
白蚁肠道木质素降解菌分离鉴定及其降解特性
2020-08-28 16:36李锋黄庶识
生物技术通报 2020年8期
李锋 黄庶识
(1. 黔南民族师范学院化学化工学院,都匀 558000;2. 广西科学院 广西海洋天然产物与组合生物合成重点实验室,南宁 530007)
随着化石能源的过度消耗及环境污染的日益严重,寻求可再生、环境友好的替代品成为当今研究热点。植物每年通过光合作用生成资源丰富的木质纤维素,其中木质素是自然界中唯一可再生的芳香族聚合物。处理不当会对环境造成污染,如我国每年的农作物秸秆资源丰富,大部分以燃烧的方式进行处理,从而加剧了大气污染。此外,我国造纸工业产生的黑液废水主要成分为木质素,如果不进行处理直接排放,会污染水源甚至可能会对人类健康带来一定的威胁。因此,探索木质素有效分解并转化为高附加值化学品资源等方面的研究,对人类健康及环境可持续发展具有重要的意义。
自然界生物圈经过长期的进化和演变,存在着许多可以降解木质素的系统,如细菌、真菌、食木昆虫和食草动物等,它们都可以将木质素进行不同程度的分解。其中白蚁肠道消化系统被认为是高效木质纤维素转化反应器。相关研究发现,白蚁肠道内寄居着大量形态和密度都不同的细菌,每只白蚁肠道中约可达106-108个[1]。通过高通量测序和分离培养的方法已经对将近40多种白蚁进行了肠道细菌多样性分布研究。Lazuka等[2]在厌氧条件下对Nasutitermes ephratae肠道细菌进行富集,富集后的菌群多样性明显发生变化,经5次富集后得到菌群TWS培养12 d可以降解42%的麦秆,菌群TWS主要由拟杆菌属(Bacteroides)和毛螺科菌(Lachnospiraceae)的细菌组成。此外,白蚁肠道共生细菌受物理环境等因素的影响,其多样性分布情况存在较大差异。利用宏基因组对高等食木瓢白蚁(Bulbitermessp.)的前肠、中肠和后肠分别进行测序,发现前肠和中肠存在相似的多样性,而后肠中细菌多样性明显比前两者丰富。其中前肠和中肠优势菌为硬壁菌门的杆菌(Bacilli)、梭菌(Clostridia)和放线菌门的放线菌目细菌(Actinomycetales),而后肠主要为螺旋体门的螺旋体科细菌[3]。
在木质素降解菌分离纯培养方面,Zhou等[4]利用木质素作为碳源,从黑胸散白蚁肠道中分离到肠杆菌PY12(Enterobacter hormaechei)和芽孢杆菌 MX5(Bacillus licheniformis),二者表现出较高木质素过氧化物酶活力分别为278 U/L和256 U/L,对木质素过氧化酶进行异源表达,该酶表现出对多种染料有较强脱色能力。Suman等[5]从土白蚁(Odontotermes obesus)肠道中分离到一株特布尔希氏菌(Trabulsiellasp.),表现出较强木质素降解能力,其木质素降解产物检测到许多芳香族化合物,如3-甲氧基苯甲酸、邻苯二酚、邻苯二甲酸等。Tsegaye等[6]以碱木质素、羧甲基纤维素和木聚糖为碳源,从麻头堆砂白蚁(Cryptotermes brevis)肠道分离到芽孢杆菌(Bacillussp.)BMP01和水稻苍白杆菌(Ochrobactrum oryzae)BMP03,其中芽孢杆菌具有较强的纤维素和木聚糖水解的能力,水稻苍白杆菌表现出较高的漆酶和木质素过氧化酶酶活力。此外,以水稻秸秆为碳源培养14 d后,水稻苍白杆菌可去除秸秆中53.74%木质素[7]。
以上这些研究表明白蚁肠道是储存木质纤维素降解细菌的资源库,具备挖掘新颖木质素降解菌及酶资源的潜力。本研究以碱木质素为唯一碳源,筛选分离白蚁肠道中潜在的木质素降解菌,对其进行分子生物学鉴定,并进一步解析其木质素的降解特性,旨在为木质素生物降解转化提供新颖微生物资源和理论依据。
1 材料与方法
1.1 材料
1.1.1 样品采集 高等白蚁采集于中国科学院西双版纳热带植物园次生雨林,经兵蚁形态特征及COⅡ基因序列鉴定为巴基斯坦小白蚁(Microtermes pakistanicus)。
1.1.2 培养基 分离培养基(g/L):K2HPO41,KH2PO41,(NH4)2SO42,MgSO40.2,CaCl20.1,FeSO40.05,MnSO40.02,CuSO40.001,ZnSO40.001,tryptone 0.05 琼脂粉 15,碱木质素1.0。
染 料 脱 色 固 体 培 养 基(g/L):K2HPO41,KH2PO41,(NH4)2SO42,MgSO4·7H2O 0.2,CaCl20.1,FeSO4·7H2O 0.05,MnSO4·H2O 0.02,CuSO4·5H2O 0.001,ZnSO4·7H2O 0.001,tryptone 0.05,琼 脂 粉15,pH 7.2,灭菌后,再分别加入100 mg经0.22 μm过滤灭菌的染料(苯胺蓝和天青B);
MSM液 体 培 养 基(g/L):K2HPO41,KH2PO41,(NH4)2SO42,MgSO4·7H2O 0.2,CaCl20.1,FeSO4·7H2O 0.05,MnSO4·H2O 0.02,CuSO4·5H2O 0.001,ZnSO4·7H2O 0.001,tryptone 0.05,碱木质素1.0,pH 7.0左右。
以上所述培养基均为115℃,灭菌15 min。
1.2 方法
1.2.1 菌株分离纯化及鉴定 取巴基斯坦小白蚁的工蚁30头,用75% 酒精对白蚁表面消毒后,在中性PBS缓冲液中漂洗2-3次。于无菌条件下,解剖白蚁肠道置于无菌0.9% 生理盐水中,然后将其研磨成匀浆状态。肠道匀浆液用无菌生理盐水梯度稀释后,取100 μL溶液涂布于分固体分离培养基的平板上,置于30℃,培养2-5 d。待有单菌落长出后,挑取单菌落,在液体LB培养基过夜培养,重新划线于分离培养基平板中,进行多次划线分离培养,直至获得纯菌株。
将单克隆接种于LB培养基,过夜培养,取1 mL培养液于离心管中,12 000 r/min离心2 min后弃上清液,菌体经无菌水漂洗后,用细菌基因组试剂盒进行DNA提取。获得细菌基因组后进行16S rRNA 基因扩增,具体扩增条件参考文献[8]。PCR产物经1% 琼脂糖凝胶电泳检测后,委托上海生工对细菌基因序列进行双向测序。将测序完成的16S rRNA 基因序列拼接后,提交至NCBI网站的GenBank数据库,并进行BLASTn(www.ncbi.nlm.nih.gov/BLAST)同源性相似度比对。下载比对序列同源性较高的菌株序列,利用MEGA 5.05 软件计算序列相似度,同时用Neighbor-Joining 法构建菌株系统发育树,确定物种的分类地位。
1.2.2 细菌胞外酶定性检测 利用染料脱色来评价细菌是否能够产胞外分泌酶的能力,通过检验细菌对染料的脱色能力,达到对细菌进一步复筛目的。挑取单克隆于LB培养基中过夜培养后,接种环取菌液划线于染料固体培养基上,置于30℃恒温培养箱中培养,每天观察菌株的生长情况以及是否有脱色圈产生。
1.2.3 细菌木质素降解率测定 在有氧条件下,细菌接种于MSM液体培养基后,每隔24 h 取样。细菌生长曲线按照如下步骤测定:取2 mL发酵液,经10 000 r/min 离心5 min后,弃上清液,底物菌体用去离子水重悬后,在波长600 nm处测定吸光度OD值,去离子水为空白对照。培养基中碱木质素浓度以溶液中化学需氧量(COD)为考察指标,采用重铬酸钾法测定COD(GB11914-89),离心后的上清液经一定倍数稀释后,经消解仪消解样品后,利用5B-3B型多功能COD快速测定仪测定COD值。以上实验均为3个平行试验。
1.2.4 细菌木质素降胞外酶活力测定 细菌降解木质素胞外漆酶(Lac)、木质素过氧化物酶(Lip)和锰过氧化物酶(MnP)测定具体操作步骤参考文献[9]。
1.2.5 扫描电子显微镜观察 未接菌的培养液作为对照试验组和经细菌处理7 d后的一定量发酵液经冷冻干燥后,取木质素粉末经乙醇洗涤去除残留物,烘干后备用,分别取少量干燥木质素样品涂布于双面导电胶上,经喷金处理后,进行JSM-7800F场发射扫描电子显微镜(日本电子株式会社)观察和拍照。
1.2.6 傅里叶变换红外光谱分析 木质素红外光谱(Nicolet nexus 470 傅里叶变换红外光谱仪,美国尼高力仪器公司)采集采用溴化钾压片法,分别取一定量经过干燥的接菌和未接菌的木质素样品,木质素样品和溴化钾按质量比为1∶100混合均匀后,将其研磨至成粉末,制备成厚度均匀薄片后,采集红外图谱,以纯溴化钾为背景,波长扫描范围4 000至400 cm-1,分辨率为4 cm-1。
1.2.7 热解特性分析 木质素热稳定分析采用TGA4000热重分析仪(Thermo-gravimetric annlyzer,TGA,美国珀金埃尔默),取大约2 mg 未经细菌处理和经过细菌处理的木质素样品,分析放入样品盘后,温度从50℃开始升温至800℃结束,升温速度为20℃/min,氮气流速为20 mL/min。每个实验重复3次。
2 结果
2.1 木质素降解菌筛选分离与鉴定
2.1.1 木质素降解菌筛选分离 待固体分离培养基上长出单菌落后,挑取单菌落继续进行划线分离多次培养后,将单菌落接种于固体分离培养基,挑取生长速度快和菌落个体较大的细菌进行编号并保藏。为了直观评价细菌分泌胞外木质素降解相关酶的能力,本研究采用较为常用的天青B和苯胺蓝作为底物,对细菌脱色能力进行评价。将菌株接种于染料培养基过夜培养后,发现菌株MP-132能够使天青B和苯胺蓝脱色(图1)。由图1可知,菌株MP-132在染料诱导的情况下,可以分泌胞外酶将其脱色。染料脱色只能定性判断产酶情况,但是,该菌分泌哪种类型的酶及其酶活大小还需进一步分析。
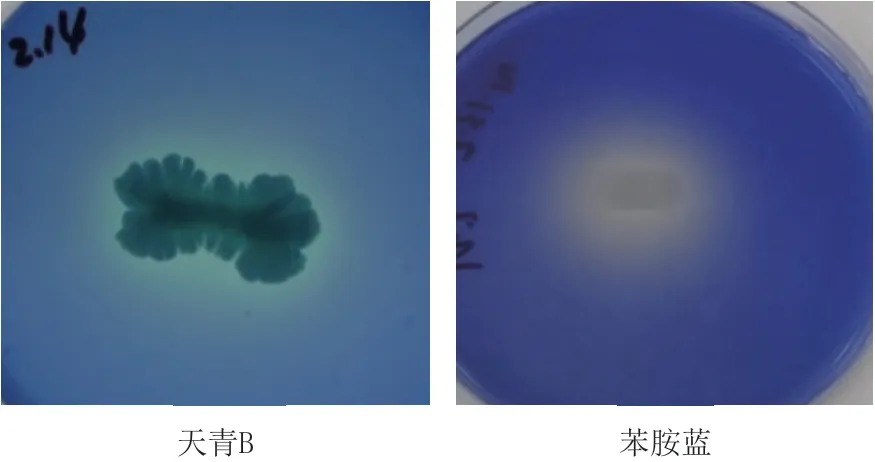
图1 菌株MP-132的染料脱色评价
2.1.2 木质素降解菌的16S rRNA 基因序列分析 双向测序获得菌株MP-132的16S rRNA 基因序列,经RNAMAN序列拼接后,提交至GenBank 数据库,序列登录号为MF455198,同时在NCBI数据库中进行序列同源性比对,选取BLAST中相似度较高的细菌序列,并下载相关序列后,利用MEGA5.05计算细菌序列相似度,同时采用其中的Neighbor Joining法构建细菌系统发育树。如图2所示,MP-132与Raoultella ornithinolyticastrain ATCC 31898在同一个分支上,序列相似性为99%。因此,将该菌株初步命名为Raoultella ornithinolyticaMP-132。
2.2 木质素降解菌的生长及酶活力分析
2.2.1 木质素降解菌的生长情况及降解率测定 为了解菌株MP-132在液体碱木质素中的生长及木质素降解情况,将细菌分别接种于以碱木质素为唯一碳源的无机盐培养基中,因碱木质素是培养基中唯一提供的有机物质,所以采用COD去除率作为指标,评价碱木质素降解率。由图3可知,菌株MP-132前4 d菌体呈现迅速生长趋势,第4天后菌体OD值达到0.87,随后OD曲线呈现平缓增长趋势,第6天OD值增长到0.913,OD数值变化不大。液体培养基中COD浓度变化曲线,前两天降低速度缓慢,第2天开始继续开始下降,可能是与细菌活性和浓度有关,直到第7天培养基中COD浓度为842.7 mg/L,降解率达到53.2%,后期COD去除率变化缓慢,由此表明降解产物的累积使菌体代谢活性受到影响。
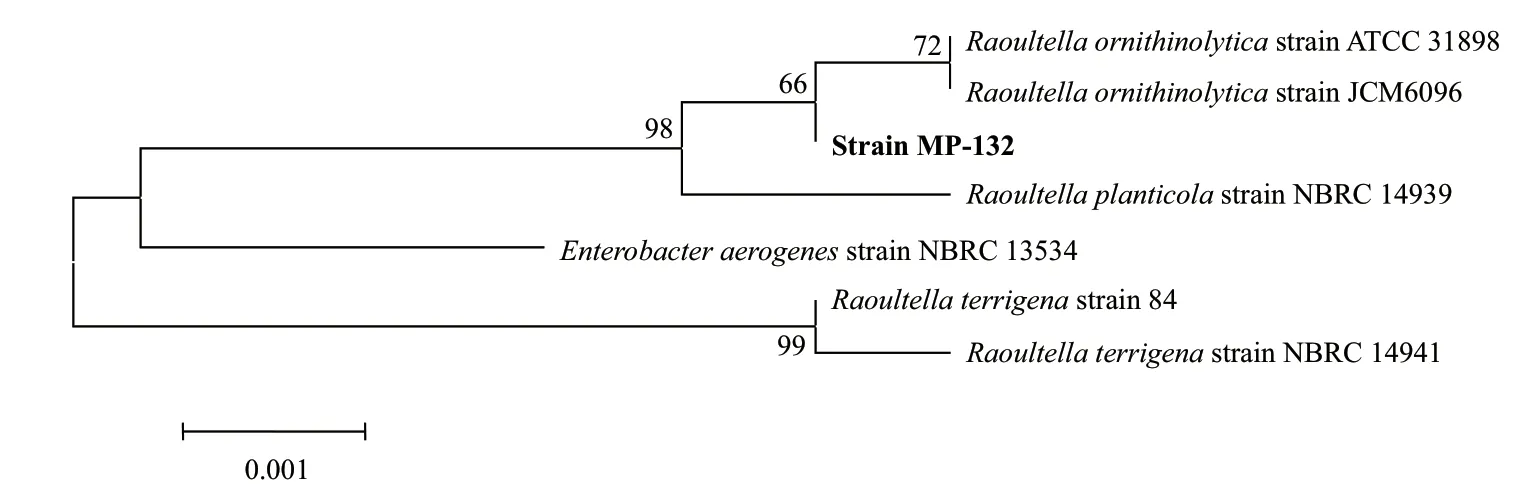
图2 菌株MP-132的16S rRNA基因序列系统发育树
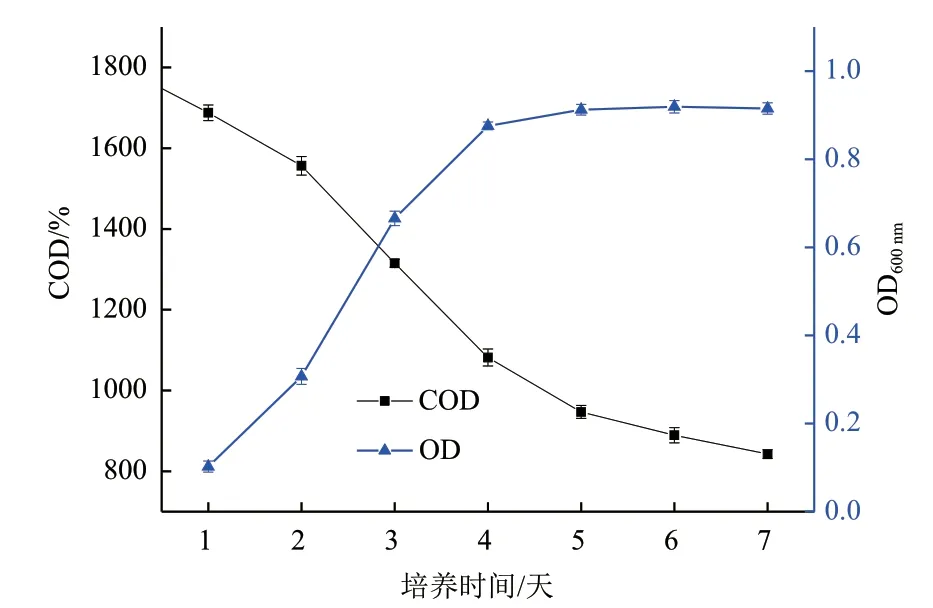
图3 菌株MP-132在碱木质素培养基中的生长及COD去除率
2.2.2 木质素降解菌的酶活力测定 由图4可知,细菌在碱木质素培养基生长过程中没有检测到锰过氧化物酶活力,只检测到漆酶和木质素过氧化物酶。由木质素过氧化物酶的增加速率趋势分析发现,培养2 d后LiP增长速率放缓,第4天LiP达到最大值为105.6 U/L,随着培养时间增加,其酶活力开始出现下降趋势,可能是由于细菌衰老,其代谢活力降低,从而导致酶活力出现下降趋势。在木质素降解过程中,Lac酶活力在第2天达到27.53 U/L,即使随着培养时间的增加,Lac酶活力在第3天达到最大值32 U/L。菌株MP-132所产LiP和Lac酶活力明显低于以前报道的白腐菌和其他类细菌所产的酶活,可能是受发酵参数影响,如温度、溶氧量和pH等,后续研究应该对其发酵过程进行条件优化。
2.3 菌株MP-132的降解特性

图4 木质素降解过程中菌株MP-132酶活曲线
2.3.1 木质素微观结构分析 菌株MP-132处理前后的碱木质素形貌进行扫描电镜观察分析比较。如图5中所示,图5-A为在放大倍数为1 000倍状态下,未处理碱木质素呈现的结构状态,其呈球形或者块状,且表面相对光滑,完整致密。图5-B为细菌处理后的碱木质素在放大倍数1 000倍状态下,呈现的碱木质素微观结构。经过细菌降解后的碱木质素已经没有规则的球形或者块状结构,且呈现疏松多孔的表面,甚至出现许多片状颗粒,它们的直径都小于10 μm。经对比木质素降解前后微观结构变化情况,证实细菌都可以通过分泌胞外酶使碱木质素微观结构遭到严重的破坏。

图5 木质素降解前后的微观结构
2.3.2 木质素的FTIR分析 由图6可知,是经菌株MP-132降解前后的碱木质素的红外光谱图。由图可知,经过细菌降解的木质素的特征峰强度减小或者发生漂移,木质素特征峰变化也是主要集中于1 800 /cm-800/cm的指纹区。1 596 /cm和1 513/cm处吸收峰归属于芳香环骨架振动和C-H变形振动,1 456/cm和 1 424/cm处吸收峰是由于甲氧基的甲基C-H不对称变形,1 374/cm处吸收峰归属于甲基和酚羟基的C-H的伸缩引起。此外还存在1 268/cm、1 217/cm处吸收峰是典型愈创木酚基结构(G),1 126/cm处是典型紫丁香基(S)的吸收峰。1 082、1 033、967 和880/cm处吸收峰分别归属于Cβ和脂肪醚的C-O变形,以及苯环C-H面内变形振动、苯环C-H面外伸缩振动和C-H变形振动。细菌降解后木质素的多个吸收峰消失,如1 374、1 151、1 082、967、880、854和 816/cm, 以 及1 596、1 513、1 456、1 424、1 268和 1 033/cm处的几个吸收峰相对吸收强度减小,上述多个吸收峰的变化表明菌株MP-132使碱木质素的侧链、C=O、C-O和芳香环骨架结构遭到破坏。
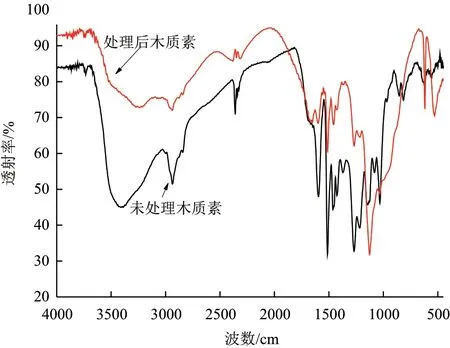
图6 木质素降解前后的红外光谱图
2.3.3 木质素的热解特性分析 碱木质素热解性质与其芳香环骨架结构及在不同温度下化学键裂解所需要的键能有着密切的关系,可以作为评判木质素结构变化的考核参数。在升温速度为20℃/min条件下,降解前后的木质素样品进行热重分析,得到TG曲线。由图7中TG曲线可知,降解前后的木质素的热稳定性存在较大差异,主要表现在热分解反应的起始和终止温度、失重速率和失重峰值点等。细菌降解前后木质素样品均呈现两个重量损失阶段。第一阶段发生在100-180℃范围内的重量损失;细菌处理碱木质素第二阶段重量损失从180℃开始,一直升温至640℃终止,最终得到剩余残渣质量达到60%。而未处理碱木质素在从200℃开始一直升温至800℃,有明显的重量损失,最终剩余残渣为29.3%。
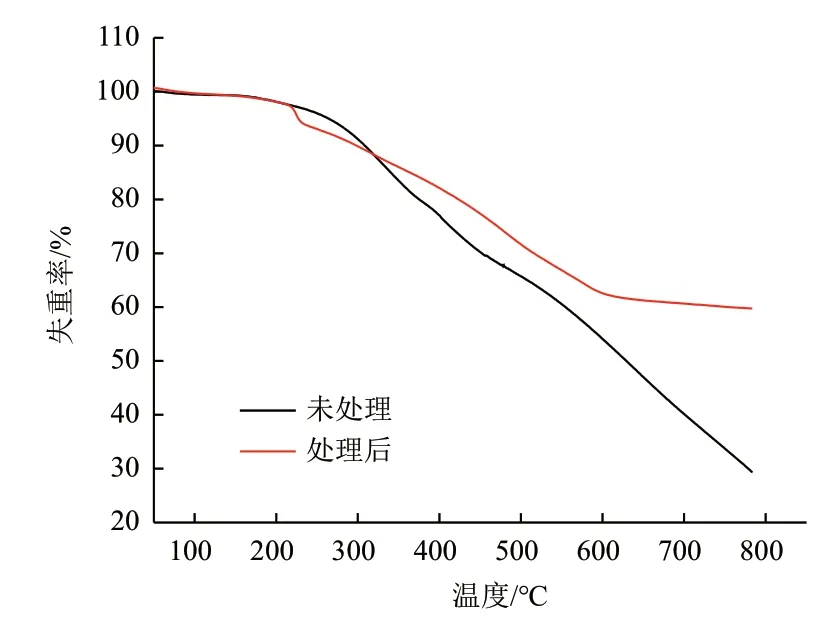
图7 木质素降解前后TG曲线
3 讨论
木质素由多种化学键构成的大分子聚合物,在自然界中的储量仅次于纤维素,如将其高值化利用对改善能源结构具有重要的意义。细菌具有生长速度快,适应环境能力强和基因易操控等优点,备受国内外研究者的青睐。近年来,关于细菌降解木质素研究逐渐增多,同时对木质素降解机制也逐渐补充完善。如Zhu等[10]从南海深海海泥中筛选到一株嗜碱菌Bacillus ligniniphilusL1,通过基因组和蛋白组测序发现该菌拥有木质素主要代谢途径:苯甲酸代谢途径、龙胆酸代谢途径和β-酮己二酸代谢途径。Higuchi等[11]从Sphingobiumsp. YK-6基因组中鉴定到两个新颖基因编码的酶,可以催化裂解芳基醇-β-芳醚结构,并发现存在可以将木质素转化为高附加值化学品(2-吡喃酮-4,6二甲酸)的HPV代谢途径。以上研究表明,挖掘自然界中木质素降解细菌具有较大潜力。
Raoultella属细菌是广泛存在生态环境中的一类革兰氏阴性细菌,具备降解多种有机污染物的能力[12-14]。本研究从高等食木白蚁肠道中分离得到一株可降解木质素的菌株MP-132,经分子生物学鉴定为解鸟氨酸拉乌尔菌。进一步对其降解木质素试验分析,培养7 d后其木质素降解率可达53.2%,明显高于已报道的Burkholderiasp. H1[15],其降解率可能受环境条件,如温度、溶氧量和pH等,后续需优化培养条件提高木质素降解率。在木质素降解变化的化学官能团方面:如1 374/cm处的吸收峰消失,表明降解后的木质素可能是发生侧链被氧化、脱去甲基以及芳香环骨架受到破坏[16]。在1 142/cm处消失的特征峰,其归属于醚键的伸缩振动,由此表明细菌在一定程度上对木质素的主要化学键β-O-4造成了破坏。其次,木质素降解前后热解特性发生变化,表明样品中化学键和官能团含量经过细菌处理以后发生改变。在200-350℃范围内,处理过的碱木质素拥有较高的热分解速率(与对照样品相比),这一阶段的热分解主要是侧链和一些键能低的化学键的断裂,如侧链的β-γ碳碳键、β或γ位的OH、β-O-4的C-O键等[17]。在350-400℃范围内,这一阶段主要是侧链被氧化,如脂肪羟基的羧基化或者羰基化和侧链的脱羟基[18]。当温度高于400℃,木质素热分解主要归因于木质素芳香环骨架的C-C键(主要是5-5'和 β-β')的裂解以及脱甲氧基[19]。
此外,在木质素降解过程中未发现锰过氧化物酶活力,可能是由其他种类同工酶发挥着MnP的作用。并且已有研究表明由于木质素结构复杂,其降解过程需要多种酶参与完成,如细胞色素P450单加氧酶、过氧化氢酶、多功能氧化酶、乙二醛氧化酶、芳醇氧化酶和葡萄糖氧化酶等[20-21]。木质素结构复杂,其降解酶系不能只通过评价目前研究中所报道的3种酶,后期应利用现有先进技术手段(如分泌蛋白组学)对其降解酶系进行宏观性评价。鲍文英等[22]从土壤中分离得到一株Raoultella ornithinolyticaS12进行基因组测序,其基因组中存在大量与木质素降解相关酶类的编码基因,如漆酶、过氧化物酶、超氧化物歧化酶、儿茶酚-1,2-双加氧酶和原儿茶酸-3,4-双加氧酶等,这些基因在以碱木质素为碳源的培养条件下的表达量,均高于以葡萄糖为碳源培养的。菌株MP-132与Raoultella ornithinolyticaS12属于同一种细菌,其木质素降解机制可能存在较大的相似性,但两个细菌来源不同,其潜在的差异性还需进一步利用先进系统生物学和先进化学分析手段进行研究,为完善细菌降解木质素机制提供科学理论依据。
4 结论
本研究从白蚁肠道筛选分离到一株可降解木质素的菌株MP-132,经鉴定为解鸟氨酸拉乌尔菌(GenBank登录号MF455198)。该菌能够利用碱木质素为唯一碳源生长,定性和定量检测其可分泌胞外漆酶和木质素过氧化物酶,并通过观察木质素降解前后的微观形貌发生明显变化。此外,细菌生物处理后木质素的官能团结构和热解特性也发现改变。在以碱木质素为碳源培养过程中,在第7天时,其降解率可达53.2%,LiP和Lac酶活力分别达到最大值为105.6 U/L和 32 U/L。本研究系统研究了拉乌尔属细菌降解木质素的特性,该菌具备木质素降解能力,为丰富木质素降解微生物提供了优良菌株资源。
猜你喜欢
草地学报(2022年3期)2022-03-28
今日农业(2021年11期)2021-11-27
上海包装(2019年8期)2019-11-11
文萃报·周二版(2018年22期)2018-09-18
中学生数理化·高一版(2018年6期)2018-07-09
故事作文·高年级(2017年11期)2017-11-15
天津造纸(2016年1期)2017-01-15
中国酿造(2016年12期)2016-03-01
中国造纸学报(2015年1期)2015-12-16
科学启蒙(2015年8期)2015-08-07
