
New Hystrix (Hystricidae,Rodentia) from the Neogene of Linxia Basin,Gansu,China
2020-07-28 12:21:12WANGBanYueQIUZhanXiang
古脊椎动物学报(中英文) 2020年3期
WANG Ban-Yue QIU Zhan-Xiang
(Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044 wangbanyue@ivpp.ac.cn)
Abstract Four well preserved skulls of Hystrix recently collected from Linxia Basin were studied and a new species,H.brevirostra sp.nov,was established for them.The new species is characterized by its larger size,relatively low and wide skull with short rostrum and diastema,slightly posteriorly convex posterior border of relatively short nasal roughly aligning with lacrimal,shorter and deeply curved mandibular diastema,and relatively low-crowned cheek teeth.It is intermediate between H.lufengensis and H.gansuensis in evolutionary level,existing in Late Miocene‒Pliocene.While it might represent a lineage separated from the above two species.Ontogenetic variation in skull morphology and direction of cheek tooth rows of the new species was also discussed.
Key words Linxia Basin,Gansu;Late Miocene,Liushu Formation;Early Pliocene,Hewangjia Formation;Hystricidae
Hystrixis one of the Old World largest rodents ranging from Late Miocene to recent.Until recentlyHystrixfossils have been recorded mostly from European Neogene–Quaternay and Asian Quaternary,with only a few from Neogene of Asia (Lydekker,1884;Wang,1981;Shevyreva,1986;Pei,1987;Sen,1996,1999,2001a,b;Weers and Zhang,1999).In the last decades records of NeogeneHystrixfrom China have been slowly accumulated (Qiu et al.,1985;Wang and Qiu,2002;Weers,2004;Wang and Qi,2005).Just recently,some new and well preservedHystrixfossils,including 4 skulls,two of them with their mandibles,were collected from Neogene deposits in Linxia Basin.This is the best Neogene material ofHystrixso far known from Eurasia.Detailed study of these fossils shows that they not only represent a new species,but also enhance our understanding ofHystrixas a whole.
For terminology and method of measurements of the skull and mandible we follow Wang and Qiu (2018),and those of the cheek teeth follow Wang and Qiu (2002).
1 Systematic paleontology
Hystricidae Fischer de Waldheim,1817
Hysrix Linnaeus,1758
Hystrix brevirostra sp.nov.
(Figs.1‒4;Tables 1‒2)
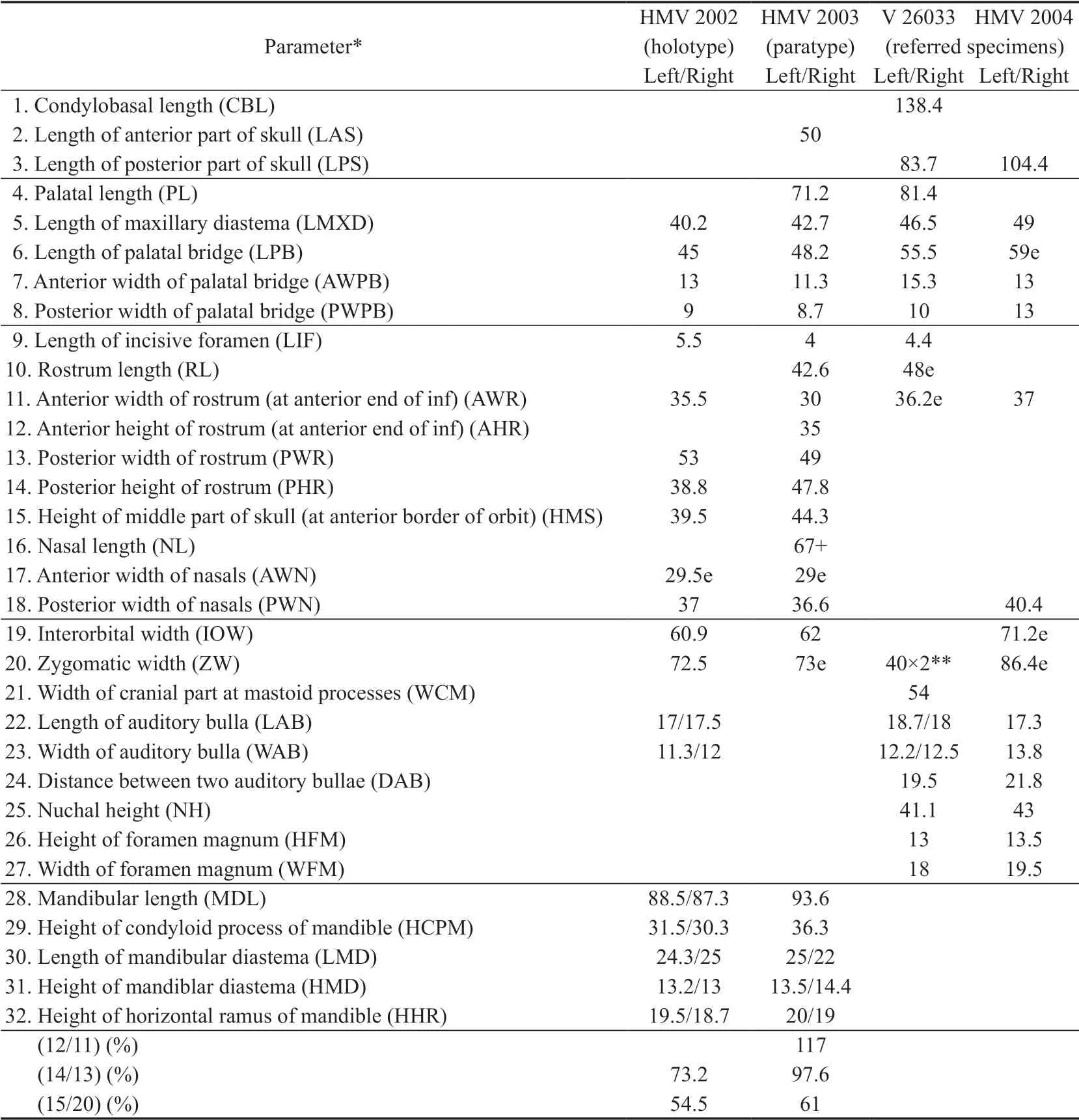
Table 1 Measurements of skulls and mandibles of Hystrix brevirostra sp.nov.from Linxia Basin (mm)

Table 2 Measurements of teeth of Hystrix brevirostra sp.nov.from Linxia Basin,Gansu (mm)
Hystrix gansuensis:Deng et al.,2011,p.455,fig.4a;Deng et al.,2013,p.260
HolotypeA nearly complete skull (but the basioccipital is slightly displaced from its original position) with mandible and a thoracic vertebra [HMV (Catalogue number of the Hezheng Paleozoological Museum of Gansu) 2002],from locality LX 200701 (= Loc.60 of Zhao Rong),Duikang,Zhuangkeji Village,Guanghe County;Early Pliocene Hewangjia Formation.
ParatypePartial skull (lacking occipital part) with mandible (HMV 2003),from the same locality and horizon as the holotype.
Referred specimensA nearly complete skull (IVPP V 26033) from LX 200205 (= Loc.54 of Zhao Rong),Baihuacun,Zhuangkeji Village,Guanghe County,upper part of the Liushu Formation,Late Miocene;and a complete skull (HMV 2004=X 2587),from LX 200041,Laohuwo of Shanchengcun,Maijiaji Village,Hezheng County,upper part of Liushu Formation,Late Miocene.
DiagnosisLarge-sizedHystrix.Skull low and wide,with relatively short rostrum and diastema;nasal relatively short,with posteriorly convex posterior border nearly aligned with the lacrimal.Horizontal ramus of mandible low,with a shorter and deeply concave diastema.Cheek teeth unilaterally subhypsodont,relatively low-crowned;sinus separated from both fold I and fold II in Dp4 and extending into mesoloph in P4.
Etymologybrevirostra:brevi,Latin,short;rostra,Latin,feminine,nose.It means that the animal has a relatively short rostrum.
2 Description
2.1 Skull
According to the age-classes of the skulls proposed by Weers (1990),theskulls of HMV 2002,HMV 2003 and IVPP V 26033 are young and belong to age-class IV,while HMV 2004 is adult and belongs to age-class VI.
The skulls are of the hystricomorphous type,relatively low and wide,and have a large infraorbital foramen (Figs.1‒3).
Dorsal viewThe skull is long ovoid in outline,with relatively narrow rostrum and occipital part.The nasals (N) are slightly longer than wide,with a slightly convex dorsal surface and a posteriorly convex posterior border,which is located much more posteriorly than the premaxillo-frontal suture and nearly aligned with the lacrimal.The premaxillo-frontal suture extends transversely.The maxillo-frontal suture extends obliquely,from the premaxillofrontal suture to the lacrimal.The lacrimal (L) is situated mainly within the orbit,with only a small part exposed on the dorsal side at the anteromesial corner of the orbit,where a distinct lacrimal tubercle (lt) forms.The orbit is large and the postorbital process (pp) is distinct.The frontals (F) are wide and have convex dorsal surface,but shorter than the nasals.The frontoparietal suture is M–shaped,roughly aligns with the posterior zygomatic root.Starting from the postorbital processes,the two weak parietal crests (pc) are convergent posteriorly to meet each other forming a short sagittal crest (sc),which reaches to the distinct nuchal crest (nc).The parietal (P) is slightly smaller than the frontal and has slightly concave surface.The interparietal (Ip) is bell-shaped,inserting between the two parietals.

Fig.1 Skulls and mandibles of Hystrix brevirostra sp.nov.from Linxia Basin,Gansu
Lateral viewThe skull is relatively low,with a slightly convex roof.The anterior end of the nasal is situated posterior to that of the premaxilla.Theincisura nasomaxilla(in) is small and shallow.The rostrum is high and short.On the lateral side of the rostrum there is a large concave surface,which is for the insertion of m.masseter profundus.The large infraorbital foramen (iof) is within the maxilla and its posterior border is above the M1 in position.The anterior border of the large orbit is situated above M2,and the postorbital process (pp) is above M3 or slightly posterior to M3.The lacrimal tubercle (lt) is located at the anterosuperior corner of the orbit.The bridge between the infraorbital foramen and the orbit is wide and thin-plate in form.The zygomatic arch is formed by zygomatic process of maxilla,jugal (J),and zygomatic process of the squamosal (Sq).The anterior zygomatic root formed from the maxilla is located near the anterior part of DP4 (P4).The anterior part of zygomatic arch (= the inferior margin of infraorbital foramen) is a slim triangular plate,and its posterior part (= inferior margin of orbit) forms a transversely thin and vertically wide plate.The small premolar foramen (prf) is located on the mesial wall of the infraorbital foramen,above the anterior border of Dp4 (P4).The sphenopalatine foramen (spf) is large and located above M1‒2.The optic foramen (opf) is relatively small and located above the posterior border of M3 or slightly posterior to M3 and superoposteriorly to the sphenopalatine foramen.The large anterior alar fissure (aafi) is located inferolaterally to the optic foramen.The tiny ethmoidal foramen (etf) is located near the superior border of the orbit and posteriorly to the lacrimal tubercle (lt).
Ventral viewThe diastema between I2 and DP4 (P4) is short compared with the upper cheek tooth row (ratios of length of diastema to that of P4‒M3 are 1.16 and 1.18 in HMV 2004).The incisive foramen (inf) is small and located at the anterior 1/3 of the diastema.A pair of tiny interpremaxillal foramina (ipmf) are located between I2 and inf.The premaxillomaxillal suture extends anteromesially and passes the posterior end of incisive foramen.On the anterior part of the maxilla there is a pair of bulges showing the positions of the posterior ends of the alveoli of the two I2s.Between the P4 (dp4) and the premaxillo-maxillal suture there is a large concave area,which may be the site for the insertion of m.buccinator.In front of P4 (dP4) and near the mid-maxillal suture there is a pair of small posterior palatine foramina (ppf).The left and right cheek tooth rows are slightly convergent posteriorly (in young) or parallel to each other (in adult).On the hard palate between the two teeth rows there are several nutrient foramina.The transversely extending mesial part of the maxillo-palatine suture aligns with the boundary between M1 and M2 (in HMV 2002 and HMV 2003) or with anterior part of M2 (in HMV 2004 and V 26033).The posterior border of the hard palate (= anterior border of the mesopterygoid fossa) is situated mesial to the M2 (in HMV 2002,HMV 2003 and V 26033),or to the anterior part of M3 in adult (HMV 2004),and usually has a small protrusion at the middle.The mesopterygoid fossa (mptf) is large and wide.The pterygoid fossa (ptf) is very narrow and small,with an internal pterygoid process (ipp) higher than the external pterygoid process (epp).The posterior foramen of alisphenoid canal (ascp) penetrates the posterior part of the external pterygoid process.The masticatory foramen (mf),buccinator foramen (bf) and deep temporal foramen (dtf) are confluent into one foramen.The posterior foramina (mfp,bfp and dtfp) of the three nerve canals mentioned above are also confluent into one,which is located dorsoposterior to the ascp and anteroventral to theforamen ovale.Theforamen ovale(fo) is confluent with the middle lacerate foramen (mlf).
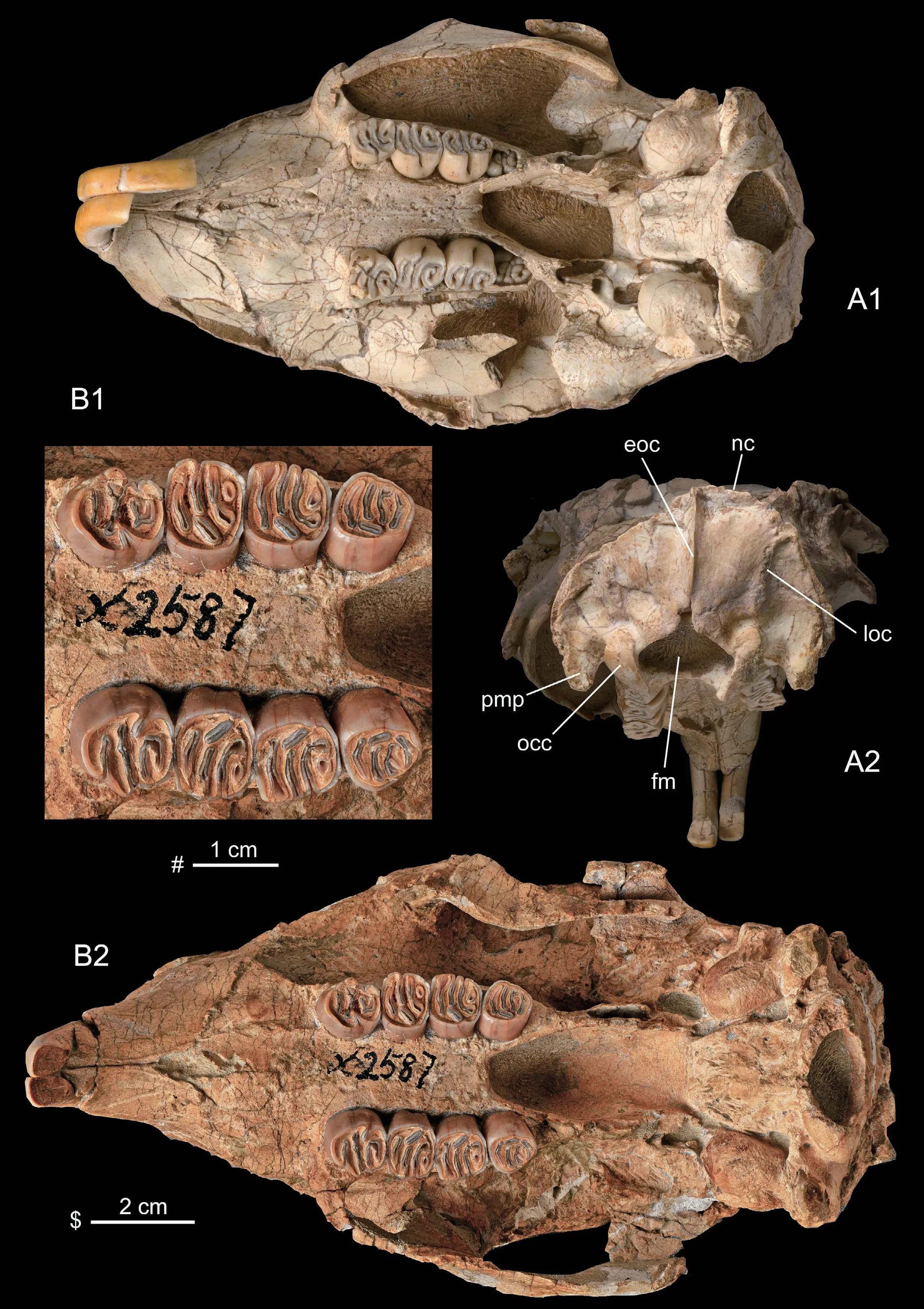
Fig.3 Skulls of Hystrix brevirostra sp.nov.from Linxia Basin,Gansu
The two zygomatic arches are slightly divergent posteriorly.The glenoid fossa (glf) of the squamosal takes a form of short and wide groove which is longitudinally straight but transversely concave.The small auditory bulla (ab) has a short external auditory meatus (eam).The bulla has a spherical surface.The stylomastoid foramen (styf) is located between the bulla and the mastoid process (msp).The jugular foramen (juf) is located mesioposterior to the bulla.The hypoglossal foramen (hyf) is located lateral to the basioccipital and anterior to the occipital condyle (occ).
Posterior viewThe nuchal surface is about semicircular in outline,rather flat and nearly vertical to the ventral surface of the skull.The external occipital crest (eoc) is developed and extending from nuchal crest (nc) to the superior border offoramen magnum(fm).On the two lateral areas of eoc there is a pair of vertical crests,which may be the lateral borders of the areafor insertion of m.rectus capitis dorsalis minor,called here as lateral occipital crests (loc).The paramastoid process (pmp) is developed.Theforamen magnumis ovoid in outline.The two occipital condyles (occ) are separated widely.
2.2 Mandible (Fig.4)
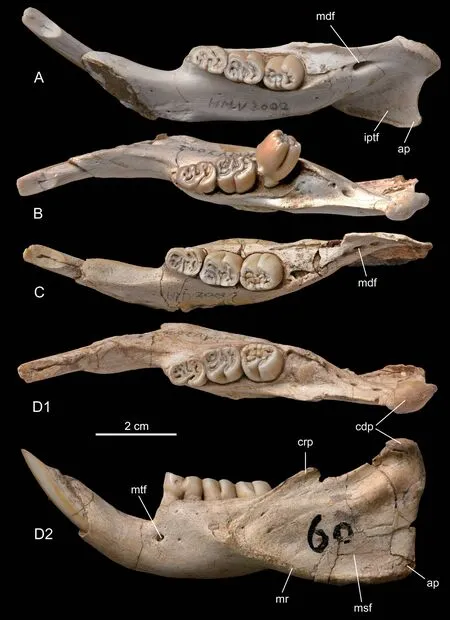
Fig.4 Mandibles of Hystrix brevirostra sp.nov.from Linxia Basin,Gansu
Both HMV 2002 and HMV 2003 preserve left and right hemimandibles.The mandible is of hystricognathous type,with the angular process extending laterally to the horizontal ramus.The horizontal ramus is straight and low.The symphysis extends posteriorly to below the anterior root of dp4.The mandibular diastema is shorter than dp4‒m2 in length and is deeply concave with a steep posterior part.The mental foramen (mtf) is located below the anterior root of dp4 (p4) in the middle height of buccal side of the horizontal ramus.The masseteric fossa (msf) is large.The distinct masseteric ridge (mr) extends anteriorly to below the m1,nearly at the same horizontal level of the mental foramen.The ascending ramus of mandible is short and low,and extends slightly laterally to the cheek tooth row.The coronoid process (crp) is very low and small,with its anterior border rising from lateral side of horizontal ramus below m1 and its top being slightly higher than the occlusal surface of the lower cheek tooth row.The condyloid process (cdp) is higher and larger than the coronoid process,and has an ovoidhemispherical articular facet.The angular process (ap) is small and bends slightly lingually below the condyloid process.On the lingual side of the ascending ramus the mandibular foramen (mdf) is situated posterior to the m3.The mdf may be represented by a large single foramen (in HMV 2002) or may be separated into three small ones (in HMV 2003).The internal pterygoid fovea (iptf) is large and deeply concave,with its anterior end extending to below m2 and its lower margin rolling up lingually.
2.3 Teeth
The dental formula is 1·0·1·3/1·0·1·3.The cheek teeth are unilaterally subhypsodont,with the lingual side higher (lower) than the buccal side on the upper (lower) cheek teeth respectively.
The I2 is orthodont,with its anterior end bending ventrally or slightly backwards.In V 26033 the two I2s have longer and more curved anterior parts,which may represent an abnormal phenomenon.The cross section of the I2 is ovoid,with a wider and slightly convex labial surface.The enamel covers the labial side,about 1/3 medial and 1/2 lateral sides.The surface of labial side is smooth and no longitudinal ridge can be seen on it.
P4 and dP4:HMV 2004 is an adult individual and preserves P4‒M3.The P4 is the largest of the upper cheek teeth.It is ovoid in outline,slightly longer than wide,with a convex anterior border.The buccal end of fold I is shallowly open (on right P4) or closed with wear (on left P4).Fold II on the right P4 is buccally open,extending upwards almost to the base of the crown on the buccal wall.Fold II on the left P4 is Y-shaped,embracing the enlarged mesostyle.Fold III and IV join together to form a U-shaped fold,which is closed buccally.The protocone joins the anteroloph,protoloph and mesoloph.The anteroloph is shorter than the other lophs.The sinus is short transversely and extends into the mesoloph and does not meet the fold II.Its lingual opening extends upwards almost to the upper 3/5 of the crown height on the lingual side.
HMV 2002,HMV 2003 and V 26033 are all young,and have erupted dP4‒M2 and M3 still in alveolus.The dP4 is smaller than M1 in size and trapezoid in outline,slightly longer than wide,with buccal side longer than lingual one.In HMV 2002 and V 26033 the fold I‒III remain shallowly open buccally,but in HMV 2003 the buccal end of the anteroloph begins to join the paracone to close fold I on buccal side.In all the dP4,mesoloph,fold III and metaloph are L-shaped.Fold IV forms a small closed basin.The paracone and mesostyle are usually distinct cusps in HMV 2002 and V 26033,and in HMV 2003 the mesostyle forks buccally.The sinus extends anterobuccally,but does not join with fold I or fold II.
M1 is rectangle in outline,slightly longer than wide in young,but wider than long in adult.In HMV 2002 fold I is open bucally and the paracone extends anteriorly.Thus,the buccal opening of fold I is shallower and easily to be worn out.In the other specimens,fold I is closed buccally.The buccal ends of fold II and III are open shallowly or closed with wear.As in dP4 the mesoloph of M1 is L-shaped and its lingual part extends posteriorly to join the posteroloph.The metaconule is variable:it may be a distinct cusp and the metaloph is only a transverse loph (as in HMV 2002);or it may extend posteriorly to meet the posteroloph forming an L-shaped metaloph (in other specimens).Thus,fold IV may join with fold III to form a U-shaped fold (as in HMV 2002) or may be a closed basin (as in other specimens).In any case,the buccal end of fold IV is closed.The sinus extends anterobuccally to join with fold II in the young,but is separated from the latter in adult.Its lingual end is open and extends upwards to about 1/3 of the crown on the lingual side in young,but closed in adult.
M2 is trapezoid in outline,longer than wide,with a slightly narrower posterior side than anterior side in young,but is rectangular in adult.The other features of M2 are similar to M1.
M3 of HMV 2004 is trapezoid in outline,with shorter and convex lingual and posterior sides.Fold I‒IV and the sinus are all closed.The sinus extends nearly longitudinally towards the fold I.Fold III and IV are transversely long folds.M3 of V 26033 is ovoid in outline,longer than wide,with a narrower and convex posterior border.It is unworn,with the occlusal feature similar to that of M2 of HMV 2002.
The i2 is slightly curved and its anterior part extends anterosuperiorly,and the posterior end extends posteriorly to the mandibular foramen.The cross section of the i2 is narrowly ovoid,with a slightly convex labial surface.As in the I2,the enamel covers the labial side,1/3 medial and 1/2 lateral sides.The surface of labial side is smooth and no longitudinal ridge is seen on it.
The dp4 is ovoid in outline,longer than wide and with a narrow and convex anterior border.Fold I is a small closed basin.The metalophid curves anteriorly,with its middle part meeting the mesolophid to separate fold II into two parts:the lingual one may open lingually (in HMV 2002) or may be closed (in HMV 2003);the buccal one is a closed fold.The mesolophid is variable:it may be a single lophid,or may be separated by small folds into several ones.Fold III is open lingually.Fold IV is closed lingually.There are some protrusions in fold III and IV.The sinusid is open buccally and its lingual end may or may not join with fold IV.The dp4 has three roots:a large anterior one and two small posterior ones.
The m1 is rectangular in outline,longer than wide.The metalophid is a very small circle and closes fold I into a very tiny basin.The mesolophid curves anteriorly to meet the anterolophid.Thus,fold II is L-shaped or U-shaped.Its lingual end is open.The hypolophid is long and may or may not join the posterior arm of the protoconid.Fold III is long and curved,and sometimes there are some small protrusions in it.Fold IV may or may not join with the sinusid,and there are also some protrusions in it.The lingual openings of fold II‒IV are very shallow and easily to be closed with wear.The buccal opening of the sinusid is deep and extends downwards nearly to the 2/3 crown height on the buccal side.
The m2 is similar to m1 in outline and structure.But it is scarcely worn and no dentine is seen on the occlusal surface.The m2 has four roots:two small anterior and two large posterior ones.
2.4 Vertebra
The thoracic vertebra of HMV 2002 preserves spinous process,vertebral body,pedicle,transverse process,pre- and postzygapophyses.The long spinous process extends vertically,with ridged anterior and posterior borders.The vertebral body is short.The caput and vertebral fossa are triangle-shaped,but their epiphyses are lost.On either of the left and right lateral borders of the caput and vertebral fossa there is an articular facet for the head of the rib.The prezygapophysis extends forwards from the pedicle.The articular facet on the prezygapophysis is oval in outline,slightly concave,facing superomesially.The postzygapophysis is located on the posterior part of the pedicle.The articular facet on the postzygapophysis is oval,slightly concave,and facing inferomesially.The robust transverse process extends laterally from the pedicle,situated lower than the pre- and postzygapophyses in position.Its distal end is damaged.
3 Comparison
The above described skulls are similar to the genusHystrixrather than toAtherurusandTrichysin being large in size;skull having convex roof,enlarged nasal cavity;long and wide nasal with a convex dorsal surface and posteriorly convex posterior border;frontal being shorter than nasal,but larger than parietal in size;having large infraorbital foramen and small and round auditory bulla,etc.
In comparison with the known Pleistocene and living species ofHystrix,includingH.brachyura,H.subcristata,H.hodgsoni,H.cristata,H.indica,H.javanica,H.kiangsenensis,H.largrelii,H.zhengiandH.magnaetc.(Lönnberg,1924;Wang,1981;Pei,1987;Weers,1990;Huang et al.,1995;Guo,1997;Weers and Zheng,1998;Sen 1999;Weers and Zhang,1999;Tong,2005;Weers,2005;Pan et al.,2007),the specimens described above are different from them in cheek teeth being lower crowned.Besides,the specimens are larger thanH.brachyura,H.subcristata,H.hodgsoni,H.cristata,H.indica,H.javanica,H.kiangsenensisandH.largreliiin size.In addition,they differ fromH.lagrelii,H.brachyura,H.indicaandH.javanicain having enlarged nasal;fromH.hodgsoni,H.subcristataandH.magna1)Hystrix magna was erected by Pei (1987).When he described the new species H.magna,Pei (1987) did not mention the holotype of the new species.Later,Weers and Zheng (1998) chose the left M1/2 (IVPP V 5036.12) from the specimens described by Pei (1987) as the lectotype of H.magna.However,having not mention the paper of Weers and Zheng (1998),Wang (2019:336) once again chose a lectotype for H.magna,the right P4 (IVPP V 11550.2) described by Weers and Zhang (1999).It seems to us that the lectotype of H.magna should be the left M1/2 (V 5036.12) rather than the right P4 (V 11550.2).in skull being lower and wider,with less vaulted dorsal surface,smaller nasal cavity and shorter nasal.
NeogeneHystrixis known including 11 species:H.parvae,H.primigenia,H.sivalensis,H.leakeyi,H.refossa,H.depereti,H.aryaensis,H.trofimovi,H.caucasica,H.gansuensisandH.lufengensis(Shevyreva,1986;Weers,1994;Weers and Montoya,1996;Sen,1996,1999,2001a,b;Wang and Qiu,2002;Weers and Rook,2003;Lopatin et al.,2003;Wang and Qi,2005;Sen and Purabrishemi,2010).
In comparison with these Neogene species ofHystrixthe specimens described above are larger thanH.parvae,H.leakeyiandH.aryaensis;the crowns of their cheek teeth are higher than those ofH.parvae,H.primigenia,H.sivalensis,H.trofimoviandH.lufengensis,but lower than those ofH.leakeyi,H.refossa,H.depereti,H.aryaensis,H.caucasicaandH.gansuensis.In addition,they differ fromH.primigenia,H.gansuensis,H.deperetiandH.lufengensisin having shorter rostrum;fromH.gansuensisandH.deperetiin having shorter nasal;fromH.primigenia,H.refossa,H.deperetiandH.lufengensisin having lower horizontal ramus of mandible,and shorter and deeper concave mandibular diastema.Besides,they differ from all of the known species ofHystrixin P4 having sinus extending into mesoloph,and fromH.gasuensisin sinus of dP4 being separated from fold I.
The above comparison tends to show that the new specimens described above represent a new species ofHystrix,named asH.brevirostrahere.
4 Discussion
Deng et al.(2011:455,fig.4a) referred a skull (without mentioning the catalogue number) from Duikang (LX 200701) toHystrix gansuensis,a species established by Wang and Qiu in 2002.According to their figure 4a,this specimen should be the HMV 2003,the paratype ofH.brevirostradescribed in this paper.As noted above,this skull (HMV 2003) is different fromH.gansuensisin having shorter rostrum and nasal,cheek teeth being lower crowned and differing in occlusal features.The skull (HMV 2003) should be referred to the new species,H.brevirostrarather than toH.gansuensisas Deng et al.(2011,2013) did.
There are considerable ontogenetic variations inHystrix brevirostra:among the four skulls three (HMV 2002,2003 and V 26033) are young and one (HMV 2004) is an adult individual.Although all the three young skulls are lumped to IV stage based on Weers (1990),their individual ages are not the same:in HMV 2002 and HMV 2003 the M3 are still in their alveoli,while in V 26033 M3 begins to erupt.Thus,V 26033 is slightly elder than the other two in age.In observing and describing the skulls we found the following variations in the four skulls:
(1) The posterior border of the hard palatine is variable in position:it alignes with the sinus of M2 in HMV 2002 and HMV 2003,with the posterior part of M2 in V 26033,and with the anterior part of M3 in HMV 2004.It would mean that the posterior border of the hard palatine ofH.brevirostramoves posteriorly with age in young individuals.
(2) The posterior border of nasal is above M2 (in young),but moves to above M3 (in adult) in the position,again moving posteriorly with age in young individuals.
(3) The two upper tooth rows are slightly convergent posteriorly in young individuals,but parallel to each other in adult.
Wang and Qiu (2002) discussed the evolutionary tendencies among the species ofHystrix.Most of these evolutionary tendencies are substantiated by the new species.They are:skull changed from relatively lower and wider to higher and narrower in proportion;the rostrum became higher;the nasals and frontals enlarged;the crown of the cheek teeth became more hypsodont;and M3/m3 became reduced.Based on these evolutionary tendencies,it is obvious thatH.brevirostrais more primitive than all of the Pleistocene and living species ofHystrix.
Up to now only three species ofHystrixare known from Neogene of China.They areH.gansuensis,H.lufengensisandH.brevirostra.As stated above,H.brevirostradiffers fromH.gansuensisin skull having shorter rostrum and nasal,and cheek teeth being lower crowned.On the contrary,the cheek teeth ofH.brevirostraare higher crowned than those ofH.lufengensis.Thus,H.brevirostrais slightly more progressive thanH.lufengensis,but more primitive thanH.gansuensis.However,H.breviostrahas a shorter rostrum than the other two species.Therefore,H.breviostramay represent a different evolutionary lineage as the other two species.
Compared with other Neogene species ofHystrixoutside China,based on the crown height of the cheek teeth,it seems thatH.brevirostrais more advanced thanH.parvae,H.primigenia,H.sivalensisandH.trofimovi,but more primitive thanH.leakeyi,H.refossa,H.depereti,H.aryaensisandH.caucasica.However,this should be further testified by more convincing material to be discovered in future.
The specimens ofH.brevirostraare collected fromthree different localities:HMV 2002 and HMV 2003 from Duikang (LX 200701),IVPP V 26033 from Baihuacun (LX 200205),and HMV 2004 from Shanchengcun (LX 200041).The geologic age of Duikang is known to be Lower Pliocene (Hewangjia Formation,Deng et al.,2011).The other fossils from Baihuacun (LX 200205) are known to include four species:Gazellacf.G.gaudryi,G.paotehensis,HipparioncoelophyesandH.hippidiodus(Deng et al.,2013;Li,2014).Among them the first three are known from Late Miocene Yangjiashan fauna.The last one,H.hippidiodus,has been known from Late Miocene deposits of Qingyang.The fauna from Baihuacun may be correlated with the Late Miocene Yangjiashan fauna.
The other fossils from Shanchengcun (LX 200041) are known to include 6 species:Hipparion weihoensis,Acerorhinus hezhengensis,Chilotherium wimani,Chleuastrochoerus stehlini,Cervavitus novorassiaeandMuntiacussp.According to Deng et al.,2013,H.weihoensishas been known to be early‒middle Bahean ranging from Guoligou Fauna to Dashengou Fauna in age;A.hezhengensis,Chi.wimaniandChl.stehliniare known to be middle‒late Bahean ranging from Dashengou Fauna to Yangjiashan Fauna in age;C.novorassiaeis known to range from late Bahean Yangjiashan Fauna to Early Pliocene in age.Muntiacusis known to range from Late Miocene to Recent.It seems that the fauna from Shanchengcun can also be correlated with the Late Miocene Yangjiashan Fauna.
To sum up,Hystrix brevirostaranges from Late Miocene late Bahean to Early Pliocene Gaozhuangian in age.
AcknowledgmentsThe authors would like to express their gratitude to Director He-Wen and Vice director Cheng Shan-Qin of the Hezheng Paleozoological Museum of Gansu (HPM) for kindly permitting the authors to study the specimens of HPM.The authors are also greatly indebted to the following colleagues of IVPP:to Profs.Zheng Shao-Hua,Ni Xi-Jun and Zhang Zhao-Qun for their valuable discussion;to Profs.Ni Xi-Jun,Deng Tao,Wang Shi-Qi,Wang Yuan and Tong Hao-Wen for their multifarious help,and to Profs.Qiu Zhu-Ding and Zhang Zhao-Qun for their careful review of the manuscript and their valuable suggestions to improve the text of the paper.Many thanks are also given to Mr.Wang Ping for his careful and skillful preparation of the studied fossils,to Mr.Gao Wei for the photographs,and to Mrs.Si Hong-Wei for her help in preparing the pictures.
