
Analysis on the Expression of TaMOR Gene in Two Different Spring Wheat Varieties at Different Years
2020-08-15 13:35XiaLIQianglinWANGChaoranLIHuafangZHUBeixiZHUXinLIUMeifengFULijunZHULiZHU
农业生物技术(英文版) 2020年4期
Xia LI Qianglin WANG Chaoran LI Huafang ZHU Beixi ZHU Xin LIU Meifeng FU Lijun ZHU Li ZHU
Abstract [Objectives] The TaMOR gene is a gene that affects the initiation and growth of the secondary roots of wheat, but the expression patterns in different parts of the wheat root system and the differences in expression in different varieties are not clear. This study aimed to investigate the expression of the TaMOR gene in the seminal roots, secondary roots and root base.
[Methods] Real-time fluorescence quantitative PCR technology was used to analyze the relative expression levels of the TaMOR gene in seminal roots, secondary roots and root base of seedlings of ancient variety Monkhead and modern variety Longchun 35.
[Results] There was no significant difference in the number of seminal roots between Longchun 35 and Monkhead, and the numbers of seminal roots of the two varieties did not change significantly during the three sampling periods. The number of secondary roots and shoot dry weight of Longchun 35 were significantly higher than those of Monkhead, and the number of secondary roots and shoot dry weight of both varieties increased with the sampling time point. The root dry weight of Monkhead increased with the sampling time, while Longchun 35 showed the largest value at the second time. The fluorescence quantitative PCR results showed that for 13-day seedlings, the relative expression of the TaMOR gene in root base was significantly higher than that in the seminal roots and secondary roots. There was no significant difference in the relative expression of gene TaMOR in the root system of Monkhead and Longchun 35.
[Conclusions] The root allocation of gramineous crops decreases with the breeding years, and the difference in gene TaMOR expression level needs further study.
Key words Spring wheat; Secondary root; Primary roots Root; TaMOR gene
As one of the worlds major food crops, wheat provides about 1/5 of the energy source for the worlds population[1]. Increasing wheat production potential is one of the effective solutions to the persistent food security problem[2]. The root configuration of crops directly affects yield, revealing that genes related to root development and gene expression patterns are of great significance to the genetic improvement of crops[3]. A large number of studies have shown that for the graminaceous crop wheat, the number and biomass of secondary roots have a very direct and important relationship with yield potential[4-5].
Several genes in rice that affect the initiation of secondary roots and the appearance of secondary roots have been cloned[6-7], and these genes ultimately affect the number and biomass of secondary roots in rice[7]. Among them, the rice CRL1/ARL1 gene specifically affects the growth of secondary roots, and the growth of the secondary roots of its mutant plants is hindered, but the aboveground part does not show abnormalities[8-9]. Jing Ruilians research group of the Institute of Crop Sciences, Chinese Academy of Agricultural Sciences first cloned the homologous gene TaMOR of CRL1/ARL1 in wheat, and found that rice transgenic lines overexpressing TaMOR produce more secondary roots and higher population yields[10]. However, the expression pattern of this gene in different parts of wheat roots is not clear. Because the expression of this gene is closely related to the yield potential, it is not yet known whether the agricultural breeding process that has targeted high yield has affected the expression of this gene. Therefore, in this study, two spring wheat varieties widely planted in different breeding years were selected to investigate the expression of the TaMOR gene in the seminal roots, secondary roots and root base.
Materials and Methods
Plant materials
The ancient variety, Monkhead, was a local variety in the arid area of central Gansu Province, which was widely planted between 1940 and 1970. The modern variety Longchun 35, is a variety bred by Gansu Academy of Agricultural Sciences in 2016 and has been widely promoted.
Pot planting experiment
The spring wheat seeds were sterilized with 2% sodium hypochlorite, placed in a refrigerator at 4 ℃ for 24 h, and then germinated in a 25 ℃ incubator. The germinated seedlings were transplanted into pots filled with paddy soil, with 7 plants per pot and 9 pots for each variety. The flower pots were randomly arranged during growth. When the seedlings grew to the tillering stage, the seminal roots, secondary roots and root base tissue were taken respectively, quickly frozen with liquid nitrogen, and stored in a refrigerator at -80 ℃. When taking root tissues at the tillering stage, the numbers of seminal roots and secondary roots were counted every 5 d, and the root system and shoot were dried and weighed, for the dry weights of the root system and shoot.
Analysis of relative expression of the TaMOR gene
In order to detect the expression pattern of the TaMOR gene in the root system, 13-day seedlings with basically the same growth trend were selected, and such three root tissue parts as the seminal roots, secondary roots and root base of the two varieties were taken. Total RNA was taken from different root parts using the general plant total RNA extraction kit from Tiangen, referring to the kit instruction. The total RNA of the TaMOR gene with good quality was reverse transcribed into cDNA using Tiangens reverse transcription kit, and the reverse transcribed cDNA was diluted 5 times as the template for quantitative PCR detection. The wheat TaActin gene was used as the internal reference gene, and the used primer sequences were as follows: TaActin-RT-F (5′-CTCCCTCACAACAACAACCGC-3′)/TaActin-RT-R(5′-TACCAGGAACTTCCATACCAAC -3′). To detect the expression level of the target gene in different tissues, the quantitative PCR primers used were TaMOR-RT-F(5′-GTCTTTGCGCCCTACTTCTG-3′)/TaMOR-RT-R(5′-TCATGACCTGCTGCTGGAG-3′). The primers were synthesized by GenScript Biotech Corp. Fluorescence quantitative PCR was performed using the SYBR Premix Ex TaqTM GC kit, and the reaction was carried out according to the procedure provided by the kit: 95 ℃, 30 s, and 35 cycles of 95 ℃, 5 s and 58 ℃, 30 s. After the reaction was completed, the quantitative PCR data was exported, and the expression level of the target gene was calculated by the 2(-ΔΔC(t)) method.
Results and Analysis
Variation trends of the numbers of seminal roots and secondary roots and biomass at the seedling stage
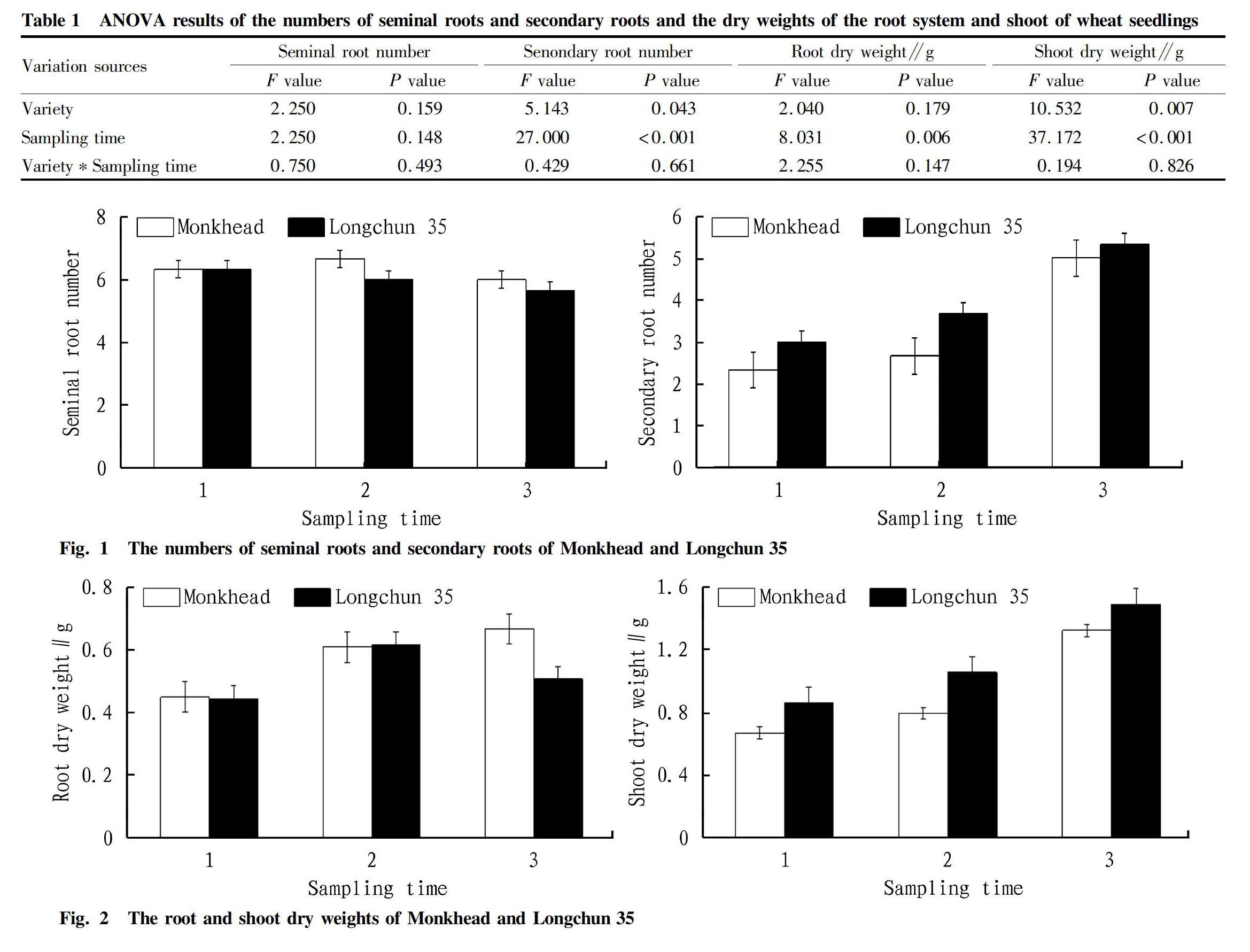
Comparing the modern variety Longchun 35 with the ancient variety Monkhead, there were no significant differences in the number of seminal roots, and the numbers of seminal roots of the two varieties did not change significantly during the three sampling periods (Table 1, Fig. 1). The number of secondary roots of Longchun 35 was significantly higher than that of Monkhead, and the numbers of secondary roots of both varieties increased with the sampling time point (Table 1, Fig. 1). The biomass data showed that the root dry weight of Monkhead at the seedling stage increased with the sampling time point, while the root dry weight of Longchun 35 increased at the second sampling time point, but then decreased at the third time point (Table 1, Fig. 2). The shoot dry weight of Longchun 35 was significantly greater than that of Monkhead, and the shoot dry weights of both varieties increased with the sampling time point (Table 1, Fig. 2).
Expression level of gene TaMOR in different root parts
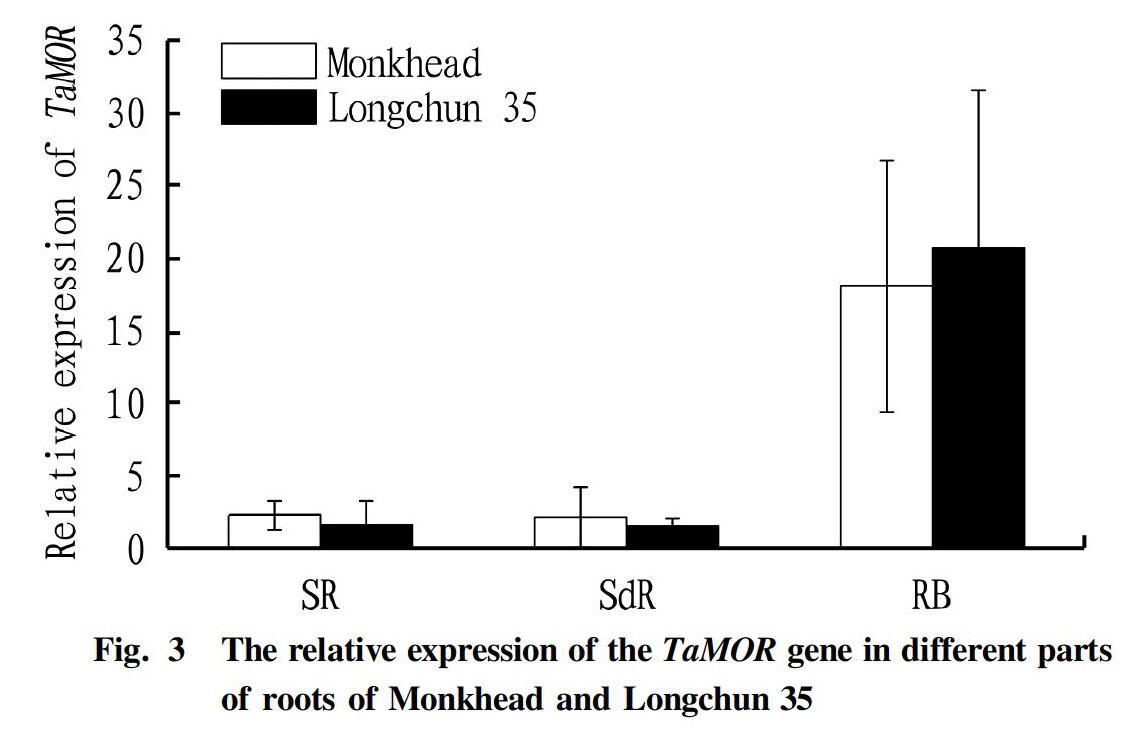
The fluorescence quantitative PCR analysis showed that for 13-day seedlings, the relative expression levels of TaMOR in seminal roots and secondary roots were not significantly different, but the relative expression in the root base was significantly higher than those in the seminal roots and secondary roots (ANOVA, F=21.797, P<0.001; Fig. 3). No matter which root part it was in, there was no significant difference in the relative expression of gene TaMOR in the roots of Monkhead and Longchun 35 (ANOVA, F=0.026, P=0.875).
Conclusions and Discussion
The root base of gramineous crops is the generation position of secondary roots[8-10]. The quantitative PCR results showed that the relative expression of the TaMOR gene in the root base of 13-day seedlings was higher than that in the seminal roots and secondary roots. This result supports the hypothesis that the TaMOR gene participates in the initiation of secondary roots. The TaMOR gene may be one of the important factors determining the number of secondary roots. Moreover, the high expression of the TaMOR gene in the root base of 13-day-old seedlings was consistent with the increase in the number of secondary roots we observed in the seedling stage with the sampling time point. Other studies have also shown that the expression of TaMOR in the initiation part of secondary roots and lateral roots is significantly higher than other tissues, and the expression pattern is consistent with the root formation time[10].
Our results showed that for Monkhead, the trends of the number of secondary roots and the dry weight of the root system were the same, but for Longchun 35, the dry weight of the root system increased first and then decreased; and at the third time point, the root dry weight of Longchun 35 was lower than that of Monkhead. This result showed that although the number of secondary roots increased at the seedling stage, the root development pattern and configuration of the modern variety Longchun 35 was different from the ancient variety Monkhead. It could be seen that as the seedlings grew, Longchun 35 might show relative restraint in root allocation. In fact, many studies have confirmed that the root allocation of gramineous crops decreases with the breeding eras[11-15].
The quantitative results of this study did not show differences between the varieties, which might be because the relative expression of the TaMOR gene at the seedling stage of the two varieties did not differ, or because the number of selected varieties was too small to be representative. Therefore, the variety differences in TaMOR expression level need to be further studied. In addition, the quantitative data in this study was only from one sampling time point, and it was impossible to observe how the TaMOR gene expression changes with wheat development. Therefore, follow-up studies should take multiple time points to further study the expression pattern of the TaMOR gene.
Agricultural Biotechnology2020
References
[1] FAOSTAT, Statistics database. Http:∥www.Fao.Org/statistics/databases/en/.
[2] BROWN LR. World on the edge: How to prevent envirenmental and economic collapse[M]. New York: W. W. Norton, 2011.
[3] OYANAGI A, NAKAMOTO T, MORITA S. The gravitropic response of roots and the shaping of the root system in cereal plants[J]. Environmental and Experimental Botany, 1993, 33(1): 141-158.
[4] MANSCHADI AM, CHRISTOPHER J, DEVOIL P, et al. The role of root architectural traits in adaptation of wheat to water-limited environments[J]. Functional Plant Biology, 2006, 33(9): 823-837.
[5] WASSON AP, RICHARDS RA, CHATRATH R, et al. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops[J]. Journal of Experimental Botany, 2012(63): 3485-3498.
[6] HOCHHOLDINGER F, TUBEROSA R. Genetic and genomic dissection of maize root development and architecture[J]. Current Opinion in Plant Biology, 2009(12): 172-177.
[7] MAI CD, PHUNG NT, TO HT, et al. Genes controlling root development in rice[J]. Rice, 2014(7): 1-11.
[8] INUKAI Y, SAKAMOTO T, UEGUCHI-TANAKA M, et al. The Crown rootless1 gene in rice is essential for crown root formation and is a target of an auxin response factor in auxin signaling[J]. The Plant Cell, 2005(17): 1387-1396
[9] LIU H, WANG S, YU X, et al. ARL1, a LOB-domain protein required for adventitious root formation in rice[J]. Plant Journal, 2005(43): 47-56.
[10] LI B, LIU D, LI Q, et al. Overexpression of wheat gene TaMOR improves root system architecture and grain yield in Oryza sativa[J]. Journal of Experimental Botany, 2016(67): 4155-4167.
[11] ZHANG R, ZHANG DY. A comparative study on root redundancy in spring wheat varieties released in different years in semi-arid area[J]. Chinese Journal of Plant Ecology, 2000, 24(3): 298-303. (in Chinese)
[12] LI L. Genetic analysis of wheat root morphology and physiological traits related to drought resistance[D]. Beijing: Chinese Academy of Agricultural Sciences, 2017. (in Chinese)
[13] ZHU L, ZHANG DY. Donalds ideotype and growth redundancy: A pot experimental test using an old and a modern spring wheat cultivar[J]. Plos One, 2013, 8(12): e70006-e70006.
[14] AZIZ MM, PALTA JA, SIDDIQUE KHM, et al. Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in Australian wheat varieties[J]. Plant and Soil, 2017, 413(1-2): 181-192.
[15] ZHU L, XI N, ZHANG DY. Lower sensitivity in responses to root competition and soil resource availability in a new wheat cultivar than in an old wheat landrace[J]. Plant and Soil 2020(450): 557-565.

- 农业生物技术(英文版)的其它文章
- Investigation on Agronomic Characters of Dwarf Mutant 778 in Broomcorn Millet (Panicum miliaceum L.) and Analysis of Its Sensitivity to GA
- Construction of Technology System on Development and Repropagation in Vitro of Several Cultivars in Pear
- Construction of Camellia oleifera Cultivation Standardization System
- Nutrients Determination in Nuts from Different Torreya grandis Cultivars
- Photosynthetic Physiological Response to Drought Stress of Populus euphratica at Different Ages in Minqin
- Effects of UV-B Radiation on the Activity of Antioxidants in Flue-cured Tobacco Leaves