
转基因棉生长前期不同叶位叶片对甜菜夜蛾种群增长的影响
2020-08-08 06:46:14杨旸谢修庆苏宏华杨益众
棉花学报 2020年3期
杨旸,谢修庆,苏宏华,杨益众*
(1.扬州大学园艺与植物保护学院,江苏 扬州225009;2. 江苏省连云港市农业农村局,江苏 连云港222023)
甜菜夜蛾(Spodoptera exiguaHübner),又名贪夜蛾、玉米叶夜蛾,属鳞翅目、夜蛾科,是一种长距离迁飞、杂食性农业害虫[1-2]。 它起源于南亚,后广泛分布于北纬57°至南纬40°之间的地区,能危害170 多种作物[3-4],除甜菜和多种十字花科蔬菜外,还危害棉花、玉米、大豆、苜蓿等,对农业生产造成严重的经济损失[5-6]。20 世纪80 年代以前,该害虫仅在我国的局部地区零星发生,且以危害蔬菜作物为主。 但自20 世纪80 年代中期特别是进入90 年代以后, 随着设施农业的兴起和蔬菜大棚面积的不断扩大,棉花成为甜菜夜蛾的重要寄主之一[7-8]。
根据国际农业生物技术应用咨询服务中心(ISAAA)统计,2018 年中国转基因作物种植面积位居全球第七, 其中, 转基因棉花应用率达到95%,包括转基因抗虫棉。中国自20 世纪90 年代初开始研究抗虫基因以来, 通过遗传转化的方法,得到3 类使棉花获得抗虫功能的基因,其中来源于苏云金芽孢杆菌 (Bacillus thuringiensis,Bt)的杀虫蛋白基因应用最为广泛。 当前,大规模投入生产的国产转基因抗虫棉主要为单价抗虫棉和双价抗虫棉, 其遗传转化的抗虫基因为Bt杀虫基因cry1A和豇豆胰蛋白酶抑制剂基因Cp-TI[9]。 随着这些转基因棉的推广,其对甜菜夜蛾影响的研究报道不断增加。周秋霞[10]以抗Bt(cry1Ac)甜菜夜蛾品系为研究对象,初步阐明了该虫对温湿度等非生物因子和寄主植物等生物因子的适应性变化。 亦有学者系统研究了转cry1Ac/1Ab基因、 转cry1Ac+cry2Ab基因棉花对甜菜夜蛾存活、生长发育、营养指标及中肠酶活性的影响[11-12]。 虽然有研究报道转基因棉不同组织器官对甜菜夜蛾种群具有抗性[13],但关于不同转基因抗虫棉不同叶位叶片对甜菜夜蛾种群的影响研究鲜见。 因此,研究甜菜夜蛾在不同转基因棉花品种及叶位上的发生为害,分析并探讨其在棉花等作物上的发展态势, 对该害虫的预警和防范,以及转基因抗虫棉的持续推广均具有重要的现实意义。 鉴于甜菜夜蛾是长江中下游植棉区棉田生长前中期的重要食叶性害虫,且以取食棉花中上部叶片为主,本研究将通过研究转基因棉不同叶位叶片对甜菜夜蛾的影响,并以实验种群生命表的形式加以描述,来确定不同品种转基因棉对甜菜夜蛾的抗性, 为甜菜夜蛾的有效防治提供参考。
甜菜夜蛾卵块由中国农业科学院植物保护研究所提供,饲养甜菜夜蛾用的人工饲料参照李腾武等[14]的方法进行配制。
主要饲养条件:温度(27±1)℃;相对湿度70%±10%;光周期(光暗比)14 h∶10 h。
供试棉花品种: 转cry1A基因棉国抗12(GK12),亲本(常规对照,下同)为泗棉3 号;转cry1Ac基 因 棉 新 棉33B, 亲 本 为DP5415;转cry1A+CpTI基因棉SGK321,亲本为石远321。
6 个棉花品种均在4 月中旬播种育苗,5 月下旬移栽于扬州大学实验农牧场试验田内,每个品种种植200 m2。 供试棉叶采自棉花苗期至现蕾期的植株。

在室内将甜菜夜蛾刚孵化的幼虫接入培养皿中(直径15 cm),每皿25 头,分别用GK12、泗棉3 号、SGK321、石远321、新棉33B 和DP5415的倒三叶、倒六叶、倒九叶饲养,叶柄以脱脂棉保湿,并以一层保鲜膜加白纸为皿盖,以防幼虫逃逸。1 龄集中饲喂,2 龄后放入指形管(直径2 cm,高8 cm)单头饲喂,每天定时观察并记载幼虫的蜕皮和存活情况,适时更换新鲜叶片,清理排泄物。 幼虫化蛹当天用万分之一电子天平称量蛹质量,并作好标记,每天观察成虫的羽化情况。 将当天羽化的雌雄成虫配对后接入罐头瓶 (直径8 cm,高度12 cm),1 瓶1 对,瓶内放入蜡纸供其产卵,同时将蘸取体积分数10%蜂蜜水后的棉花球放入,以供成虫补充营养,瓶口扎上纱布;每天更换蜡纸并统计产卵量及成虫死亡情况,至成虫全部死亡止。 雌成虫产卵后,将蜡纸取出,剪下卵块。 在整个研究过程中,持续观察并统计幼虫死亡率、发育历期、化蛹率、蛹质量、蛹历期、羽化率、产卵量、成虫寿命等生物学参数[15-16]。 每个试验处理均重复3 次。

在产卵盛期, 挑选大小适中的初产卵块,接入放有6 种棉叶的培养皿(直径15 cm)中,每皿300 粒左右,重复3 次,孵化期间每天检查2 次,除去未受精卵,统计孵化率。 幼虫期用原处理中孵化的幼虫100 头,饲养幼虫方法与条件同上。
参照张小丽等[16]、吴坤君等[17]、韩兰芝等[18]的方法,并借助Morris-Watt 种群趋势指数模型I=S1·S2·S3……Sn·F·PF·P♀[19-20]编制甜菜夜蛾实验种群生命表。 式中,I为种群趋势指数,S1,S2,S3……Sn为生命表中各期致死因子作用后的平均存活率,P♀为雌性概率,F指标准产卵量,PF为达标卵量概率。

相关数据的平均数和标准误采用SPSS 25.0软件进行计算。 取食转基因棉品种不同叶位叶片对甜菜夜蛾生长发育及繁殖的影响与亲本间的差异显著性检验采用独立样本t检验[21]。 在进行甜菜夜蛾取食同一棉花品种不同叶位叶片对其生长发育及繁殖影响的方差分析时,使用单因素方差分析(ANOVA),并选择Tukey’s HSD 法进一步检验。


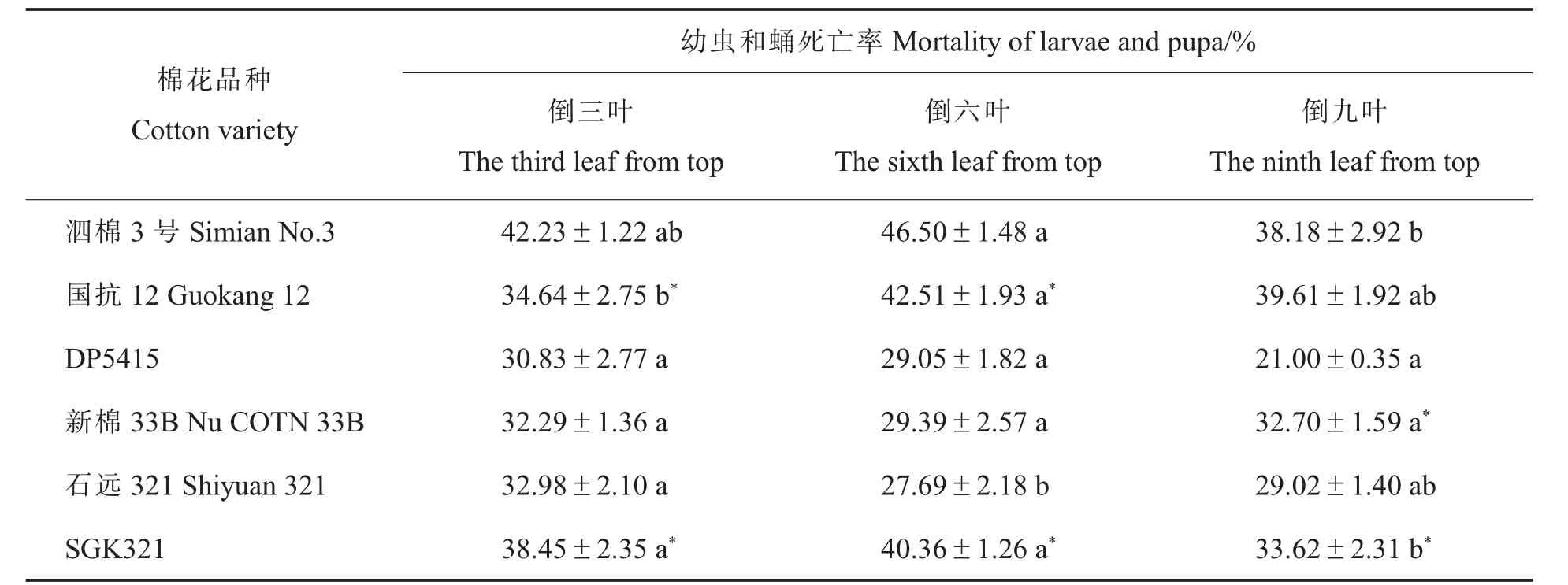
表1 取食转基因棉不同叶位叶片甜菜夜蛾幼虫和蛹的死亡率及其与亲本的差异比较Table 1 Comparison of the mortality of larvae and pupae of Spodoptera exigua fed on different leaves of transgenic cotton and their differences with their parents

进一步分析品种间的差异, 发现取食新棉33B 倒九叶的蛹质量显著高于其对照材料DP5415,取食新棉33B 倒六叶和取食SGK321 倒三叶的蛹质量与其相应对照材料(DP5415、石远321)差异不显著,而其他均表现为取食常规棉对照的蛹质量显著高于对应的转基因棉品种(表2)。
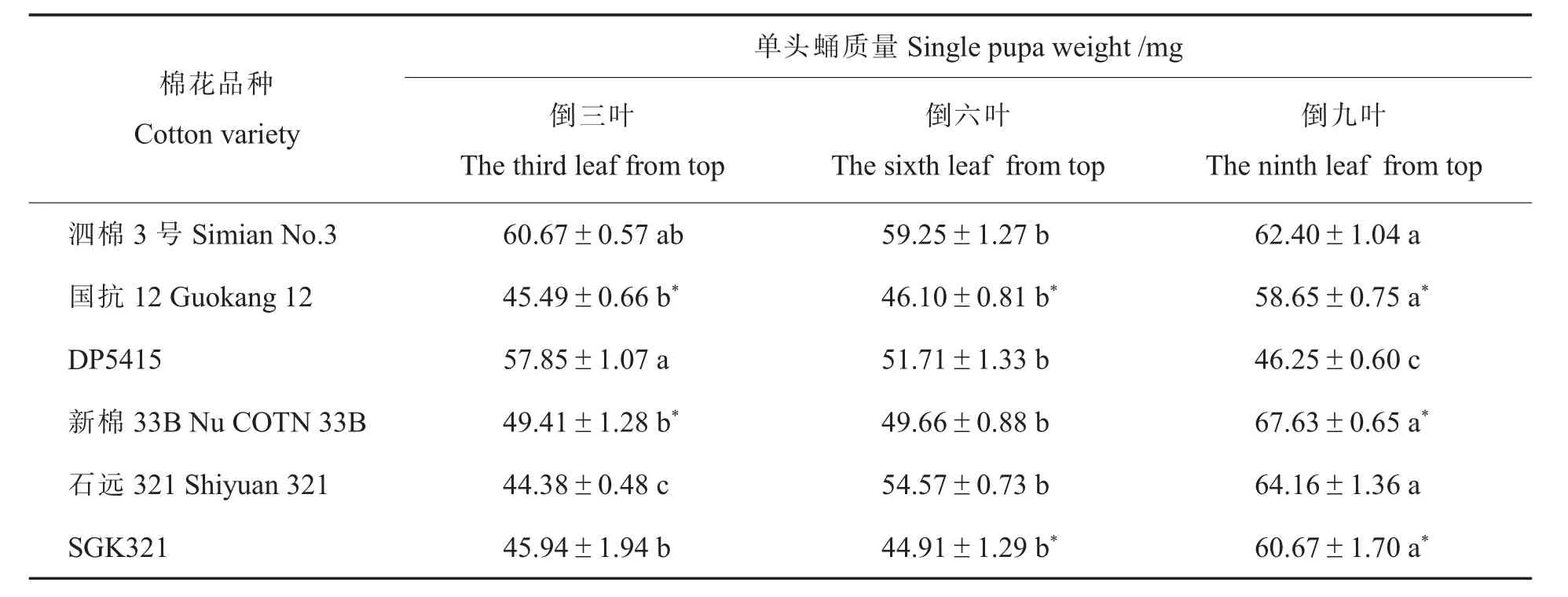
表2 取食转基因棉不同叶位叶片甜菜夜蛾的蛹质量及其与亲本的差异比较Table 2 Comparison of the pupae weight of Spodoptera exigua fed on different leaves of transgenic cotton and their differences with their parents

由表3 结果可见,在转基因棉与常规对照棉之间,仅取食转基因棉品种GK12 倒三叶、新棉33B 倒六叶和SGK321 倒三叶、倒九叶的甜菜夜蛾单雌产卵量与其亲本相比差异显著。 比较甜菜夜蛾取食同一品种不同叶位间的单雌产卵量后发现:只有取食新棉33B 倒九叶的甜菜夜蛾产卵量显著高于倒三叶和倒六叶,其他2 个转基因棉品种不同叶位间其产卵量均没有显著差异;取食亲本泗棉3 号倒三叶、DP5415 倒六叶和石远321倒九叶的单雌产卵量均高于同品种的其他叶位(表3)。

甜菜夜蛾实验种群生命表组建的结果(表4)表明,取食3 种转基因棉及其亲本不同叶位叶片的种群趋势指数(I)远大于1,说明取食不同棉花品种的甜菜夜蛾种群均处于增长态势。 分析发现,除取食GK12 倒三叶、倒九叶的甜菜夜蛾种群趋势指数略高于亲本泗棉3 号的相同叶位外,其他均表现为取食常规对照棉的甜菜夜蛾种群趋势指数高于对应的转基因棉品种,且一些取食转基因棉品种的甜菜夜蛾种群趋势指数只有对照的60%~70%。 另外,随叶位下移,取食GK12的甜菜夜蛾种群趋势指数总体呈现下降趋势,取食新棉33B 的甜菜夜蛾种群趋势指数呈先增后减的趋势,取食SGK321 的甜菜夜蛾种群趋势指数呈先减后增的趋势,但取食转基因棉花品种的甜菜夜蛾种群均呈增长状态(I>1)。
棉花是甜菜夜蛾的重要寄主之一[7-8],随着转基因抗虫棉的推广,分析并探讨其在不同转基因棉品种上的发展态势,对该害虫的预警和防范有重要的现实意义。
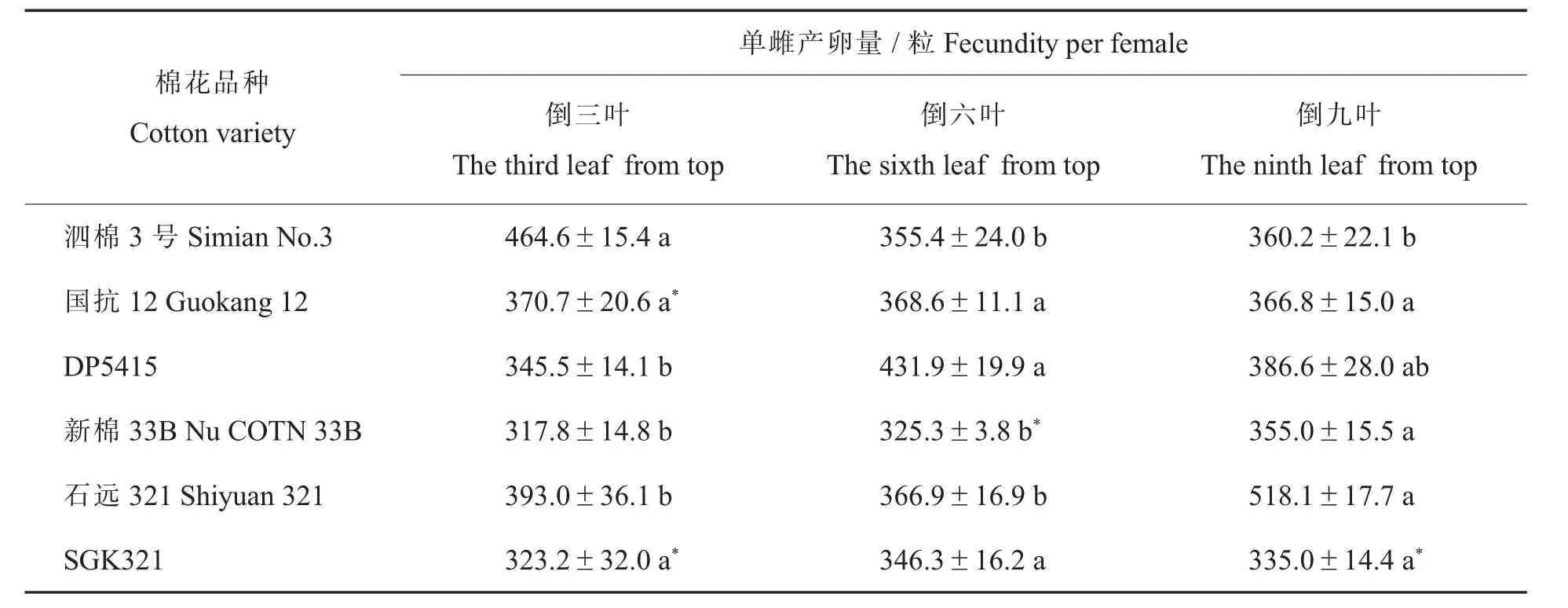
表3 取食转基因棉不同叶位叶片甜菜夜蛾的单雌产卵量及其与亲本的差异比较Table 3 Comparison of the fecundity per female of Spodoptera exigua fed on different leaves of transgenic cotton and their differences with their parents

比对后发现: 甜菜夜蛾取食GK12 倒三叶、倒九叶的幼虫和蛹累计死亡率与亲本间差异显著,这与长江中下游大面积推广种植的国产单价转基因抗虫棉遭遇甜菜夜蛾严重危害的情形相一致[9,22]。取食SGK321 不同叶位叶片的甜菜夜蛾累计死亡率均显著高于亲本,这表明复合性状的转基因抗虫棉对毒杀甜菜夜蛾有一定的效果[23]。在今后的研究工作中,一方面要系统测定棉花全生育期不同叶位叶片体内的毒蛋白表达量,另一方面要系统探讨甜菜夜蛾在棉花不同生育时期的种群增长动态。 考虑到毒蛋白对甜菜夜蛾幼虫的影响也许存在后效应,本研究中关于甜菜夜蛾的存活率没有将幼虫与蛹分开计算,这在以后的研究中需要进一步优化。
取食GK12 倒三叶、 新棉33B 倒六叶和SGK321 倒三叶、 倒九叶的甜菜夜蛾单雌产卵量与其亲本相比差异显著。 毒蛋白的影响可能在后续虫态中还有所表现。 但由于本研究没有关注存活甜菜夜蛾成虫的雌雄比和雌虫寿命等生物学参数,因此不能推断是因为死亡率上升而导致雌虫数量减少所引起的产卵量下降、还是由于毒性的后效应导致雌虫产卵量减少,这在以后的研究中值得深入研究。
甜菜夜蛾取食同一棉花品种不同叶位后的分析结果发现,幼虫至蛹阶段的死亡率在同一转基因棉品种不同叶位之间存在差异: 取食GK12倒三叶的死亡率显著低于倒六叶, 取食SGK321倒九叶的死亡率显著低于倒三叶、倒六叶。 蛹质量及产卵量的结果显示: 除取食亲本材料DP5415 和泗棉3 号外, 甜菜夜蛾取食其他棉花品种的叶位越低, 其蛹质量越高; 除取食GK12及其亲本泗棉3 号外,甜菜夜蛾取食其他品种棉花的叶位越低,整体上产卵量较高。 上述结果反映甜菜夜蛾取食的棉花叶位越低,越利于该种群的发展。 至于取食棉花叶位影响甜菜夜蛾蛹质量与产卵量的机理,还需要进一步阐明。

种群增长趋势指数是由各期存活率和成虫生殖力构成的[24]。 实验种群生命表的分析结果显
示,与常规棉对照相比,转基因棉GK12 不能抑制甜菜夜蛾种群的增长,新棉33B、SGK321 对甜菜夜蛾种群的增长有一定的抑制作用,取食个别叶位后甜菜夜蛾的种群趋势指数比对照常规棉品种降低了30%~40%,与这些品种的各虫态累积存活率下降、成虫达标卵量概率(PF值)下降有关 (比如倒三叶的新棉33B 与DP5415 之间、倒九叶的SGK321 与亲本石远321 之间)。 但本研究认为,虽然取食转基因棉花品种对甜菜夜蛾的种群有一定的抑制作用,但这种抑制不影响甜菜夜蛾的种群增长(I>>1)。

表4 取食不同叶位叶片的甜菜夜蛾实验种群生命表Table 4 The experimental population life table of S. exigua feeding on leaves of different leaf positions
进入21 世纪以来, 甜菜夜蛾已逐步成长江中下游植棉区棉花上的重要食叶性害虫,在棉花生长的前中期对棉花上中部叶片的为害越来越明显。 本研究中,甜菜夜蛾取食棉花现蕾期前后的上部叶片后,其种群趋势指数均大于1。表明棉花上中部叶片均是甜菜夜蛾的嗜食部位。 肖留斌等测定了8 种转基因抗虫棉叶片中Bt毒蛋白的含量,结果表明8 个转基因棉品种不同部位叶片的毒蛋白含量呈现顶叶最高、功能叶次之、老叶含量最低的趋势[25]。 也有研究者测试了转基因抗虫棉Bt毒蛋白含量的时空变化,指出转Bt基因抗虫棉根、茎和叶片等组织器官杀虫蛋白表达量在棉花不同生育时期有显著差异, 顺序为叶>铃、花蕊、花萼、蕾>花瓣>苞叶[26]。邢朝柱等采用酶链免疫法测定了转基因棉花铃期不同器官的Bt蛋白含量, 结果同样表明转基因棉体内的Bt蛋白含量表现为功能叶>幼铃>苞叶、老叶[27]。虽然有大量转基因棉Bt毒蛋白含量时空动态的研究报道,但各品种棉花不同生育时期、不同叶位叶片中毒蛋白含量变化的研究不多,这有待结合甜菜夜蛾种群的增长动态探讨。 同时,取食转基因棉花品种为什么会影响甜菜夜蛾蛹的存活率和成虫的达标卵量概率,其机理是什么,以及这种毒性的后效应对甜菜夜蛾种群衰减的贡献有多大,需要在今后深入阐释。
实验种群生命表的分析结果还表明,尽管取食转基因棉的甜菜夜蛾种群趋势指数总体低于相应的亲本,但种群趋势指数远大于1,表明该害虫种群在转基因棉花种植区仍将呈快速增长态势[24],需要密切关注。虽然在长江中下游棉区甜菜夜蛾种群的暴发与该害虫的迁飞扩散有关,也与一个地区棉花和大棚蔬菜面积的配比有关,但甜菜夜蛾在棉田的发生动向值得警觉。 有研究发现, 随着甜菜夜蛾在棉花上持续危害时间的延长,其种群增长的差异在常规棉与转基因棉花间有缩小的趋势[16]。

取食本研究所选取的3 类转基因棉花品种的不同叶位叶片后, 甜菜夜蛾幼虫和蛹死亡率、蛹质量及产卵量虽受一定的影响,但叶位间不存在明显的变化趋势。 虽取食转基因棉GK12 和SGK321 叶片对甜菜夜蛾种群增长有一定的抑制作用,但其种群趋势指数均大于1,表明其种群仍呈增长趋势;因此,生产上应进一步加强棉田甜菜夜蛾种群动态的监测,并逐步建立起防范甜菜夜蛾种群暴发的应急机制。
猜你喜欢
农业与技术(2023年11期)2023-06-19 02:28:18
干旱地区农业研究(2022年4期)2022-07-28 05:37:08
今日农业(2022年8期)2022-07-05 00:05:26
少儿科学周刊·儿童版(2022年10期)2022-06-27 14:08:45
四川蚕业(2022年1期)2022-06-06 02:03:42
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
中国糖料(2016年1期)2016-12-01 06:49:04
西南农业学报(2016年5期)2016-05-17 05:42:39
乡村科技(2014年23期)2014-03-08 06:37:27
