
母婴分离应激对幼年大鼠脑衰反应调节蛋白2及微管动态性影响☆
2020-09-15 08:37:44魏艳艳陈景旭吴作天肖玲何静张楠王高华
中国神经精神疾病杂志 2020年7期
魏艳艳 陈景旭 吴作天 肖玲 何静 张楠 王高华
生命早期是神经系统发育的关键时期[1]。早年应激事件可对个体产生持久影响,与抑郁症等疾病的发生密切相关。不同程度早年应激对个体具有不同影响[2],其产生的作用类似于U型曲线,适度温和的应激会使动物成年后表现优于未应激或严重应激大鼠,而其具体发生机制目前并不清楚[3]。脑衰反应调节蛋白2(collapsin response mediator protein 2,CRMP2)在神经系统中高度表达,通过与细胞骨架蛋白相互作用调节骨架动态性[4]。而其活性及功能受翻译后修饰方式影响,磷酸化是其主要修饰形式,磷酸化修饰可导致其与微管蛋白解聚,降低微管动态性,导致神经可塑性受损[5]。而目前关于母婴分离(maternal separation,MS)的研究多集中在应激对成年大鼠神经生物学影响,对幼年大鼠神经可塑性影响的研究较少。本研究旨在研究不同程度母婴分离应激对幼年大鼠海马CRMP2、P-CRMP2及微管动态性的影响,探讨早年应激对幼年大鼠神经生物学影响。
1 对象与方法
1.1 研究对象 10只孕期15~16 d的SPF级SD孕鼠(华中科技大学实验动物中心购入),在武汉大学人民医院动物实验中心SPF环境单笼饲养,安置在12 h:12 h光暗周期的环境中,自由获取食物和水,并保持恒定的温度 (22±1)℃和湿度50%±10%。尽量减少环境刺激,每天定期观察孕鼠生产情况。孕鼠生产的36只新生雄性幼鼠纳入研究。
1.2 研究方法
1.2.1 动物分组及模型建立 将36只新生雄性幼鼠随机分为3组,包括母婴分离360 min组(MS360)、母婴分离 15 min 组(MS15)以及对照组(NC),每组12只。其中MS360组和MS15组从出生后第4~10天进行连续1周给予母婴分离应激,从早上9点开始将幼鼠从母鼠笼中转移至另一个新笼中单独饲养,每日分离360 min或15 min。在分离结束后,幼崽送回母鼠笼中与母鼠团聚。NC组不进行母婴分离。对3组幼崽进行相同次数的抚触,以确保检测到的差异都只是由于分离时间的不同而产生。
1.2.2 标本采集 应激结束后处死幼鼠,迅速在冰面上剥离海马组织,置于-80℃保存待用。
1.2.3 检测海马CRMP2、tubulin、actin的mRNA表达 使用q-RT-PCR方法。每组取6只幼鼠海马组织,用Trizol提取RNA,采用RevertAid First strand cDNA systhesis Kit#1622试剂盒进行逆转录合成 cDNAs。采用 SYBR®Premix Ex TaqTM RR820A试剂盒在CFX96型qPCR仪上进行荧光定量PCR反应,引物由谷歌生物公司合成,引物序列见表1。以 GAPDH的 Ct值为参照,采用2-ΔΔCt反映mRNA相对表达水平。
1.2.4 检测海马CRMP2、P-CRMP2及微管动态性相关蛋白表达 使用Western-Blot方法。每组剩余6只幼鼠海马组织,采用RIPA裂解液进行裂解匀浆,离心后收集蛋白上清液,采用BCA蛋白定量试剂盒进行蛋白浓度测定。将蛋白样品煮沸变性,取适量样品(20 μg)进行 10% SDS-PAGE 电泳,并转移至PVDF膜上,5%脱脂奶粉室温封闭1 h,洗膜后分别加入抗CRMP2抗体(ab129082,1:200000)、抗 P-CRMP2 抗体(ab85934,Thr514 位点,7 μL:5 mL)、抗 tubulin 抗体(ab7291,1:5000)、抗 Tyr-tubulin抗体 (T9028,1:2500)、抗 Acettubuin 抗体(sc-23950,1:200)、抗 actin 抗体(sc-8432,1:250)以及抗 GAPDH抗体 (#2118,1:1000),4℃摇床孵育过夜。次日加入对应二抗室温孵育1 h,在PVDF膜上滴加ECL化学发光试剂,置于BIO-RAD ChemiDoc Touch Image System仪器上进行曝光显影。应用Image-Lab软件对条带进行灰度值分析,将GAPDH的灰度值作为参照标准。

表1 q-RT-PCR引物序列
1.3 统计学方法 采用SPSS 21.0进行统计分析。采用单因素方差分析进行组间比较,进一步两两比较使用LSD法。检验水准α=0.05,双侧检验。
2 结果
2.1 海马 CRMP2、tubulin、actin的 mRNA 表达水平 MS360组、MS15组以及NC组海马CRMP2、tubulin、actin的mRNA表达水平差异无统计学意义(P>0.05)。见表 2。
2.2 海马CRMP2、P-CRMP2及微管动态性相关蛋白表达水平 3组的P-CRMP2蛋白表达水平差异具有统计学意义 (F=11.172,P=0.001),MS360组高于 MS15组及 NC 组(P<0.01),MS15组与 NC 组差异无统计学意义(P>0.05)。3组Tyr-tubulin水平差异有统计学意义 (F=4.311,P=0.049),MS360组较 MS15组及 NC组下降 (P<0.05),MS15组与NC组无统计学差异 (P>0.05)。3组Acet-tubulin水平差异具有统计学意义 (F=8.939,P=0.003),MS360组高于 MS15 组及 NC 组 (P<0.01),MS15组与 NC组无统计学差异 (P>0.05)。CRMP2、tubulin、actin蛋白表达水平3组间无统计学差异(P>0.05)。见图 1 及表 3。
3 讨论
啮齿类动物母婴分离模型是目前公认的早年应激模型。动物研究发现不同程度母婴分离应激对成年后行为具有不同影响:短期母婴分离应激可促进个体出现应激抵抗,避免抑郁样行为的产生;而长期母婴分离则会导致个体出现应激敏感,出现明显的抑郁样行为[2,6]。其发生机制目前并未阐明,且目前研究多集中在母婴分离应激模型成年后神经生物学指标改变,对幼年时期神经可塑性影响的研究较少,故本研究旨在探索不同程度母婴分离应激对幼年大鼠神经可塑性机制的影响。
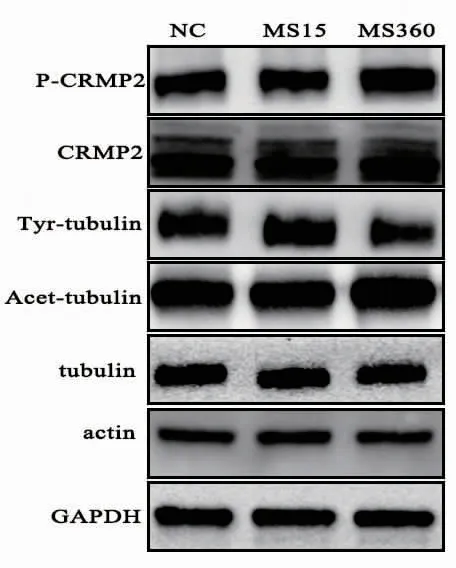
图1 母婴分离应激后3组幼鼠海马各蛋白表达灰度图 NC为对照组;MS15为母婴分离15 min组;MS360为母婴分离360 min组。

表 2 幼鼠海马 CRMP2、tubulin、actin 的 mRNA 表达水平(2-ΔΔCt)

表3 幼鼠海马各蛋白表达水平
本研究结果显示长期母婴分离导致幼鼠海马P-CRMP2表达水平升高,而短期母婴分离并未引起P-CRMP2水平改变,并且两者均未影响海马CRMP2水平。提示长期母婴分离应激主要对幼年大鼠海马CRMP2的磷酸化水平产生影响,而对CRMP2的总体蛋白及mRNA水平无明显影响,而短期母婴分离应激对幼鼠海马CRMP2及PCRMP2均未产生影响。CRMP2与神经可塑性调节密切相关,其在神经系统广泛表达,并在微管动态性调节、轴突生长、神经元极化及迁移等调节中发挥重要作用[7]。磷酸化是CRMP2主要修饰形式,CRMP2磷酸化可导致其与tubulin等下游分子的结合能力受损,进而影响微管聚合及轴突生长等功能[8]。既往研究发现CRMP2与应激、精神疾病有密切关联[9]。尸脑蛋白质组学研究发现抑郁症患者脑内CRMP2的表达水平较对照组下降[10],慢性不可预计性应激小鼠模型脑内CRMP2表达水平亦下降[11]。抗抑郁药物可影响脑内CRMP2的水平,情绪稳定剂锂盐可降低脑内CRMP2的磷酸化水平[12]。这些研究均提示CRMP2以及P-CRMP2在应激、抑郁症的发病机制以及抗抑郁药物作用机制中起着重要作用。动物研究发现早年长期母婴分离应激与抑郁症密切相关,会导致个体成年后出现抑郁症等精神疾病[13]。本研究结果发现早年长期母婴分离应激会导致幼鼠海马P-CRMP2水平升高,结合以上文献结果,提示大鼠成年后出现抑郁症等精神疾病可能与个体幼年时期海马CRMP2的磷酸化水平改变有关。但本研究并未发现早年母婴分离应激对CRMP2总体水体有影响,却发现长期母婴分离应激会导致幼年大鼠CRMP2的磷酸化水平升高,提示不同时间及不同程度应激可能对大脑产生不同影响[14]。
细胞支架在神经可塑性调节中发挥重要作用,微管动态性变化是神经元形态、功能以及生长发育的基础[15]。tubulin为微管蛋白,actin为微丝蛋白,均为细胞支架的构成蛋白。乙酰化微管蛋白(Acet-tubulin)及酪氨酸化微管蛋白(Tyr-tubulin)为tubulin的修饰形式,Acet-tubulin为稳定微管标志物,Tyr-tubulin为动态微管标志物[16-17]。本研究发现长期母婴分离应激导致Tyr-tubulin表达下降,Acet-tubulin表达升高,而短期母婴分离应激并未影响其表达水平,提示长期母婴分离应激导致微管动态性下降、神经可塑性受损,而短期母婴分离未影响微管动态性。并且本研究发现早年应激并未影响 tubulin、actin蛋白及 mRNA表达水平。既往关于抑郁动物模型的研究发现其海马Acet-tubulin升高,Tyr-tubulin下降,存在微管动态性受损[17],提示应激导致抑郁样行为的发生机制可能与微管动态性受损有关。本研究与其研究结果相似,且发现长期母婴分离应激导致CRMP2磷酸化水平显著升高。既往研究发现磷酸化修饰可降低其与微管蛋白结合,影响微管的聚合与解聚动态调节[5]。因此早年长期母婴分离应激导致微管动态性下降的机制可能与CRMP2磷酸水平升高相关。
本研究还存在一定的局限性:研究材料均为雄性幼鼠,未进行雌性幼鼠的研究;此外,只检测了幼鼠神经生物学指标,未进行成年后大鼠行为学及神经生物学指标的研究。下一步研究需扩大范围,将雌鼠及成年鼠纳入研究,从而得到更有说服力的研究成果,以期为探明早年应激的神经病理学机制提供依据。
综上所述,本研究结果提示,早年长期母婴分离应激导致幼年大鼠海马微管动态性下降,神经可塑性受损,这一过程的可能机制源于CRMP2的磷酸化水平升高;而短期母婴分离并未影响微管动态性及CRMP2的磷酸化水平。不同程度母婴分离应激在大鼠幼年时期已经引起海马发生不同的生化改变,这可能是早年应激导致成年后行为发生改变的基础,本研究对于进一步阐明应激影响的神经机制研究具有一定启发意义。
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:40
——水芹主要害虫识别与为害症状
长江蔬菜(2022年13期)2022-07-29 01:21:32
山东警察学院学报(2021年2期)2021-08-24 07:08:30
郑州大学学报(医学版)(2021年1期)2021-03-03 01:06:30
池州学院学报(2017年5期)2018-01-23 02:54:27
自然资源情报(2017年11期)2017-11-26 08:02:26
中外医疗(2016年15期)2016-12-01 04:25:49
饲料博览(2016年6期)2016-04-05 14:11:14
首都医科大学学报(2015年4期)2015-12-16 13:00:08
武汉理工大学学报(交通科学与工程版)(2015年5期)2015-12-05 02:20:00
