含水量较低的品种分别为河岸10号、101、Ganzia、山河3号和河岸4号,其叶片相对含水量为60%~45%;而河岸7号和1613这2 个品种的叶片相对含水量均低于45%。与浇透水后第2 天的相比,河岸9号和山河4号的降幅最小,分别为23.23%和24.24%;而1613 和河岸7号的降幅均大,分别47.16%和44.62%。

表1 不同时间的干旱胁迫下9 个葡萄砧木品种叶片相对含水量的变化情况†Table1 The changes of relative water content in leaves of nine grape rootstocks varieties under different drought stress time
2.2 干旱胁迫下不同砧木叶片细胞质膜透性变化
相对电导率是观测植物细胞膜状况的一个重要生理指标,逆境条件会造成膜蛋白损伤,从而引起胞液外渗,电导率也随之增加。一般认为,相对电导率越高,其上升幅度越大,则表明细胞膜透性越差,植物受到伤害的程度越高,其抗旱能力越差。不同时间的干旱胁迫下9 个葡萄砧木品种叶片的相对电导率见表2。由表2可知,干旱胁迫下各砧木品种叶片的相对电导率始终存在显著性差异(P<0.05),且随着干旱胁迫时间的延长,所有葡萄砧木品种叶片的细胞质膜透性不断增大,细胞膜遭到了不同程度的伤害。在干旱胁迫的11 d内,各品种叶片的相对电导率均处于缓慢上升阶段;在干旱胁迫的11 ~21 d 内,9 个葡萄砧木品种叶片的相对电导率均出现了明显的增加趋势。与胁迫第11 天的叶片相对电导率相比,胁迫第21 天的叶片相对电导率上升了38.56%~24.45%;与胁迫第2 天的叶片相对电导率相比,胁迫第21 天的叶片相对电导率上升了51.44%~39.93%。9 个葡萄砧木品种中,河岸9号的叶片相对电导率最低,为49.06%;1613 的叶片相对电导率最高,为64.27%;二者相差15.21%。

表2 不同时间的干旱胁迫下9 个葡萄砧木品种叶片相对电导率的变化情况Table2 The changes of leaf relative conductivity of nine grape rootstocks varieties under different drought stress time
2.3 干旱胁迫下不同砧木叶片脯氨酸含量的变化
渗透调节是植物受到干旱胁迫后主动适应环境变化的重要调节机制之一,而脯氨酸(Pro)作为该机制中最重要的调节物质之一,其含量的多少在一定程度上可以显示出植物在缺水情况下渗透调节能力的大小,进而反映出植物抗旱能力的强弱。不同时间的干旱胁迫下9 个葡萄砧木品种叶片中的Pro 含量见表3。从表3中可以看出,在干旱胁迫条件下,所有测试葡萄砧木品种叶片中的Pro含量均呈上升趋势。胁迫前期,各葡萄砧木品种之间其含量无显著性差异(P>0.05);从胁迫的第11天开始,其含量出现了显著性差异(P<0.05)。整个试验期间,河岸9号、101、山河3号和河岸4号这4 个砧木品种叶片中的Pro 含量均呈“慢—慢—慢—快”的上升趋势,而河岸10号、河岸7号、Ganzia、山河4号和1613 叶片中的Pro 含量则均呈“慢—快—慢—快”的上升趋势。所有砧木品种叶片中的Pro 含量随着干旱胁迫时间的延长都呈现出大幅度的增加趋势,其中,河岸9号、101 与河岸4号的表现均较为明显,与胁迫第11 天的相比,其叶片中的Pro 含量分别增加了1.54、1.41 和1.39 倍。9 个葡萄砧木品种叶片Pro 含量增幅的大小顺序为:河岸9号>河岸10号>101 >Ganzia >山河3号>山河4号>河岸7号>河岸4号>1613。其中,与胁迫第2 天的叶片Pro 含量相比,含量增加幅度最大的品种为河岸9号,其含量增加了2.26倍;含量增加幅度最小的品种为1613,其含量仅增加了1.80 倍。

表3 不同时间的干旱胁迫下9 个葡萄砧木品种叶片脯氨酸含量的变化情况Table3 The changes of proline content in leaves of nine grape rootstocks under different drought stress time
2.4 干旱胁迫下不同砧木叶片丙二醛含量的变化
干旱胁迫可以诱导植物产生大量的活性氧,引起膜脂过氧化反应,而丙二醛(MDA)作为该反应的最终分解产物,其累积量通常被用来表示逆境对植株的伤害程度。不同时间的干旱胁迫下9 个葡萄砧木品种叶片中的MDA 含量见表4。表4表明,9 个葡萄砧木品种叶片中的MDA 含量随着干旱胁迫程度的加重均明显增加。在干旱胁迫的6 d内,9 个砧木品种叶片中的MDA 含量没有显著性差异(P>0.05);随着胁迫程度的加重,所有测试品种之间叶片中的MDA 含量,从胁迫的第11天开始其差异显著(P<0.05),且其含量均呈现出明显的上升趋势,其中增幅较小的品种有河岸9号和101,其次分别是河岸10号、Ganzia 和山河4号,而山河3号、河岸4号、1613 和河岸7号这4 个品种叶片MDA 含量的增加均较明显;至干旱胁迫的第21 天,9 个葡萄砧木品种叶片中的MDA 含量的大小顺序为:河岸7号>1613 >河岸4号>山河3号>山河4号>Ganzia >河岸10号>101 >河岸9号。

表4 不同时间的干旱胁迫下9 个葡萄砧木品种叶片丙二醛含量的变化情况Table4 The changes of MDA content in leaves of nine grape rootstocks varieties under different drought stress time
2.5 干旱胁迫下不同砧木叶片超氧化物歧化酶活性的变化
超氧化物歧化酶(SOD)是植物体内的重要保护酶之一,作为植物体内清除活性氧系统的第一道防线,它在酶促活性氧自由基清除系统中发挥着至关重要的作用。不同时间的干旱胁迫下9 个葡萄砧木品种的叶片SOD 活性见表5。表5表明,在干旱胁迫期间,所有葡萄砧木品种的叶片SOD活性始终存在显著性差异(P<0.05),且总体上其SOD 活性均有所增强,但其增强趋势因品种不同而有所不同。其中,101、河岸9号、河岸10号、山河4号和山河3号这5 个品种的叶片SOD 活性在整个胁迫期间的变化曲线均为“单S”曲线;而河岸7号、Ganzia、河岸4号和1613 这4 个品种叶片SOD 活性的变化曲线均呈缓慢升高—快速下降—快速升高—较快下降的“双S”曲线,且其SOD 活性于胁迫的第15 天均达到最大值。
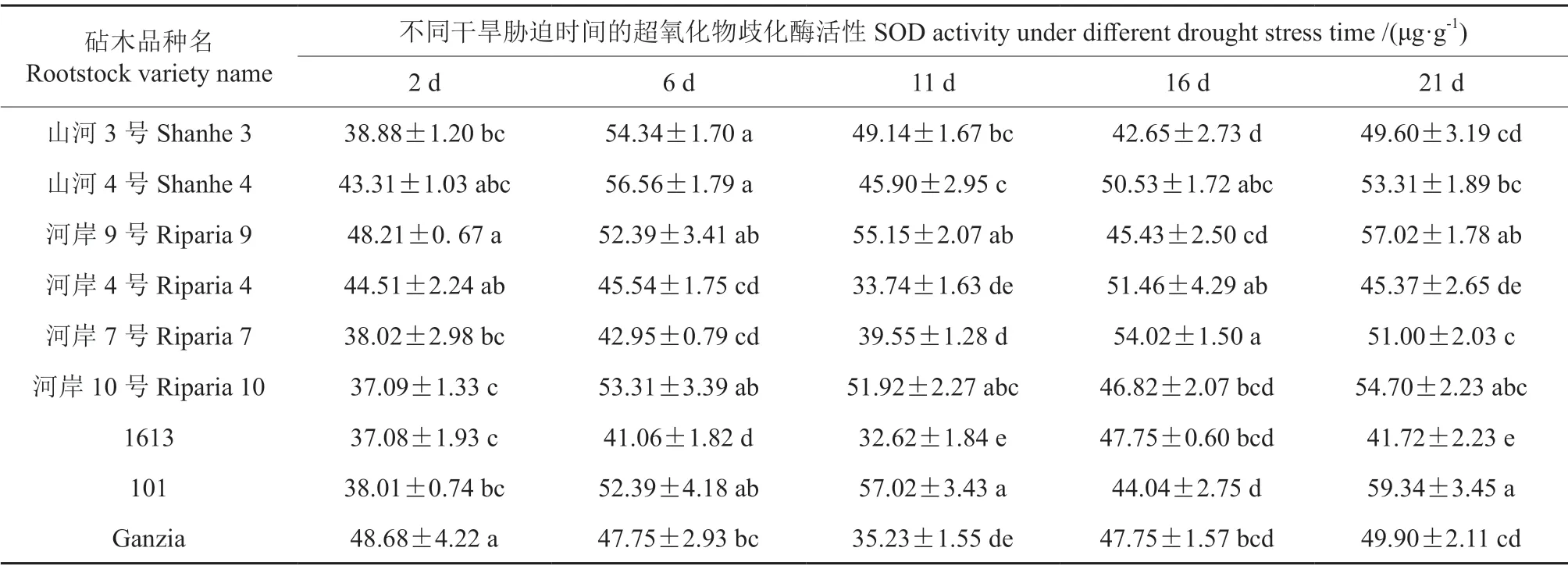
表5 不同时间的干旱胁迫下9 个葡萄砧木品种叶片超氧化物歧化酶活性的变化情况Table5 The changes of SOD activity in leaves of nine grape rootstocks varieties under different drought stress time
2.6 干旱胁迫下不同砧木叶片过氧化物酶活性的变化
过氧化物酶(POD)广泛存在于植物体内,是除SOD 外另一种具有较高活性的保护酶,其主要功能是催化以H2O2为氧化剂的氧化还原反应,保护植物的细胞膜系统。不同时间的干旱胁迫下9 个葡萄砧木品种叶片过氧化物酶活性的变化情况见表6。从表6中可以看出,在干旱胁迫条件下,9 个葡萄砧木品种间叶片POD 活性的差异始终显著(P<0.05),且随着胁迫时间的延长其活性均呈“抛物线”的变化趋势。至浇透水后的第11 天,所有砧木品种叶片POD 的活性均出现缓慢升高而后迅速上升的变化趋势;随着胁迫程度的加重,河岸7号和1613 叶片POD 的活性均开始降低,其余7 个砧木品种叶片POD 的活性均继续以较快速度上升,但所有砧木品种叶片POD 的活性在胁迫的第21 天均大幅度下降。
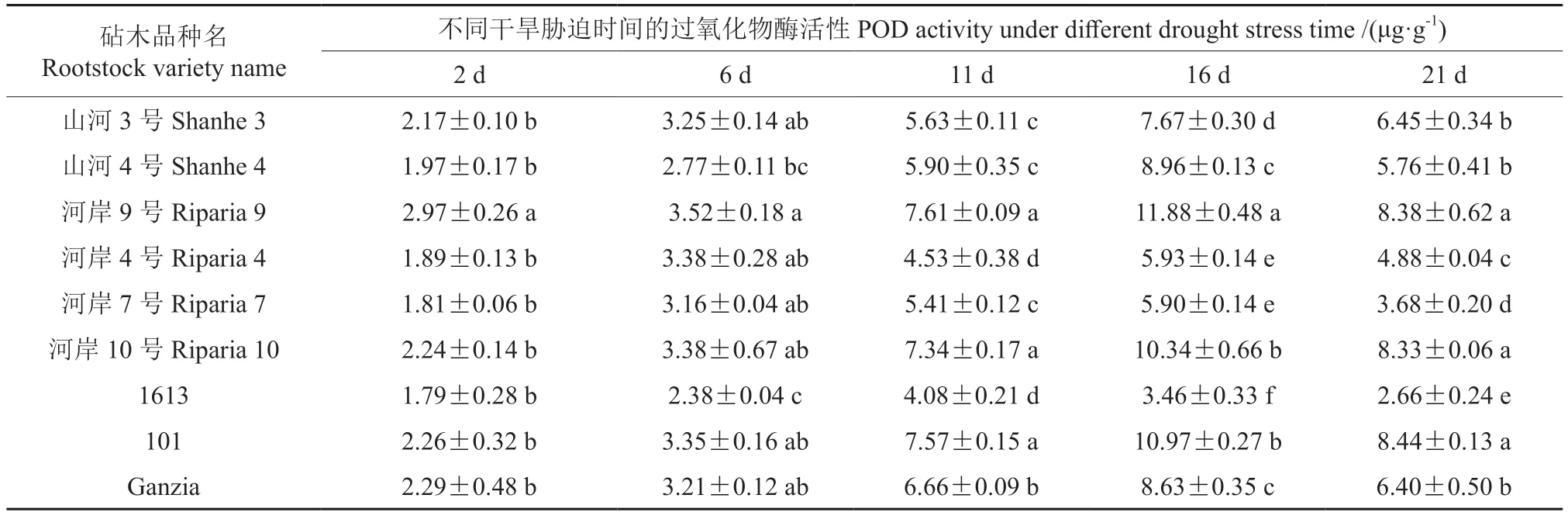
表6 不同时间的干旱胁迫下9 个葡萄砧木品种叶片过氧化物酶活性的变化情况Table6 The changes of POD activity in leaves of nine grape rootstocks varieties under different drought stress time
2.7 干旱胁迫下9 个葡萄砧木品种生理指标的主成分分析
9 个葡萄砧木品种在相同的干旱胁迫条件下表现出的抗旱能力各不相同,为了更好地评价其抗旱能力,采用主成分分析法对上述6 个抗旱生理指标进行了综合分析。通过分析得到了各测定指标主成分的特征值、贡献率和累计贡献率(表7)。根据主成分得分矩阵和标准化后的数据分别计算出主成分1(F1)、主成分2(F2)、主成分3(F3)、主成分4(F4)、主成分5(F5)和主成分6(F6)的表达式,再以表7中6 个主成分的特征值与所提取主成分总特征值之和的比值作为权重,计算得出主成分的综合模型,据此得到表8。6 个主成分的表达式和主成分的综合模型分别如下:
F1=-0.232X1+0.242X2+0.230X3+0.222X4+0.097X5+0.162X6;
F2=0.280X1-0.110X2+0.103X3-0.329X4+0.730X5+0.433X6;
F3=0.108X1-0.056X2+0.145X3-0.321X4-0.771X5+0.938X6;
F4=0.571X1+0.190X2+1.849X3-0.777X4-0.298X5-0.850X6;
F5=2.100X1+2.978X2-0.974X3+0.057X4-0.171X5-0.035X6;
F6=2.787X1-1.718X2+0.974X3+3.490X4-0.175X5+0.479X6;
F=0.667X1+0.179X2+0.103X3+0.032X4+0.012X5+0.007X6。
上列各式中:X1表示平均相对含水量;X2表示平均相对电导率;X3表示平均脯氨酸含量,X4表示平均丙二醛含量;X5表示平均SOD 活性,X6表示平均POD 活性;F代表主成分的综合得分。
分析表7中各个测定指标主成分的累积贡献率可以发现,在6 个主成分中,前2 个主成分的累计贡献率达到84.594%,这表明前2 个主成分代表了原有6 个主成分的84.594%的信息,因此可以分别用这2 个主成分对葡萄砧木品种的抗旱能力进行综合分析。其中,主成分1 的贡献率是最高的,为66.660%,说明其反映的信息量最大,主成分1 中除了相对含水量外,其余指标都是正向载荷的,在主成分1 中相对电导率、脯氨酸含量和丙二醛含量都有较高的载荷,说明主成分1 能基本反映这些指标的信息。结合表8和前2 个主成分的表达式分析后可以得知:在主成分1 的表达式中,载荷最高的是平均相对电导率(X2),表明主成分1 主要反映的是相对电导率的信息,其中得分最高的砧木品种是河岸7号,其在主成分1 中的得分为17.300 分,次高分为河岸9号,其得分为16.878 分,得分最低的砧木品种为河岸4号,其得分为15.314 分;主成分2 的贡献率为17.934%,6 个指标在主成分2 中载荷最高的是平均SOD 活性(X5),表明主成分2 主要反映的是SOD 活性因子的信息,得分最高的砧木品种是河岸9号,其在主成分2 中的得分为44.153 分,次高分为101,其得分为42.062 分,得分最低的品种则为1613,其得分为31.366 分。
根据表8的排名可知,9 个葡萄砧木品种抗旱能力的强弱顺序为:河岸9号>山河4号>河岸10号>101 >山河3号>河岸7号>Ganzia >1613 >河岸4号。在9 个葡萄砧木品种中,河岸4号的综合得分最低,只有0.528 分,表明其抗旱能力最差;而河岸9号的综合得分最高,达到了0.601 分,表明其抗旱能力最强。

表7 各测定指标主成分的特征值、贡献率和累计贡献率Table7 The feature value,contribution rate and accumulative contribution rate of principal component of detection indexes

表8 主成分综合评价结果Table8 Results of comprehensively principal component evaluation
3 讨 论
3.1 干旱胁迫下叶片相对含水量的变化
叶片相对含水量是叶片组织水质量占饱和组织水质量的比率,常用于衡量植物受到干旱胁迫后叶片水分的亏缺状况,它比普通的叶片含水量能更敏感地突显植物水分状况的变化情况,在一定程度上可以反映出植物的保水持水能力,与植物的抗旱性呈正相关[7,11-12]。研究中发现,在干旱胁迫期间,9 个葡萄砧木品种的叶片相对含水量随着胁迫时间的延长均逐渐降低,其降幅为23%~45%,表示葡萄砧木在此期间水分丧失较为严重。在干旱胁迫条件下,河岸9号可以维持较高的叶片相对含水量,表明其体内水分亏缺程度较小,可以保持体内水分平衡,能为其各种代谢活动的正常进行提供保障,拥有较强的抗旱能力[13]。这一试验结果与高鹏等[14]对4 个葡萄品种抗旱性的研究结果一致。高鹏等[14]在研究中也得出,各葡萄品种在受到干旱胁迫时,叶片相对含水量均有不同程度的下降,而叶片相对含水量较高的品种,其有较强的保水力。
3.2 干旱胁迫下叶片相对电导率与丙二醛含量的变化
干旱胁迫会引起植物细胞膜过氧化,导致细胞膜的选择透性发生改变甚至丧失,造成胞内物质渗出,组织浸出液电导率增加[15]。同时,丙二醛作为膜脂过氧化分解的重要产物,其产生后会进一步加重对膜的伤害,丙二醛的含量可以代表膜脂过氧化的程度,进而反映细胞膜质受伤害的程度[16]。本试验结果表明,随着干旱胁迫程度的加重,所有测试葡萄砧木品种的叶片相对电导率和丙二醛含量都呈现上升趋势,说明在持续的干旱胁迫条件下,叶片组织的保护能力逐渐衰退,细胞膜的结构和功能均遭到不同程度的破坏,透性变大,脂膜过氧化严重受损[17]。而在9 种受试砧木中,河岸7号的叶片相对电导率和丙二醛含量的增幅均最大,表明其抗旱能力较弱;而河岸9号的叶片相对电导率和丙二醛含量的增幅均最小,表明其抗旱能力均较强。宋海鹏等[18]在研究5 种景天属植物时也得出,随着干旱胁迫程度的加重,植物的电导率和MDA 含量均呈递增趋势,且增幅最大的植物其抗旱能力最弱。
3.3 干旱胁迫下叶片脯氨酸的含量变化
脯氨酸具有偶极性,其两端分别与水分子和蛋白质相连接,蛋白质可以通过脯氨酸束缚更多的水分,所以脯氨酸含量在一定程度上可以反映出植物体内缺水状况[19]。试验中发现,9 个葡萄砧木品种叶片中的脯氨酸含量随着干旱胁迫时间的延长均呈现出较为明显的上升趋势。在此条件下,植株通过渗透调节维持细胞内外的渗透平衡,减少水分散失,保护质膜的完整性,降低干旱胁迫带给植物的伤害[20-22]。其中又以河岸9号叶片中脯氨酸含量的增幅为最大,而1613 叶片中脯氨酸含量的增幅最小。黄桂华等[23]和祁伟亮等[24]分别对柚木无性系和5 种桑树品种的抗旱性进行了试验研究,他们得出,植物叶片中的脯氨酸含量会随着干旱胁迫程度的加重而呈明显的上升趋势,且脯氨酸含量与品种的抗旱性呈正相关。本试验所得结果与他们研究得出的结果具有一致性。
3.4 干旱胁迫下叶片保护酶的活性变化
保护酶活性在一定程度上决定了植物的抗旱性[21],可以作为葡萄抗旱性评价的鉴定指标[25]。SOD 和POD 是目前葡萄抗氧化物质中研究最多的两种酶[26],在植物受到干旱胁迫后二者协同作用。本研究发现,所有测试葡萄砧木品种的SOD 活性在前期都呈现先升高后下降的变化趋势,分析认为,各砧木品种前期受到的轻度或短期的干旱胁迫导致其体内SOD 活性的增加,降低了膜脂过氧化作用对植物造成的伤害,保护了酶系统。随着胁迫时间的延长,9 个品种中有5 个品种出现了SOD 活性再次升高的现象,SOD 在清除超氧自由基、过氧化氢和过氧化物及减少羟基自由基形成等方面都起着重要作用,推测认为,SOD 活性再次升高是细胞抵御活性氧伤害的膜保护系统再次启动,从而减轻因胁迫产生的自由基对细胞膜系统的伤害。关于这种现象,张青等[27]对七叶树幼苗、肖涵等[28]对3 种宿根花卉的抗旱性研究时也都得出了同样的论断。而Fazeli 等[29]通过研究也指出,干旱胁迫程度的加重,会导致植物叶片中SOD 活性的增加,而变化越大的植物,其抗旱性越强;反之抗旱性较弱。
POD 是植物体内重要的抗氧化保护酶,SOD与POD 活性共同作用于活性氧的消耗,植物在受到干旱胁迫时,活性氧的消耗方面SOD 通常起主要作用,而POD 起辅助作用;当干旱胁迫程度进一步加重,SOD 活性达到较稳定的状态,转而由POD 起主要作用[30]。靳月等[31]在研究干旱胁迫对闽楠幼苗的生长和生理影响时发现,SOD 和POD活性的变化趋势相似,整体均呈先上升后下降的趋势,具有协同性。而本研究中所有砧木品种的POD 活性随着干旱胁迫程度的加重均呈现先升高后下降的变化趋势,并没有出现和SOD 相似的变化趋势,其主要原因是,干旱胁迫前期为了阻止过氧化作用带给植物的伤害,维持植物正常的生理活动,帮助其抵御逆境,POD 一直呈现增加趋势。干旱胁迫后期植物体内氧代谢失调,活性氧的生成速度加快,而抗氧化系统清除活性氧的功能降低,使得活性氧大量积累,加重了膜脂过氧化程度,超过植物的承受能力,膜的完整性遭到了破坏,进而POD 活性也随之下降[30,32-33]。冀鹏飞等[34]在对葡萄根系、杨义伶等[35]在对柑橘砧木的研究中也都得出,POD 活性随着干旱胁迫的增强呈现出先上升后下降的趋势,且抗旱性强的品种其POD活性维持在较高的水平上。本试验所得结果与其具有一致性。
3.5 本研究存在的局限性与下一步研究的方向
在自然条件下生长的植物由于经历了长时间的温度、水分等变化,自身各方面发生了不同程度的改变。本研究仅采用盆栽苗进行试验,不能完全地反映出各砧木品种在田间的抗旱能力,且植物的抗旱力受到遗传基因、起源地、生长环境和植株生长发育状态诸多因素的影响,本研究仅对特定环境下部分生理指标的变化进行了探讨。因此,若要综合评价品种的抗旱能力尚需进一步结合大田栽培环境下各个砧木品种的表型、叶片解剖结构、生长发育动态、光合特性甚至是砧穗组合的变化来评判,以便更加科学全面地评价各砧木品种的抗旱性,为实际生产提供更加可靠的理论依据。
4 结 论
植物对干旱的应答反应是多种因素相互作用所产生的结果,单一指标不能准确反映出其抗旱性。在人工控水给予干旱胁迫的条件下,随着干旱胁迫时间的延长,9 个葡萄砧木品种表现出不同的抗旱性,通过测定叶片中的相对含水量、相对电导率、脯氨酸含量、丙二醛含量、SOD 活性和POD 活性等生理指标,结合主成分分析法综合评价其抗旱能力得出,各个砧木品种抗旱能力的大小顺序为:河岸9号>山河4号>河岸10号>101 >山河3号>河岸7号>Ganzia >1613 >河岸4号。
猜你喜欢
中老年保健(2022年3期)2022-11-21
今日农业(2021年14期)2021-11-25
落叶果树(2021年6期)2021-02-12
今日农业(2020年13期)2020-12-15
棉花学报(2020年3期)2020-08-08
河南水利年鉴(2020年0期)2020-06-09
河北果树(2020年2期)2020-05-25
科学与财富(2019年15期)2019-10-21
建材发展导向(2019年11期)2019-08-24
现代园艺(2017年21期)2018-01-03
