
中国湖泊古生态研究进展
2020-09-09 09:04:54羊向东董旭辉
湖泊科学 2020年5期
羊向东,王 荣,董旭辉,王 倩,陈 旭,徐 敏,张 科
(1:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京 210008)(2:广州大学地理科学与遥感学院,广州510006)(3:中国地质大学(武汉)地理与信息工程学院,武汉430074)
湖泊古生态学(Lake Palaeoecology)是利用湖泊沉积记录中保存的各种生物遗存,研究远近地质时期生物群落演替的过程,进而重构过去食物网各营养级生物组合以及生物与它们所处环境之间的关系. 湖泊古生态研究兼顾地质学和生态学属性,具有多学科交叉和综合的特征. 作为地球表层物质和能量交换最为频繁的地理单元,湖泊生态环境的变化受到流域过程、气候变化、人类活动以及它们之间相互作用的影响[1]. 因此,湖泊古生态研究的一个主要目标就是获得湖泊生态演化的宏观规律性认识,厘清不同时空尺度气候变化和人类活动的影响机制[2-3]. 湖泊古生态研究也是生态学向过去的延伸,可以从时间上弥补生态试验和生态系统监测数据过短的缺陷,提供特定研究时段的物种多样性、群落结构更替和系统弹性变化的信息,揭示食物网中生物相互作用(如竞争、捕食等)对生态系统的影响,检验生态学理论,如临界转换等. 近年来,湖泊古生态研究在帮助界定、描述和指导生态系统管理方面也得到广泛应用[4].
湖泊古生态学是古湖沼学的一个主要分支学科,其研究始于20世纪初期,当时仅限于沉积硅藻的分类和生态信息的定性描述及其对环境的示踪研究. 直至20世纪中期以来,硅藻指标被陆续运用到古气候重建、湖泊富营养化和酸化等环境问题研究中. 最近40 a来,随着科学技术的不断创新和分类学的发展,以及湖沼学与古湖沼学的不断融合,各种生物遗存的指标被挖掘,标准分析方法也纷纷建立,数据统计分析手段得到广泛应用,湖泊古生态研究学科体系逐步得到完善,研究主题涉及到第四纪地质学、湖泊科学、生态学、环境科学等诸多领域[5-8]. 围绕过去全球变化、湖泊-流域相互作用、人与自然对湖泊生态系统影响等科学问题,开展了从基础到应用基础的系统性研究,并实现了从定性描述到定量表达的跨越发展.
我国湖泊古生态研究开始于1990s. 尽管起步较晚,但研究者们围绕过去全球变化和我国区域湖泊生态环境问题开展了大量的工作,并取得系列重要进展. 本文在简要介绍湖泊古生态研究基本原理的基础上,重点综述近20 a来在环境定量化、不同时间尺度区域湖泊古生态重建、生态系统演变与突变等方面的主要研究成果,以及这些成果在指导湖泊管理和治理中发挥的作用. 最后,对湖泊古生态研究的前景进行了展望.
1 湖泊古生态指标与方法学基础
湖泊中的各种生物有机体死亡后,经沉降和搬运,最终以其难以降解的几丁质、硅质、文石、方解石等壳体和骨骼或者分子构架成分,保存在沉积物中. 通常称为化石生物指标或古生态指标,是湖泊古生态研究的主要对象. 这些指标大体可分为外源和内源两大类. 外源性指标主要来自流域植被,常见指标包括陆生植物的残体、孢粉、炭屑、植硅石等;内源性指标来自大型水生植被及其花粉、细菌、各种浮游动植物和底栖动物等. 古生态研究就是通过不同来源生物指标的群落组成、丰度变化等信息,进行过去生态环境演化的重建. 本文重点针对内源性指标进行国内湖泊古生态研究进展介绍.
表1中列举出了常见的湖泊化石生态指标及我国学者的主要研究文献. 这些指标中,硅藻、摇蚊、枝角类是最广泛使用的古生态指标,在我国许多湖泊沉积物中均有较好的保存,这些指标已经分别用于对过去湖水营养、盐度、水深和温度等生境条件的推导. 介形类和有壳变形虫化石指标也较常见,但它们分布在特定的湖泊中,如介形类主要集中分布在青藏高原和西北干旱区封闭湖泊,其群落组合常被用于指示盐度和水文的变化,但目前介形类化石群落的研究远不及其壳体地球化学研究的深入. 有壳变形虫属于原生动物,目前的研究都集中在我国三江平原等沼泽湿地,在指示湿地水位和湖泊营养变化方面具有优势. 沉积物中保存了丰富的沉水、漂浮/浮叶、挺水植物残体及果实,尤其适用于浅水湖泊的水生植物群落演替的重建. 将大型水生植物残体与花粉指标结合,在回答草/藻稳态转换的生态学问题、指导水生植被修复实践中有一定优势. 近年来,一些特定生物标记化合物指标正备受关注,以藻类色素、正构烷烃和GDGT(甘油二烷基甘油四醚脂类化合物)指标等为代表. 藻类在光合作用过程中合成多种色素(如:叶绿素、类胡萝卜素和叶黄素等),大部分色素会在水体中经化学氧化、捕食和微生物降解等过程而分解,但一些稳定性好的色素能较完整地保存在沉积物中,而且具有十分明确的指示性意义,如双四氧嘧啶(alloxanthin)和海胆酮(echinenone)能有效指示隐藻和蓝藻的变化[9]. 具有稳定类脂物分子的正构烷烃,依据碳链长短和奇偶优势,结合正构烷烃不同碳链组合与比值,可以区分不同生物来源,指示湖泊各种藻类、水生植物类型和流域植被的发育状况. GDGTs主要由两条烷基链与甘油分子以醚键形式合成,研究认为其在指示古温度和pH值重建中具有潜力.
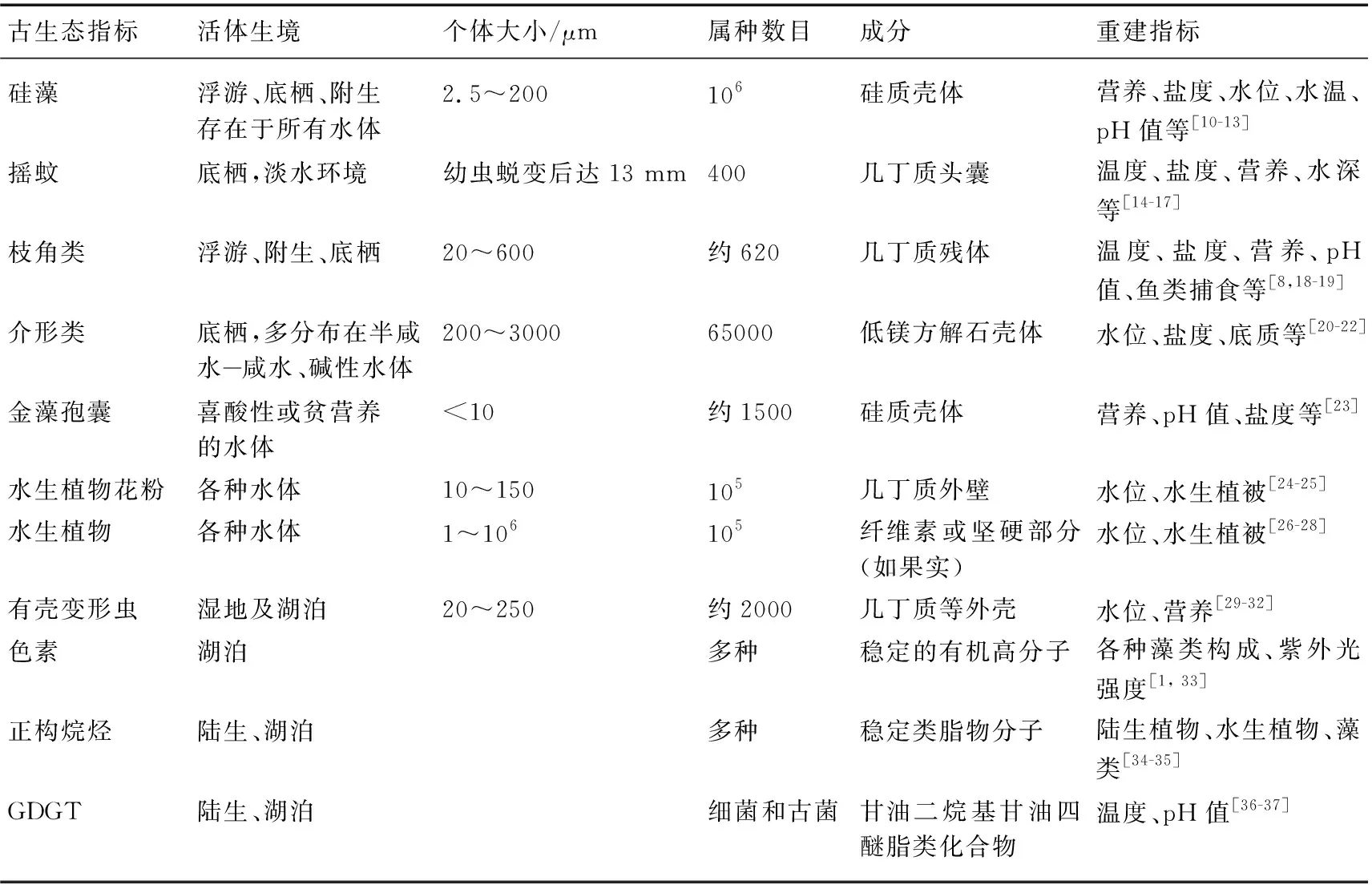
表1 常用的古生态指标特征及其环境指示意义
湖泊古生态重建基于两个重要的假设前提:(1)沉积物中的生物化石组合应同当时水体生物群落主要构成一致. 尽管一些研究认为诸多生物门类沉积过程中存在一定程度的损失,但总体上仍能反映当时的主要物种类群组合[38-39]. Wang等[40-41]通过对泸沽湖不同水层水体和沉积物捕获器的硅藻与枝角类多年监测数据的对比分析表明,沉积物中主要的硅藻和枝角类属种与水体优势属种和组合基本对应. (2)均变假说,即各个生物物种在过去和现今的生理特征、生态习性不变,对外界环境变化响应的物理机理一致[42]. 换言之,影响现代生物丰度和分布的环境因子可以作为推断控制古生物丰度以及分布变化的环境因子.
基于上述的假设,“将今论古”的研究思路被古生态工作者广泛接受. 借助于湖泊现代过程监测和空间调查,更多的现代生态学信息被用于古生态数据的解释和环境推导中. 各种数值分析技术的应用,为分析物种和群落分布特征、揭示生物对环境的响应模式、定量重建环境变化提供了重要途径. 数值分析包括各种统计和回归分析方法,目前古生态研究者普遍使用的CANOCO和R软件可以满足多种分析用途[43-45],如利用主成分分析(PCA)和降维对应分析(DCA)等,可以判识生物分布的主要特征及其对环境梯度响应的线性或非线性响应模式;利用冗余分析(RDA)和典型对应分析(CCA),可以揭示时间和空间尺度上环境对生物群落分布影响的重要性和显著性,同时可以对不同环境要素的影响程度进行量化. 环境定量重建方法包括加权平均(WA)、回归树(Regression Tree)、贝叶斯函数(Bayesian)、神经网络(Neural Network)等方法[46-48],如基于回归树的多要素定量重建,其优势在于可以通过限制其他环境梯度,优化目标环境梯度,同时能对环境变量突变的阈值提供信息[49].
2 湖泊古生态定量化研究
环境要素的定量化一直是古湖沼学研究的重点,对认识过去气候和湖泊环境演化规律、模拟和预测未来变化趋势有着重要的意义. 湖泊环境要素的定量化还可实现与现代监测数据的对接,评估人类干扰前的自然本底特征. 由于水生生物对湖泊理化环境变化具有快速、敏感的响应特点,硅藻、摇蚊类等生物以及生物标记化合物指标尤其适合开展环境要素的定量重建研究,其中以生物-环境转换函数研究成果最具代表性. 其基本思路是通过对特定区域内多个湖泊的调查,建立表层沉积生物和水体理化环境数据库;利用多种数理统计方法,分析并提取影响生物分布的显著环境变量,并判识两者之间的线性和非线性关系;然后选择不同的多元回归分析手段,在对环境推导值与观测值间误差、推导能力等比较的基础上,获得合适的转换函数模型;最后依据沉积记录的生物数据,进行古环境要素的定量重建,并对重建的效果进行评估(图1).
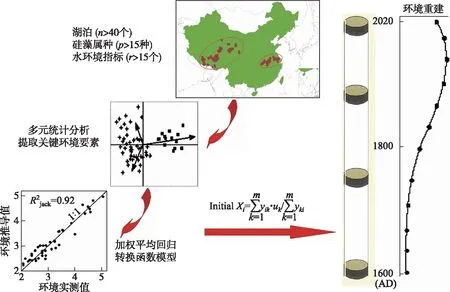
图1 转换函数研究技术思路图Fig.1 Technical diagram of transfer function
在我国,古生态研究者已经建立了包括青藏高原、西北、西南和长江中下游地区湖泊的生物转换函数数据库,湿地有壳变形虫转换函数也在东北地区建立. 这些转换函数被成功应用到不同区域湖泊和湿地水体的盐度、营养浓度、水深、温度等环境要素的定量重建中. 例如,作者基于青藏高原近80个湖泊表层沉积硅藻-环境数据库的分析,通过典型对应分析,揭示出电导率(或盐度)是影响高原湖泊硅藻分布的显著环境变量,利用加权平均-偏最小二乘法(WA-PLS),建立了硅藻-电导率(盐度)转换函数,并对西藏沉错和纳木错过去百年来盐度变化进行了定量恢复[50-51]. 研究结果表明,小冰期结束以来至20世纪末,随着区域增温,湖水盐度逐渐增高,与多年观测的水位持续下降对应. 2000年之前,浮游硅藻丰度增加和盐度的升高,一方面与升温引起蒸发量的增加有关,另一方面与冰融水增多导致流域营养物质的输入增多有关.
人类活动干扰引起的湖泊富营养化是全球性问题,长江中下游浅水湖泊富营养化问题尤为突出. 为揭示富营养化原因与趋势,评估强烈人类活动前的营养本底值,我国学者利用长江中下游近90个湖泊的硅藻、摇蚊、有壳变形虫和水质调查数据,分别构建了它们与湖水总磷浓度的转换函数模型[10, 52-53]. 基于沉积钻孔的生物化石数据,先后对10个以上不同营养类型湖泊进行了过去百年来湖水总磷浓度的定量估算[54]. 基于不同定量模型获得的总磷值,在变化趋势和幅度上有很好的一致性. 研究发现近百年来,这些湖泊富营养化普遍发生在1950s后,至1980s以来一些重要湖泊如太湖和巢湖的富营养化程度进一步加重. 这些研究显示,长江中下游地区湖泊总磷本底值维持在50 μg/L左右,湖泊由草型向藻型转变的阈值在80~110 μg/L之间. 上述研究表明,研究区浅水湖泊普遍具有较高的营养背景值,在富营养化湖泊治理中,对营养消减目标的设定应该从历史演变的过程中去寻找参照系.
西南高山地区,湖泊分布在不同海拔,这也决定了在空间上山地湖泊生物群落组成与气候存在一定的关系. Zhang等采集了近100个高山湖泊表层沉积物硅藻和摇蚊数据,并利用GCM输出界面和地形参数方法,建立了研究区湖泊生物和气候环境数据集[55]. 统计学分析揭示出夏季温度(或水温)是显著影响因子,由此分别基于加权平均回归和增强回归树方法建立了摇蚊-夏季温度、硅藻-夏季温度的转换函数模型. 目前这些转换函数模型正用于西南地区多个湖泊末次盛冰期以来的古气候定量重建[56-57]. 上述研究不仅可为定量重建结果的相互验证和区域比较提供独立依据,也为深入认识西南季风气候演化对全球变化的响应及机制提供了关键证据.
3 自然和人类驱动的湖泊生态环境演变特征与机理
湖泊生态系统演变经历了从自然驱动到人与自然共同驱动的历程. 围绕全球变化和生态响应主题,我国湖泊古生态研究目前主要关注末次盛冰期以来、全新世以及过去200 a来的人类世3个时段,开展了不同时间尺度的湖泊关键生态环境过程重建研究,并以“气候-人类活动-流域-湖泊环境”关系链分析为主线,探讨了湖泊生物群落结构演替和环境变化及其与流域过程、气候、构造等自然因素和人类活动的关系.
3.1 末次盛冰期以来的湖泊生态环境演变
地质历史时期我国湖泊自然生态环境的变化主要受季风气候的影响. 已有的湖泊古生态研究主要来自我国东北和南部地区的玛珥湖以及西南地区的断陷湖泊. Wang等[13,58]通过云南泸沽湖沉积钻孔的硅藻等记录,重建了过去3万年来湖泊生物群落结构演替的历史,研究得出硅藻群落的演替与水体分层/混合、营养等关键环境过程有关. 并在此基础上,结合孢粉和环境替代指标等多指标综合分析,较深入地探讨了湖泊-流域相互作用对生物群落演替的影响机理,揭示出在太阳辐射的驱动下,温度通过直接影响湖泊水体分层/混合持时,季风降水变化通过影响流域过程(植被发育、土壤侵蚀、有机物和营养输出),直接或间接地影响了湖泊热量平衡和营养循环过程,最终导致湖泊生物群落结构的演替(图2). 来自中国东北一玛珥湖(小龙湾)和云南一高山湖泊(天才湖)过去1万多年来的硅藻古生态研究发现,硅藻群落演替敏感响应于晚冰期结束至全新世开始前一次气候突然降温事件(新仙女木事件). 研究认为气候变冷通过延长湖泊水体冰封时间、缩短水体混合和热量分层时间、降低流域溶解有机质等营养输入,进而影响湖泊营养、pH值等与生物间相互作用,最终导致藻类群落结构的变化[59-60]. Liu等[61]利用黄土高原地区一山地湖泊(公海)沉积钻孔过去2000 a来的硅藻记录,发现了隋唐暖期湖泊自然富营养化过程. 研究进一步对比了现代暖期下的湖泊生态环境特征,认为在历史时期季风降水通过影响流域侵蚀,导致湖泊富营养化及硅藻生态响应,而在现代气候变暖背景下,快速工业化产生的气溶胶造成亚洲夏季风减弱,导致其对山地湖泊生态环境的差异响应特点.
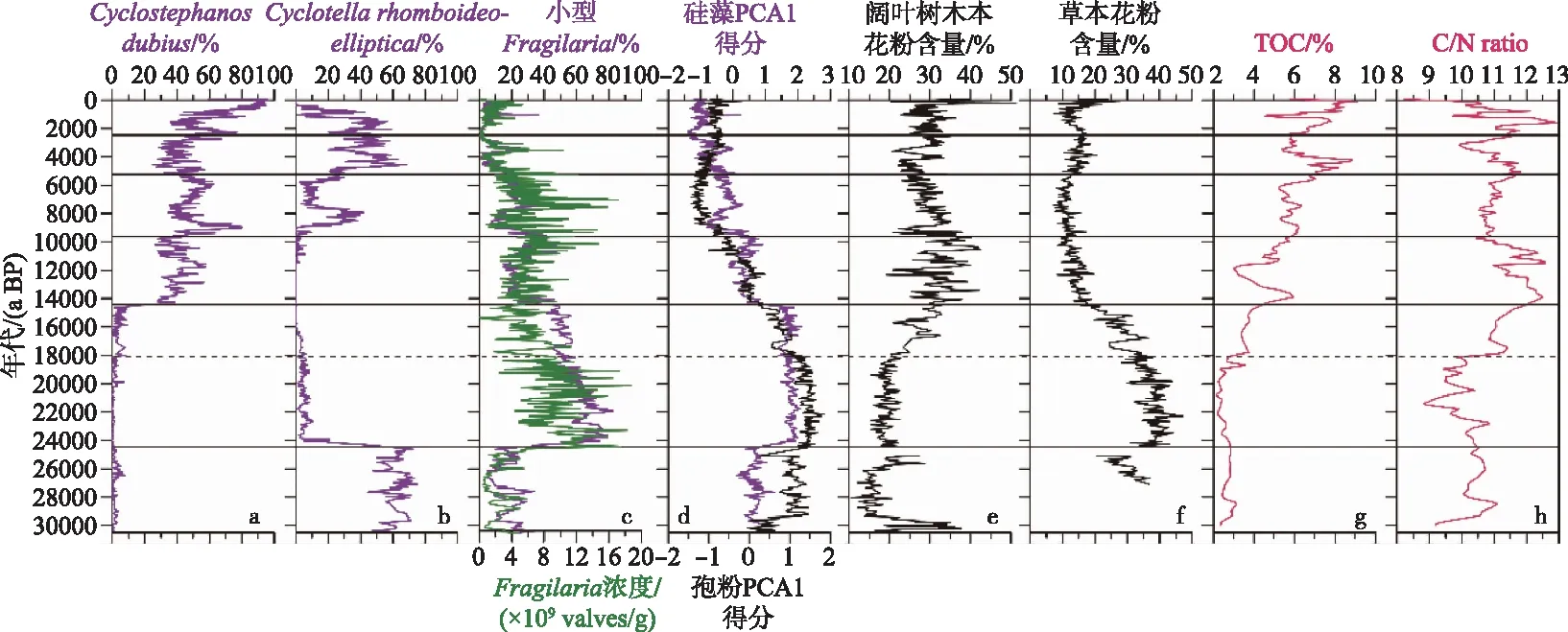
图2 末次盛冰期以来泸沽湖硅藻群落演替及其与流域植被和环境的关系Fig.2 Diatom community succession in Lake Lugu and its relationship with catchment vegetation and environment since the Last Glacial Maximum
基于生物对气候响应关系,一些学者尝试将生物指标直接用于古季风的重建中[62-63]. 例如,Wang 等[62]通过广东湛江湖光岩玛珥湖硅藻监测数据,分析了硅藻季节性分布的规律及其与环境指标的关系,发现颗粒直链藻(Aulacoseiragranulata)的发育与冬季强风速有关,星状小环藻(Cyclotellastelligera)的发育与水体热分层和营养有关. 随后,利用时间序列上这两个主要指示种的丰度和比值(AG/CS比值)与季风指数变化进行了比较,进一步证实了其对季风气候的指示意义. 由此利用湖光岩玛珥湖沉积硅藻记录重建了1.4万年来东亚冬季风演化的历史.
利用外源性孢粉、植硅体和炭屑等指标,全新世早期人类活动的足迹已经逐渐被揭示[64-65]. 这些研究探索了全新世中期以来人类活动对流域植被生态景观的影响,以及人类对农作物驯化和传播的历史. 但早期人类活动在多大程度上影响到湖泊生态系统很少被关注. 来自洱海全新世钻孔的多指标研究揭示出洱海流域早期人类活动开始于中全新世(6000 a BP前后),森林砍伐引起流域土壤侵蚀作用增强,硅藻中耐营养种Cyclostephanosdubius的丰度明显增加,反映流域早期人类活动已开始对洱海硅藻群落产生一定影响,但钻孔硅藻数据的DCA分析显示湖泊硅藻生态变化并没有明显偏离自然演化的轨迹. 泸沽湖晚全新世以来沉积钻孔多生物指标序列(硅藻、藻类色素、枝角类)的高分辨率记录,同样揭示了以自然气候变化主导的湖泊生态系统响应的特点[66].
古生态记录也很好地揭示了构造-气候-流域相互作用对湖泊形成发育和藻类生态的影响. 位于我国西南高山地区的木格措是一个典型的由地震滑坡堵塞沟谷形成的古堰塞湖. Hu 等[67]通过沉积硅藻分析,结合岩性、年代学、粒度、有机碳、δ13C等证据(图3),发现该湖于9000 a BP前后形成. 在此时间之前该湖为沟谷沼泽湿地,但此后浮游硅藻丰度呈现快速增加趋势,指示了湖泊水位持续上涨,并一直维持到现在. 基于西南地区湖泊硅藻-水深转换函数重建的全新世水深变化趋势,与钻孔硅藻数据PCA第1轴得分呈显著反向关系;而PCA第2轴得分与区域温度和孢粉记录大体一致,说明影响木格措硅藻古生态变化的主导因子是水位,其次是气候. 研究进一步探讨了在构造和气候共同驱动下,藻类群落演替响应水文、水体分层/混合、光透度等环境变化的机理.
3.2 人类干扰下的湖泊生态环境演变
最近一个世纪,全球环境变化已经从全新世以自然因素影响为主发展到以人类活动影响为主的人类世时期[68]. 针对湖泊生态系统退化原因、环境胁迫下生物群落演替与响应等问题,我国学者利用浅钻沉积物硅藻、摇蚊、枝角类、水生植物、水生花粉、正构烷烃等指标,对不同地区近现代湖泊古生态开展了系列重建研究,取得了重要规律性认识. 在人类干扰强烈的长江中下游地区,多个浅水湖泊古生态记录的对比表明,区域湖泊过去200 a来生态演化主要存在3种模式:(1)以巢湖和太白湖等为代表的富营养湖泊,生态环境都经历了两次群落组合演替的过程(分别在1950s-1960s和1980s-1990s)[10,69-70];(2)以龙感湖和梁子湖等为代表的中营养湖泊,只经历了一次生物群落组合演替的过程(1950s-1960s)[26,71-72];(3)不同于上述湖泊,大型通江湖泊鄱阳湖和洞庭湖的生物群落演替则发生于1990s中期,湖泊开始富营养化,2003年后群落结构又一次出现明显变化,耐营养种明显增多[73-74]. 作者归纳总结了区域湖泊近百年来生态环境变化的总体趋势. 1950s前,长江中下游湖泊水体硅藻以颗粒直链藻为优势组合,水生植被以清水型沉水植物类型为主;1950s后,除鄱阳湖和洞庭湖等少数通江湖泊外,沉水植物和浮叶植物发育,附生硅藻种类丰度增加,并出现少量耐营养种类; 1980s后随着富营养化程度的加重,一些湖泊(如太白湖等)耐营养硅藻种类和丰度明显增加,水生植被向浮叶和漂浮植物类型组合转变,而在更高营养环境下,巢湖和西太湖水生植被快速消失(图4).
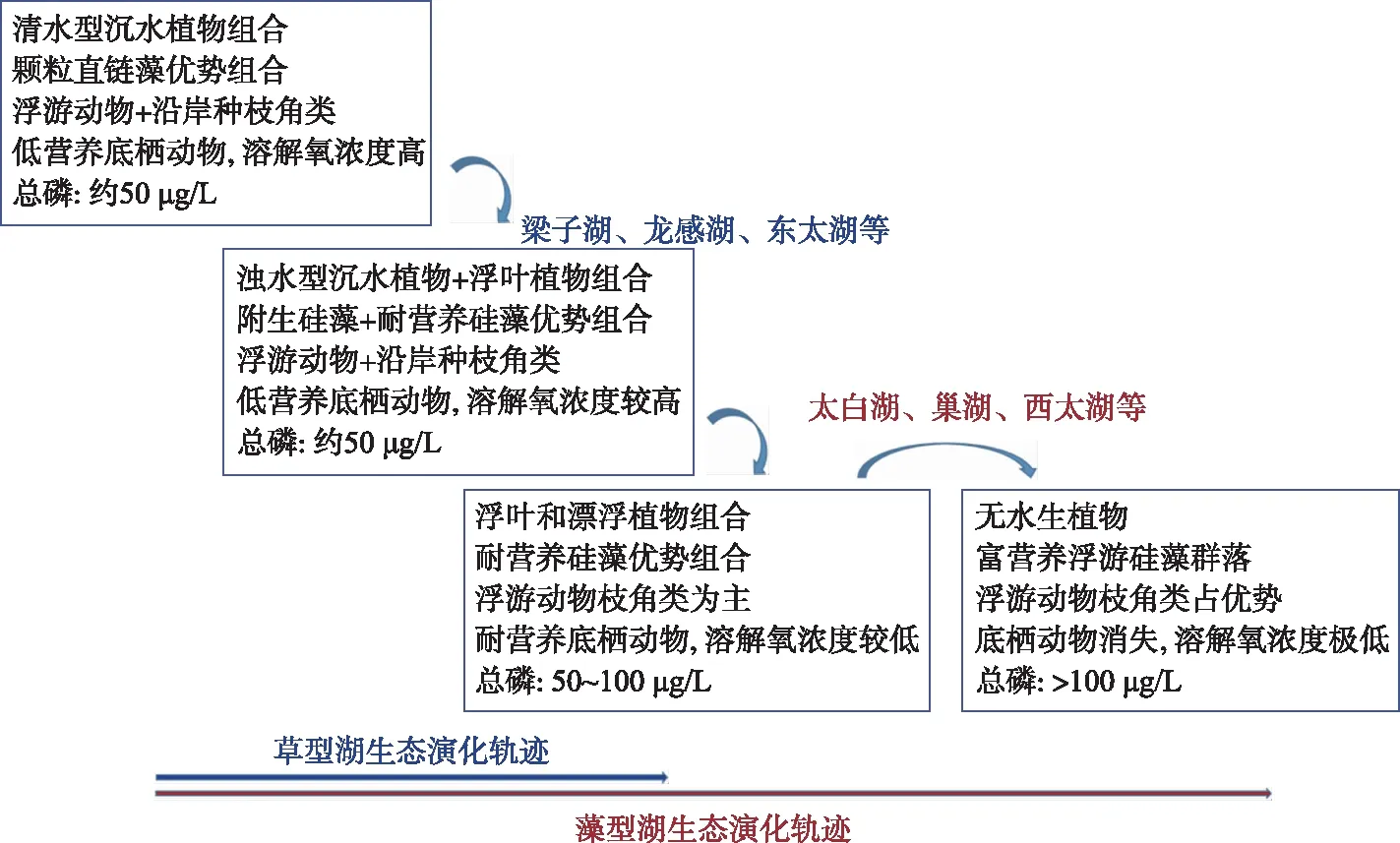
图4 近百年来长江中下游湖泊生态演化阶段示意图Fig.4 A diagram of the ecological evolution stages of lakes in the middle and lower reaches of the Yangtze River in the past century
为揭示生态响应原因,利用典型湖泊多指标综合分析,借助于各种统计分析手段,研究揭示出不通江的湖泊1950s-1960s普遍发生的群落组合演替与江湖阻隔导致的水文变化有关,1980s-1990s的群落演替和显著富营养化是流域营养输入持续增加的结果. 硅藻、摇蚊等生物指标与气候、水位和营养指标的冗余分析(RDA)与方差配置分析表明,近几十年来,增温及其与营养的相互作用对生物群落更替也产生了一定的影响. 鄱阳湖和洞庭湖1990s中期生物群落演替与水位下降有关,导致枯水期一些湖湾换水周期延长,水体营养富集,藻类生产力提高. 而2003年后的富营养化趋势加重,可能是三峡工程运行后长江中下游水位下降,枯水期湖湾进一步与湖泊主体隔离的后果.
上述古生态数据也为研究区湖泊的生态本底评估提供了依据. Dong等[54]利用时空置换的方法,结合现代硅藻数据库和多个湖泊沉积钻孔的硅藻数据,通过生态距离计算(弦距分析),评估了人类干扰前(1850s前)的本底生态特征,即湖泊硅藻群落总体以喜流动水体的颗粒直链藻为主,以附生底栖种类为辅. 本底与表层硅藻的生态距离分析还显示,大多数湖泊目前的生态已经偏离了本底水平. 由于研究区近200 a来显著的硅藻演替发生在1950s,Zhang等[73]提出了可以将该时间之前的生态状况作为湖泊生态修复的人类世本底. 水生植被的历史反演,也可为富营养湖泊的水生植被修复提供参考依据. Zhang等[26]根据水生植被演替的阶段性特征,提出了逐步优化水生植物群落结构的修复对策,并将历史时期清水型沉水植物种类组合作为富营养湖泊修复的最佳参考目标.
位于西南地区林线附近的高山湖泊,其古生态研究揭示出1850年前藻类、枝角类群落组合和生物量的变化敏感响应于冰期结束前后的温度变化,但近百年来藻类生物量的增高则反映了额外的营养供应,提出在无直接人类活动干扰下,大气活性氮沉降的增加和区域气候共同影响了高山湖泊浮游植物生物量的升高[75-76]. 为了区分气候和污染沉降的影响,Hu等[77]通过对多个高山湖泊的沉积记录的硅藻数据集成分析,获得了区域生态变化的趋势(图5). 在此基础上对比了近百年来区域大气氮沉降、粉尘沉降、温度等变化,揭示出大气沉降带来的营养物质输入对高山湖泊生态环境的影响程度已经超过气候变化的影响. 但不同湖泊的响应存在差异,流域植被、土壤稳定性、溶解有机物的输入对湖泊生态系统起着一定的调节作用. 例如,在周围植被覆盖率低的情况下,湖泊的生境更易受到侵蚀和污染的影响,其生态响应比流域植被茂密的高山湖泊更明显.
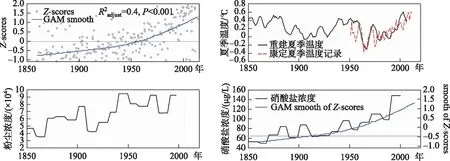
图5 近150 a来西南地区高山湖泊的生态变化趋势与温度、粉尘和氮沉降变化的比较(Z-scores是利用9个高山湖泊沉积钻孔硅藻数据的PCA得分的集成分析结果;区域夏季温度、粉尘和氮沉降数据分别取自文献[78-79];氮沉降数据用喜马拉雅冰芯硝酸盐浓度数据表示)Fig.5 Comparison of the ecological change of alpine lakes in southwest China with the changes of temperature, dust and nitrogen deposition in recent 150 years (Z-scores is an integrated PCA score analysis using diatom data from sediment cores in 9 alpine lakes; regional summer temperature, dust and nitrogen deposition data were obtained from references [78-79]; nitrogen deposition data were shown by nitrate concentration from Himalayan ice core)
不同于西南小型高山湖泊,云南断陷湖泊受人类活动直接影响,近百年来的硅藻、枝角类和摇蚊记录揭示了1980年以来这些断陷湖泊生态普遍开始变化,明显的转变出现在1990s末或20世纪初[66,80-81]. 硅藻群落组合中,一些曾经广泛分布的地方种如椭圆菱形小环藻(Cyclotellarhomboideo-elliptica)等在最近几十年来几乎消失,取而代之的是一些对营养增加敏感响应的先锋种类,与之相应的藻类生物量也明显升高. 泸沽湖底栖摇蚊沉积记录表明,1980年前后流域土壤侵蚀的加强导致摇蚊群落开始变化,底栖动物生物量下降,1995年后进一步下降,与流域大规模的森林砍伐和农业活动导致的水体营养和悬浮颗粒物质明显增加有关,对摇蚊的栖境、下水层溶解氧及食物可得性均产生了较大影响,进而改变了底栖种群结构特征. 浮游动物几乎全由象鼻溞(Bosmina)种类组成,反映了在鱼类捕食压力下浮游动物的特点.
上述不同区域近现代湖泊古生态的研究,为认识气候变化和人类活动背景下湖泊生态退化机理、预测未来变化趋势提供了重要依据,可从历史演化的视角,提出不同区域湖泊生态环境管理和恢复的策略.
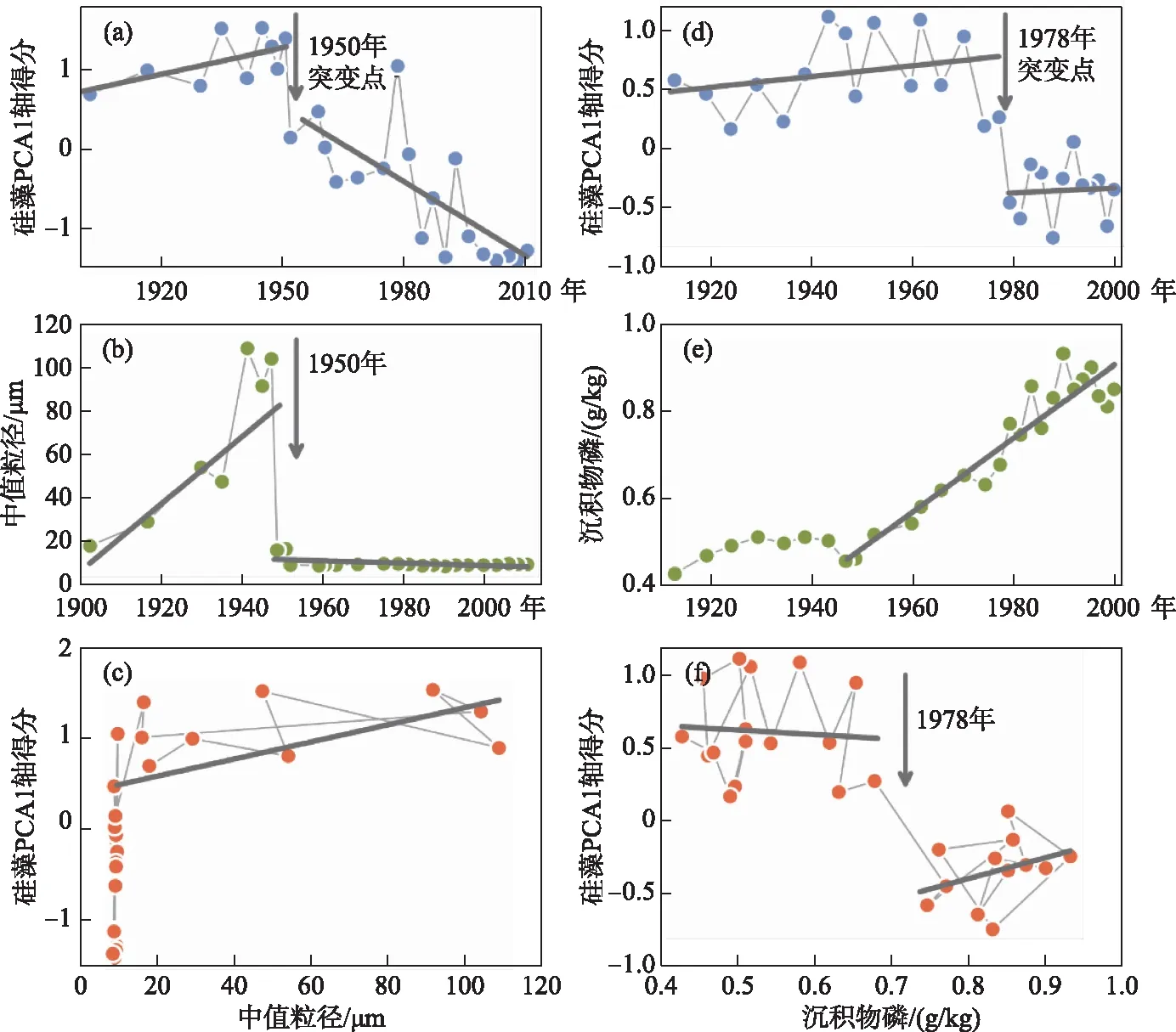
图6 长江中下游湖泊古生态揭示的生态系统突变模式(图中a~c为涨渡湖的相关结果,d~f为巢湖的相关结果. 其中:a和d为硅藻数据的PCA1得分,表示硅藻生态变化趋势;b为沉积物粒度中值粒径,表示湖泊水文变化;e为沉积物磷浓度,表示流域来源营养盐积蓄;c和f表示生态变化与驱动力的关系)Fig.6 Regime shift scenarios in Lake Zhangdu (a-c) and Lake Chaohu (d-f) of the Yangtze River Basin (a and d, the PCA1 scores of the diatom assemblages indicate the diatom ecological change; b, the median particle size of the sediments shows the hydrological change of the lake; e, phosphorus concentration in sediments indicates the accumulation of nutrient from catchment; c and f are the relationships of ‘pressure’ and ‘state’ in the respective lakes)
4 湖泊古生态在生态系统突变与弹性研究中的应用
湖泊生态系统对驱动力的线性和非线性响应是近年来国际湖泊古生态研究的热点之一,研究在揭示生态系统响应模式、临界转换、突变早期信号、生态系统结构和弹性等方面取得了可喜进展. 基于长江中下游多个湖泊古生态重建数据,Zhang 等[82]利用突变点检测方法,发现研究区近百年来的生态变化普遍存在两个突变点,分别对应了1950s-1960s和1980s-1990s的生物群落演替时间点. Xu等[83]进一步选择涨渡湖和巢湖,将硅藻生态变化序列与反映流域来源的沉积物磷浓度以及指示水文变化的沉积物粒度数据进行了对比分析(图6),发现1950年前后附生硅藻属种迅速增多和群落结构明显改变,与筑坝引起的快速水文条件改变相对应,认为这种由水文驱动的生态突变,属于典型的生态系统突变“外力驱动”模式,即生态系统突变响应驱动力快速变化的线性模式. 而在巢湖,沉积物硅藻记录揭示了长期营养富集造成1978年的生态突变. 这种突变是系统非线性响应驱动力(营养持续变化)的结果,属于临界转换模式[84-85]. 类似的现象同样发生在云南洱海、阳宗海等湖泊的硅藻和摇蚊记录中[86-87]. 如洱海古生态研究揭示的2001年系统突变与监测记录的藻类持续暴发的开始时间一致[86].
突变理论模型和试验研究认为,临界点到来之前生态系统主要表现为临界放缓现象,即生态系统逼近突变临界点时恢复速率会降低,并得出早期预警信号为系统状态的变率和自相关性同时升高的推断. 研究提出临界放缓理论可以用于指导生态系统的管理. 然而,这些研究缺乏来自自然生态系统的验证,备受学术界争议. Wang等[86]利用多元统计分析,首次从洱海古生态数据分析中,诊断出时间序列上,临界转换前系统状态频繁波动于不同稳态之间,表现为“闪烁(flickering)”特征. 数值分析推导出临界转换的早期预警信号指标,表现为状态变率的升高和自相关性的下降. 基于沉积钻孔多指标数据和监测数据的综合分析,进一步揭示了系统弹性损失和临界转换发生机制,该研究为利用湖泊古生态数据开展自然生态系统突变研究提供了典型案例,并为验证和丰富临界转换理论提供了重要依据.
生态系统弹性大小取决于生态系统结构稳定性,而早期信号只是生态系统弹性损失的间接指标. 近年来国内外学者在利用关联性、嵌套性和生物多样性等指标揭示生态系统弹性变化研究方面进行了有益的探索[88-91]. 古生态学数据再次展现了时间尺度长的优势. 例如,Wang等[91]根据我国青藏高原、西南山地和长江中下游地区273个湖泊、452个种的跨区域硅藻数据库,首次利用网络学手段进行了大数据分析,构建了硅藻网络图谱,建立了硅藻群落结构关联性指标(偏度、聚类系数、异质性)的定量重建模型,并结合不同区域5个典型湖泊沉积钻孔的硅藻分析数据,定量重建了这些湖泊近百年来的硅藻群落结构参数的演化轨迹[91]. 研究表明在时空尺度上,群落结构指标响应人类活动和营养梯度,在人类活动干扰较小的青藏高原和西南高山地区,湖泊生物群落结构关联性最低,反映了生态系统的高稳定性和高弹性;而在长江中下游地区,强烈的人类活动导致生态系统弹性损失最为严重.
不难看出,湖泊古生态研究在揭示湖泊生态系统演化、验证相关生态学理论中已表现出独特优势,但相关研究尚处于初步阶段,研究结果需要更多的案例进行支撑,也需要结合理论模型和实验室实验开展综合分析. 在未来的研究中,应该加强多学科的融合,特别是与生态学、系统科学、网络学等的交叉研究.
5 湖泊古生态研究展望
综上所述,我国湖泊古生态研究已在沉积物不同生物指标挖掘、环境定量化、湖泊生态环境演变和生态学理论研究等方面取得了长足的进展. 围绕湖泊生态系统演变及其对全球变化的响应主题,湖泊古生态仍需要通过加强基础和应用基础研究,通过技术创新和多学科交叉,深入理解不同时空尺度湖泊生态系统演化的特征、区域差异和动力学机制,发挥其在湖泊管理和治理中的指导作用.
5.1 加强古今结合研究,提高湖泊古生态重建的精度
尽管湖泊现代过程已为古生态数据解释提供了重要依据,并发展了基于古今结合的环境要素定量重建方法,但长期以来古生态和古湖沼学研究与湖沼学和生态学之间的脱钩现象依然存在. 古生态重建研究至今仍大多基于线性响应的思维模式,常常侧重对单一关键环境过程的推导,忽视潜在的“共线性要素”驱动效应,以及生物群落演替过程中食物网各营养级生物之间的关联性. 因此,需要加强日趋丰富的生物物种生理生态信息库及更大空间尺度上的现代生物-环境数据库的建立,从而有利于更准确、全面地揭示历史生态环境变化过程. 此外,目前工作中对许多生物指标的埋藏学特征仍缺少必要的研究. 如湖泊生物死亡后保存在沉积物中的生物遗存与现实的群落构成存在一定差异. 定量化这种群落差异及评估所造成的重建误差,需要通过各种观测实验,分析生物残体与母体的数量关系、沉积物中各物种之间的溶蚀差异等特征,提取丢失的数据信息,对古生态结果进行修正. 因此,在今后的研究中仍需加强各种现代过程与湖泊古生态研究的融合,通过现代过程数据的深入分析,揭示生物埋藏学特征、不同生物之间的关系以及它们对生境响应的敏感和差异特征,为精确的湖泊古生态重建服务.
5.2 重视多生物指标的综合和新指标技术的应用
近年来越来越多的沉积生物指标被提取,无疑为促进湖泊古生态研究提供了机遇. 但目前的古生态重建主要基于单生物指标,将多生物指标进行综合分析的研究偏少,而且对生态系统水平的演化研究涉及更少. 因此,加强多生物指标的综合分析应该是今后的一个必然趋势,这不仅可以对不同指标独立的逻辑解释起到相互验证和补充的作用,而且可以提供生态系统层面的信息,包括系统各要素相互作用、食物网结构和功能、生境和物种多样性以及系统的物质和能量循环等信息. 例如,沉积物中的环境DNA是最近古湖沼学研究中发展起来的一个新指标,在揭示个体生物、群落和种群结构组成及生态多样性的长期动态演化方面具有很大的潜力. 相对于生物化石指标,环境DNA指标的优势在于能够提高物种鉴定的高分辨率信息,提供较为完整的生物多样性信息,以及提取地层中的生物变化的信息,具有广阔的应用前景.
5.3 强调多胁迫因子对湖泊生态系统影响的定量区分
古湖沼研究的一个重要目标就是对湖泊生态环境演化的动力学机制进行解释. 在不同时间尺度上,湖泊生态系统演化对内部和外部强迫的响应是复杂的,往往受多个胁迫因素的影响,既存在独立的、协同增强的影响效应,也存在此消彼长的补偿效应. 利用古湖沼数据进行生态演化机制的解释,目前仍以定性的逻辑推理分析为主,即基于对不同生物与环境指标的响应机理认识,对关键环境变化与生物群落演替与驱动力关系进行推断. 近年来,借助于数值分析和模型技术,一些研究已经开始尝试生态环境-驱动力关系的半定量和定量研究,在揭示多胁迫因子相互作用对藻类群落影响机理、区分气候和人类活动影响贡献、量化生态系统中的捕食和竞争与营养循环相互作用关系等方面显示了很好的潜力[1, 92].
5.4 发挥湖泊古生态研究在评估生态系统服务长期变化中的作用
湖泊生态系统为人类社会提供多种有价值的生态系统服务,在维持人类福祉方面发挥着至关重要的作用. 然而,在强烈的人类活动干扰下,湖泊生态系统服务功能急剧退化. 在缺乏长期生态系统服务尤其是调节服务和支持服务变化的情况下,理解不同时期人类活动资源利用对生态系统服务变化产生的影响,以及各个生态系统服务间的动态相互作用关系,对于湖泊生态系统安全空间评估、自然资源的可持续利用和科学管理意义重大. 依据“千年生态系统评估(MA)”的分类标准,一些学者利用长江中下游典型湖泊古生态指标,对过去百年来生态系统服务指标(包括土壤维持力、沉积物质量、水质净化、生物多样性等)的变化过程进行了重建,并揭示了调节服务和供给服务长期变化过程的协同和权衡关系[93-94]. 进一步的研究可以通过构建湖泊流域社会-生态系统安全操作空间,深入理解生态系统与人类社会系统的长期动态变化关系,为区域可持续发展提供重要的理论依据.
5.5 加强湖泊古生态在湖泊管理和治理中的应用基础研究
面对不断加剧的湖泊生态环境问题,如何将过去环境变化研究更好地服务于湖泊环境的保护与治理,是当前湖泊古生态研究的一个重要目的. 古生态研究不仅可以提供湖泊受干扰前的环境和生态背景值,还可以从历史演变趋势上提供建立区域和具体湖泊生态质量演化、治理路径和阶段性治理效果评估等信息. 对于退化的湖泊生态系统,可以通过古生态重建和生态系统动态模拟相结合的手段,开展湖泊系统弹性损失机理、临界转换阈值和预警信号等研究. 一方面,通过界定湖泊生态系统的生态和环境本底,可对湖泊进行有针对性的保护和修复,有助于细化我国湖泊资源的开发与保护. 类似的研究计划在加拿大和美国已经开展多年. 在我国,由于不同区域湖泊的生物群落迥异,生态系统类型多样,目前已开展的湖泊古生态本底调查工作仍非常有限;另一方面,提高对湖泊生态系统不同演化过程机理的认识,提出针对性的保护和修复策略.
猜你喜欢
大自然探索(2023年7期)2023-11-14 13:07:36
课堂内外(小学版)(2023年9期)2023-10-11 14:39:26
天天爱科学(2022年9期)2022-09-15 01:12:54
天天爱科学(2022年4期)2022-05-23 12:41:48
当代水产(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
黑龙江水利科技(2020年8期)2020-01-12 06:26:00
少儿美术(快乐历史地理)(2019年4期)2019-08-27 00:51:40
阅读(低年级)(2018年4期)2018-05-14 17:39:57
上海建材(2018年1期)2018-04-18 12:15:16
