
生境梯度影响下的天然红松种群空间格局与种内关联
2020-09-17 00:01:14孙志虎谷会岩陈祥伟
生态学报 2020年15期
董 雪,杜 昕,孙志虎,谷会岩,陈祥伟
东北林业大学林学院, 哈尔滨 150040
植物种、植物群落及植被景观有其自身的空间分布规律,称为广义空间分布格局;植物种群在群落中的非随机分布称为种群的分布格局,即狭义格局,其是植物群落斑块与植物景观斑块形成的基础[1-2]。种群格局分析的实质是研究单个或多个种群在群落内部的镶嵌结构,进而揭示空间格局的形成原因。分析种群分布格局有助于认识种群的生物学特性、生态学过程及物种与环境因子间的关联性。种群的空间格局对研究尺度具有很强的依赖性[3-4]。而相对于传统格局分析方法,空间点格局分析能够量化植物种群在不同尺度上的空间分布格局与种内、种间联结关系;其以个体空间坐标为基础,在分析过程中最大限度的利用二维空间点信息,具有较强的检验效力[5-9]。在点格局分析基础上演伸出的标记点格局[10-11]及重复采样点格局分析[12-15]等方法,可以更详尽的探索不同空间尺度上的生态学问题。种群空间格局的形成受到物种自身生物学特性与群落环境的共同影响。不同生境中相同物种的空间分布特征存在差异性[16-17],而明确尺度的点格局分析可以清晰的揭示不同生境中的种群分布状态与生态学过程[9,18-19]。
红松是第四纪冰川孑遗种,以其为建群种的阔叶红松林长期存在于东北亚地区,是该区的山地地带性顶级植被,具有复杂的动植物成分[20]与系统稳定性。建群种红松(Pinuskoraiensis)具有较宽的超体积生态位,在不同的环境下可与不同树种混交,构成不同林型原始红松林。曾有学者探讨过地形因子对原始红松林型的影响,认为阔叶红松林林型的变化主要是由坡位、坡度等地形因子变化,进而造成土壤水分条件变动引起的。在同一分水岭内随着坡位下降,坡度不断减小,土壤水分形成干燥、潮湿、湿润、重湿润的环境梯度,并产生与之对应的红松林林型。基于此,许多学者对阔叶红松林林型组与林型进行了划分,如徐化成[21]将典型阔叶红松林划分为3个林型组,6个林型,其中高海拔林型组包括高海拔藓类云冷杉红松林;坡地林型组包括细叶苔草蒙古栎红松林、毛榛紫椴红松林、灌木枫桦红松林;谷地林型组包括蕨类云冷杉红松林和春榆水曲柳红松林。李文华[22]将阔叶红松林划分为谷地潮湿红松林型组、缓坡潮湿红松林型组、斜坡半湿润红松林型组及陡坡半干旱红松林型组。
近年,已有较多研究者应用空间点格局分析的方法对阔叶红松林中立木空间分布规律进行了探索,如刘妍妍[23-24]与侯红亚和王立海[25]分别基于大样地调查数据分析了典型红松林中主要乔木树种空间分布;夏富才[26]研究了南部阔叶红松林与不同演替阶段次生林中林木空间分布格局等。目前,研究多以阔叶红松林或特定林型中各乔木树种为研究对象,探讨树种的生物学特性与生长过程。然而需要指出的是,不同生境中的相同植物种群在其定植、生长的过程中所经历的生态学过程必然有所差异,生境梯度下阔叶红松林中各树种的空间分布规律有待进一步探索。通过分析处于不同生境的原始红松林中乔木空间分布格局的差异,加深对影响林木定植、生长的生态学过程的理解,将有利于阔叶红松林的经营管理与维持红松林生态系统的长期稳定。基于此,本研究参考李文华对不同原始红松林型组的界定,选取7块分别位于4种不同坡位的原始红松林小样地,以建群种红松为研究对象,应用重复采样点格局分析的方法研究生境梯度作用下不同林层红松的空间分布格局,并回答以下科学问题:①随着坡位升高、坡度增大、典型红松林各林层中红松种群空间分布格局有何变化规律?②生境梯度影响下典型红松林中红松种群种内联结规律有何变化趋势?
1 研究区概况及研究方法
1.1 研究区概况
研究区位于黑龙江省凉水国家级自然保护区(47°10′55″N,128°53′20″E),属小兴安岭南坡。研究区气候类型为温带大陆季风气候,年平均气温-0.3℃,年平均最高气温7.5℃,年平均最低气温-6.6℃,年平均降水量676 mm,全年降雨多集中于6—8月份,有效积温2200—2600℃,无霜期100—120 d。原生植被类型是以典型阔叶红松林为代表的温性针阔混交林。目前,在逆行演替和进展演替共同作用,形成了以不同演替阶段的次生林为基底,以人工林及原始林为斑块的镶嵌分布的森林景观。地带性土壤为暗棕壤,非地带性土壤有草甸土、泽土、泥炭土、棕色森林土、白浆土等。
1.2 样地布设与调查
在凉水自然保护区19林班内,参照李文华[22]对红松林型生态系列的界定,综合林木组成、林下植被、坡位、坡度以及土壤水分等信息,在由坡底到坡顶的生境梯度上平行等高线设置7块50 m×50 m的样地[27],各样地基本信息见表1。样地调查采用相邻格子法,将调查样地划分为100个5 m×5 m的调查单元,调查并记录全部株高大于10 cm的乔木树种在样地中的相对位置坐标与株高。综合考虑徐化成[21]、李景文[28]、安慧君和张韬[29]对原始红松林林层的划分,并参考行业标准(LY:1572-2000)[30]中更新层的划定,将原始红松林林层划分为主林层(H≥16 m)、亚林层(10 m≤H<16 m)、演替层(1.5 m≤H<10 m)与更新层(0.1 m≤H<1.5 m)。依据调查乔木株高,判定乔木所处林层,以待进一步分析。各样地中,土壤取样按五点法进行,去除土壤表面杂草、枯落物与腐殖质后,挖开表土,在5—10 cm土层取土样,用于土壤含水率分析。
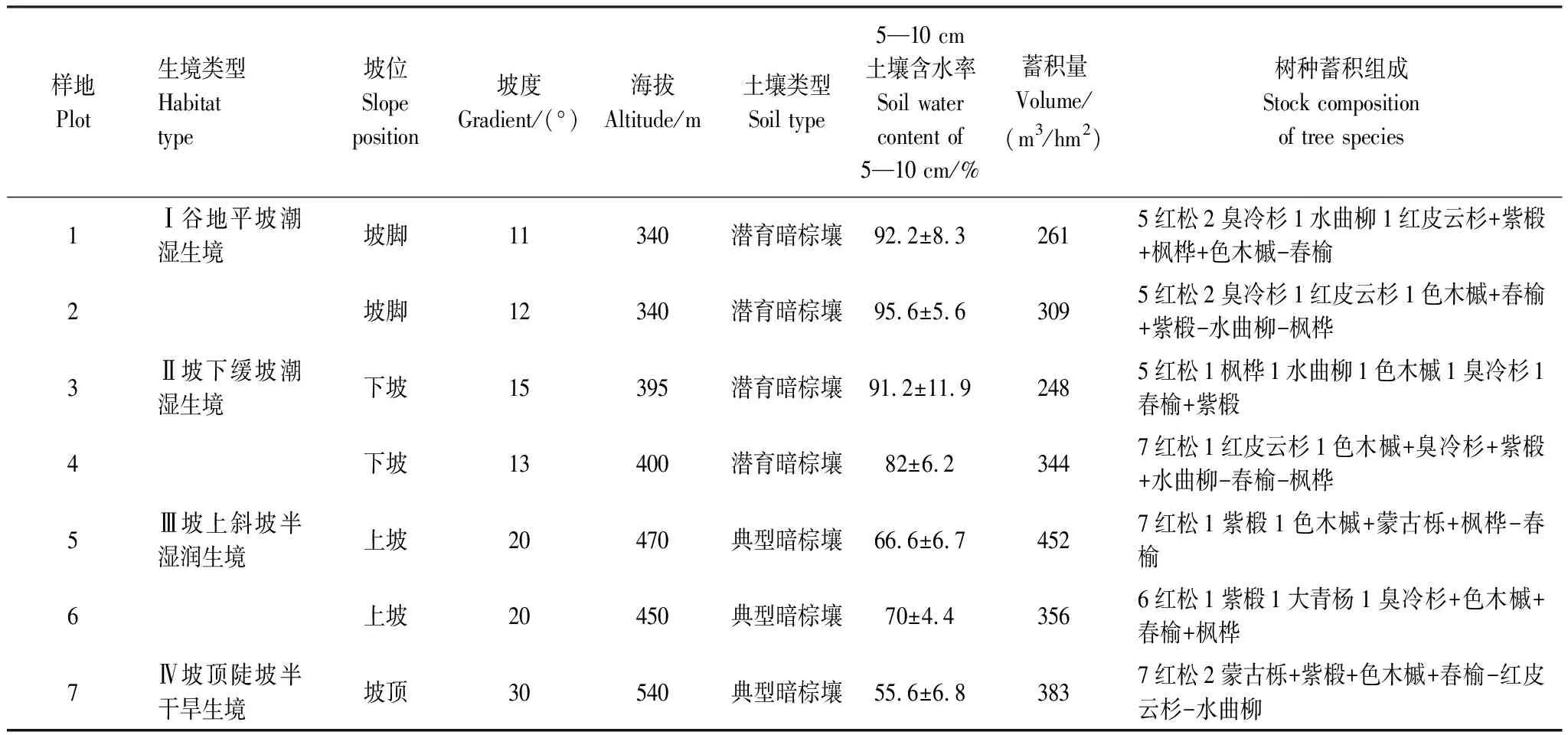
表1 红松原始林各样地基本信息表
1.3 点格局分析与重复采样点格局分析
点格局分析(Point pattern analysis)是以植物个体空间坐标为基础,分析不同尺度下种群格局与种间关系的分析方法[5-9]。而重复采样点格局分析(Replicate pattern analysis),是基于多个小样本空间的空间分析方法[12-15]。相对于依赖单一大样本的点格局分析,重复采样点格局分析可以增强试验结果的普适性、提高结果的可靠程度。
本研究采用O-ring(以下简称Or)函数为基本函数,进行点格局分析与重复采样点格局分析[31]。Or函数基于Ripely′s K函数与Mark相关函数,通过计算点外特定距离上的发生频率确定空间格局[3]。相对于Ripely′s K函数与Ripely′s L函数,以特定宽度环为统计基础的Or函数可以消除小尺度上的尺度累积效应[32-33]。
1.4 生境间种群空间分布规律比较
参考Pamela等[18]与Wang等[19]的方法,进行不同生境中红松种群空间分布规律的比较。该方法通过Or函数计算值、蒙特卡洛(Monte Carlo)法构建的包络线(envelop)及完全空间随机(Complete spatial random,CSR)假设下Or函数的理论值(Theoretical value,Tv)间关系计算不同生境中种群空间格局的无偏估计y,对比不同生境的种群空间分布格局。
如果Or>99%置信区间上限(CL+),则y=(Or-CL+)/|CL+-Tv|>0
如果Or<99%置信区间上限(CL+),且Or>99%置信区间下限(CL-),则y=0
如果Or<99%置信区间下限(CL-),则y=(Or-CL-)/|CL--Tv|-<0
对单一种群(林层)进行重复采样点格局分析,完全空间随机假设的理论值Tv为:
研究两种群(林层)间空间关系时,完全空间随机假设的理论值Tv为:
式中,j为重复采样次数;Areaj为第j次采样的样地面积;nj为第j次采样时种群一中个体数量;mj为第j次采样时种群二中个体数量。
研究中Or函数环宽设置为5 m,Monte Carlo法获取99%置信区间。采用Programita 2014、Excel 2013、SigmaPlot 12.5进行数据分析及绘图。
2 研究结果
2.1 生境梯度下红松种群数量特征
从图1可以看出,四类生境中,红松种群结构差异较大。在由坡底到坡顶的生境梯度上红松种群密度分别为340、412、160、308 株/hm2。不同林层中红松种群密度亦有较大差异,生境Ⅰ与生境Ⅱ中,红松更新分别达到了226 株/hm2与208 株/hm2,生境Ⅲ与生境Ⅳ中红松更新密度仅为20 株/hm2与60 株/hm2。生境Ⅰ、Ⅱ、Ⅲ、Ⅳ演替层中红松密度分别为56、118、18、72 株/hm2。而在生境梯度上红松原始林亚林层与主林层中红松种群密度有增大趋势,各生境亚林层与主林层红松种群密度分别为8、18、18、52 株/hm2及50、68、104、124 株/hm2。总体而言,在由坡底到坡顶的生境梯度上,红松种群结构有从倒J型向J型转换的趋势。
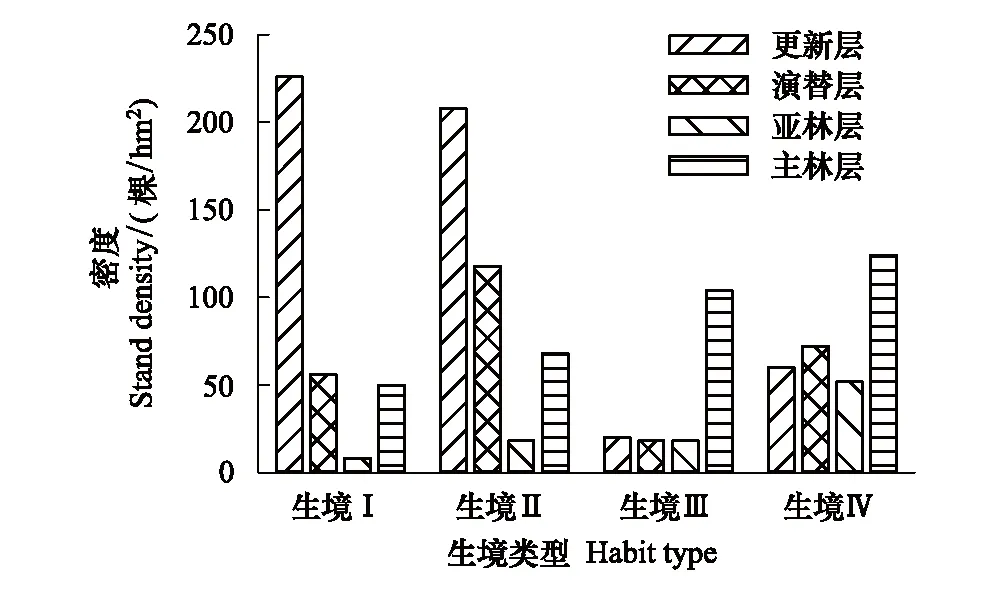
图1 不同生境中原始红松林红松种群结构
2.2 四类生境中各林层红松种群重复采样点格局
将隶属于不同生境的红松林样地作为重复采样,对不同林层的红松立木进行空间点格局分析,并根据零模型与包络线的关系简要换算,用于比较不同生境间红松种群空间分布规律的差异(其中2号样地亚林层中红松不足两株,生境Ⅰ亚林层规律为1号样地亚林层规律)。结果如图2所示:不同生境原始红松林中红松种群在由更新层向主林层生长的过程中,种群空间格局均由聚集分布向随机分布转变;各生境原始红松林主林层与亚林层中红松种群基本均服从随机分布,四类生境原始红松林内红松种群空间分布规律的差异主要体现在更新层及演替层的较小研究尺度上。各生境中,更新层内红松种群聚集强度及聚集规模排序为生境Ⅰ>生境Ⅱ>生境Ⅳ>生境Ⅲ。生境Ⅰ与生境Ⅱ演替层中红松种群在较小研究尺度上体现为聚集分布,而生境Ⅲ与生境Ⅳ中红松种群则表现为空间随机。总体而言,处于相对湿润生境中的原始红松林更新层与演替层中红松种群聚集规模与聚集强度大于处于相对较干旱生境中的原始红松林。
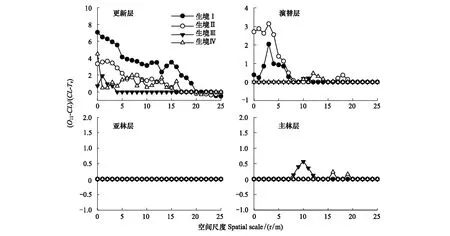
图2 不同生境中原始红松林中各林层中红松种群分布规律比较
2.3 不同生境梯度下红松种群种内联结差异
将分布于不同生境中的原始红松林样地分别作为重复采样,合并分析生境梯度影响下红松种群种内联结关系。基于所得结果中零模型与包络线间的关系,比较不同生境中原始红松林的红松种内联结关系。结果如图3所示:各生境中,亚林层中红松种群在全部研究尺度上均未与主林层表现出空间关联性;更新层、演替层与主林层的红松种群间均表现出一定的空间关联性,相对于更新层与主林层,演替层与主林层中红松种群空间关联性更强。生境Ⅰ原始红松林中更新层中红松种群在1—2 m研究尺度下与主林层表现出微弱的空间负关联;生境Ⅱ更新层与主林层中红松种群在1—5、7、9—12 m的尺度上均表现出弱空间负关联;生境Ⅲ更新层中红松种群与主林层在全部研究尺度上均未表现出空间相关性;生境Ⅳ中,红松林更新层与主林层在7—11 m尺度上表现出空间微弱正关联,其余尺度未表现出空间相关性。生境Ⅰ及生境Ⅱ中原始红松林演替层与主林层在1—5 m研究尺度上表现出空间负关联,而生境Ⅲ与生境Ⅳ中原始红松林则具有相反规律,在较小研究尺度上与主林层表现为空间正关联。
3 讨论
3.1 生境对不同林层红松种群空间分布格局的影响
研究显示,各生境中原始红松林更新层与演替层中红松种群表现出一定的空间聚集性,其聚集强度随着研究尺度增大逐渐减弱,种群空间分布格局逐渐由聚集分布像随机分布转变;亚林层与主林层中红松种群的分布格局趋近于随机分布,规律与前人相同[23-25]。各生境原始红松林演替层中红松种群聚集规模与聚集强度相较于更新层中红松种群均有所下降,体现出红松更新在生长过程中的环境过滤与种内、种间竞争。
各生境原始红松林更新层中红松种群聚集强度与规模有随着坡位增高而减弱的趋势,更新层中红松种群的聚集强度与林下更新数量相关,即更新数量越多,聚集强度越大。各生境更新层中,0—1 m尺度上红松种群聚集强度为生境Ⅰ>生境Ⅳ>生境Ⅱ>生境Ⅲ,而根据调查数据(未发表数据),在0—1 m尺度上更新层中红松种群聚集强度较高的生境Ⅰ与生境Ⅳ中,红松苗高均较低。生境Ⅱ演替层中红松种群聚集强度较大,其原因是该生境调查样地演替层中红松立木多且树高普遍较低(1.5—5 m),这也可侧印以上论点。
相对于生境Ⅲ,生境Ⅳ中林下植被稀疏。根据徐化成的划分,生境Ⅳ中分布的红松林型为典型的陡坡细叶苔草蒙古栎红松林[21],样地内坡度较大处灌木层仅有少量卫矛(Euonymusverrucosus)分布,为红松更新空出了生态位。这或也是相对生境Ⅲ,生境中Ⅳ具有更多红松更新,进而具有更高聚集规模与强度的原因。
总体而言,四类生境中,红松种群从更新层向主林层的发育过程即是一种种群空间格局由聚集分布向随机分布转变的过程;相较于生境Ⅲ与生境Ⅳ中红松种群,处于相对湿润的生境Ⅰ与生境Ⅱ中更新层与演替层中红松种群在较小研究尺度上具有更大的聚集强度与聚集规模,这应与林下幼树、幼苗数量有关;而相对于生境Ⅲ,生境Ⅳ更新层与演替层中聚集规模与聚集强度均更大,这是由于在生境Ⅲ中林下植被发育较为强烈,侵占了红松更新的最适生态位,而生境Ⅳ中林下植被不发达,为红松更新预留了生态位。
3.2 生境梯度下红松种群种内联结差异性
在自坡底到坡顶的生境梯度上,不同的原始红松林型中,亚林层与主林层中的红松种群间空间关系均表现为空间独立、演替层—主林层、更新层—主林层中的红松种群则表现出一定的空间关联性。演替层与主林层间的空间关联强度较之更新层更强,分析其原因应与红松幼苗生长过程中受到的生境过滤作用有关,这使红松幼苗从更新层生长到演替层的过程中逐渐体现出了物种适宜的生态位。
在自坡底到坡顶的生境梯度上,演替层与主林层的空间关联性有从空间负关联向空间正关联转变的趋势,造成这一现象的可能原因有,生境梯度上红松在主林层中的株数占比不断提高,因此红松幼树有更大机会分布在主林层中红松立木周围。这一现象的产生也与不同生境中灌木、草本层的空间分布有关,如生境Ⅲ中林窗中狗枣猕猴桃(Actinidiakolomikta)发育旺盛[13],阔叶树林冠下生境中则常分布有较为茂密的毛榛(Corylusmandshurica)灌丛,这些林下植被的分布,侵占了红松更新及幼树的生态位,使红松幼树、幼苗分布在林下植被较为稀疏的红松林冠下。不同生境中,林下优势灌、草有所不同,在坡位较低、坡度较小的林型内,东北珍珠梅(Sorbariasorbifolia)、东北山梅花(Philadelphusschrenkii)优势度较大,而在坡位较高的生境中,毛榛、刺五加(Acanthopanaxsenticosus)等优势度较大,而林下植被变化均会影响幼树、幼苗的空间分布规律。此外,坡位较高的阔叶红松林中光资源可获取性较强,更新层中红松种群间生长到演替层的过程中受到光资源的制约较小,红松树冠下光环境可以满足幼苗、幼树生长的需求;而在坡底的生境中,红松幼树、幼苗更倾向分布于阔叶树树冠下[34],这与不同林冠下红松幼树、幼苗的保存率有关,中国科学院林业土壤所得材料表面,红松林冠下15年以下的更新不少于阔叶树林冠下,而15年以上的幼苗则以阔叶树下的为高。同时,由于光资源的制约,坡位较低生境中红松树冠下的幼苗生长更慢[35],因此下坡位生境中演替层与主林层中的红松种群在较小研究尺度上表现出空间负关联。植物的建植与生长受到生境过滤的影响,而生境过滤又涵盖环境过滤与生物互做两层意义,前者指非生物环境对植物的筛选作用,后者指植物间的正、负关联[36-38],在探讨生境梯度上植物空间分布规律时,二者都应加以考虑。
4 结论
(1)小兴安岭不同生境的原始红松林中,主林层与亚林层中的红松种群均表现为随机分布;而在自坡底到坡顶的生境梯度上,更新层与演替层中红松种群的聚集规模与聚集强度均有减弱趋势,其原因与在自坡底到坡顶的生境梯度上,红松幼树、幼苗减少有关。
(2)小兴安岭不同类型原始红松林内,亚林层与主林层中的红松种群均表现为空间独立;而在较小研究尺度上,幼苗、幼树与主林层中红松种群间的空间关系则随着坡位上升、坡度增大由空间负关联向空间正关联转变,这是生物与非生物因子共同作用的结果。相较于更新层,演替层与主林层中红松种群的空间正、负关联的程度均更强,这是生境过滤的结果。
猜你喜欢
格言·校园版(2022年17期)2022-07-06 22:01:48
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
落叶果树(2021年6期)2021-02-12 01:29:10
中国林副特产(2020年5期)2020-10-09 08:24:14
防护林科技(2017年7期)2017-09-01 01:08:43
现代营销(创富信息版)(2016年6期)2016-11-21 06:00:03
体育科技(2016年2期)2016-02-28 17:06:07
现代营销(创富信息版)(2016年12期)2016-02-08 02:32:08
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
满族文学(2014年5期)2014-09-22 23:50:36
