
运动通过调节纹状体A2AR/D2DR表达改善帕金森病模型大鼠行为功能障碍
2020-07-22 01:31刘晓莉赵刚王弘王宗兵张丹昱
中国运动医学杂志 2020年5期
刘晓莉 赵刚 王弘 王宗兵 张丹昱
北京师范大学体育与运动学院(北京100875)
帕金森病(Parkinson’s disease,PD)是一种以进行性运动功能障碍为主要临床特征的中枢神经系统退行性疾病[1]。黑质致密部多巴胺能神经元变性缺失、纹状体多巴胺(dopamine,DA)水平显著减少,诱发基底神经节直接与间接通路功能紊乱是导致PD 运动障碍发生的主要原因[2]。体力活动或身体运动能够改善PD 患者的自主活动能力,提高DA 药物的疗效,延缓PD 的病理进程[3],其神经生物学机制备受关注。近年来,本实验室利用PD 动物模型证实,跑台运动通过调节基底神经节功能失衡改善了PD 大鼠的行为功能障碍[18],但其作用的分子靶点尚不明确。腺苷(adenosine,A)作为一种神经调质参与多种神经递质的释放或突触后神经元电活动的调节。腺苷2A 型受体(adenosine A2Arecep⁃tor,A2AR)和多巴胺Ⅱ型受体(dopamine D2receptor,D2DR)共存于纹状体中等多棘神经元(medium spiny neurons,MSNs)突触后膜,通过变构与配体结合或与其它受体相互作用形成多聚体聚合物,调节突触后神经元的功能。A2AR 拮抗D2DR 表达是导致间接通路过度激活的重要原因,这与PD 状态下运动障碍发生密切相关[4]。运动可能通过抑制A2AR 或激活D2DR 纠正PD 状态下纹状体-苍白球通路γ-氨基丁酸(γ-amino butyr⁃ic acid,GABA)神经元的过度激活,抑制间接通路过度兴奋,缓解基底神经节直接与间接通路的失衡,从而达到改善病人行为功能障碍的治疗效果[5],但目前尚缺失直接的实验证据。为此,本研究从A2AR、D2DR 入手,进一步揭示跑台运动干预改善PD 大鼠行为障碍的神经生物学机制,为运动康复在PD 临床治疗中的推广与应用提供实验依据。
1 材料与方法
1.1 实验对象与分组
实验选取雄性清洁级SD 大鼠,6 周龄,体重230~250 g,由北京华阜康生物科技股份有限公司提供[生产许可号:SCXK(京)2009-0007]。所有大鼠分笼饲养,自由饮食,自然光照,动物房室内温度控制在20℃~25℃,相对湿度为 45%~50%。大鼠进入实验室后适应性饲养7 天,期间进行为期3 天的跑台适应性训练,剔除无法完成预设跑台训练的大鼠。被选取的所有实验大鼠随机分为:假手术组(Control组)、假手术运动组(Control+Ex 组)、帕金森模型组(PD 组)和帕金森模型运动组(PD+ Ex 组);模型组(PD 组和PD+ Ex 组)造模术后根据第7天旋转行为结果剔除不符合PD模型标准的大鼠,各组大鼠样本量为:Control 组22 只,Control+Ex 组 22 只,PD 组 38 只,PD+ Ex 组 38 只。4 组大鼠分别进行酪氨酸羟化酶(tyrosine hydroxylase,TH)染色(n=6)、A2AR 及D2DR mRNA 表达(n=8)和蛋白表达检测(n=8)。PD 组和 PD+Ex 组大鼠还分别进行 D2DR 激动剂QUIN 和A2AR 拮抗剂SCH+QUIN 联合药物干预实验(QUIN 组、SCH+QUIN 组、Ex+QUIN 组、Ex+SCH+QUIN,每组均n=8)。
1.2 PPDD大鼠模型制备与评价
1.2.1 PPDD大鼠模型的制备
采用向右侧内侧前脑束(medial forebrain bun⁃dle,MFB)单侧注射 6 羟基多巴胺(6-Hydroxydopa⁃mine,6-OHDA)的方法制备PD模型。大鼠造模前禁食24 h,10%水合氯醛(0.35 mL/kg)腹腔注射麻醉。剪去头顶毛发,皮肤消毒后沿正中线切开约1 cm 开口,剥离骨膜,充分暴露前、后囟。PD 和PD+Ex 组大鼠俯卧位固定于脑立体定位仪,参照Paxinos 与Watson 大鼠脑立体定位图谱[6],定位右侧MFB(AP:-4.3 mm;R:1.5 mm;V:7.6 mm),颅骨钻垂直钻透颅骨,微量注射器吸取4 μL 6-OHDA 溶液,缓慢进针后以1 μL/min 速度注射,注射完后留针5 min,以1 mm/min 速度缓慢退针,缝合伤口并涂抹青霉素防止感染。Control 组与Control+Ex 组大鼠采用同样方法注射同等剂量含0.02%抗坏血酸的生理盐水。术后注意禁食与保暖,待动物清醒后放回笼中饲养。
1.2.2 PPDD大鼠模型的评价
阿扑吗啡(apomorphine,APO)诱导旋转行为实验:在造模手术后第7、14、28 天,按照 0.5 mg/kg 剂量于颈背部皮下注射非特异性DA受体激动剂APO溶液,诱发大鼠出现旋转行为,计数旋转圈数。根据第7 天旋转行为结果,以向健侧旋转圈数与损毁侧旋转圈数差值>100 r/30 min作为判定PD造模成功的标准[6],剔除不符合标准的模型组大鼠。
酪氨酸羟化酶免疫组织化学检测:全部实验结束后,大鼠腹腔注射10%水合氯醛麻醉,经左心室-升主动脉插管灌流(30 mL生理盐水和30 mL 4%多聚甲醛溶液),灌流结束后迅速取出脑组织置于4%多聚甲醛溶液中固定24 h。将脑组织取出后置于30%蔗糖溶液中脱水后修块、包埋、修整,并连续冠状切片(片厚10 μm),每只6张。免疫组织化学染色、脱水、透明、封片,所用一抗:兔多克隆抗体(1∶1000,Abcam 公司,美国),二抗:过氧化物标记的抗兔IgG(H + L)抗体(1∶200,KPL 公司,美国);利用荧光显微镜(DP72,奥林巴斯公司,美国)对黑质致密部区域拍照,采用Image-Pro Plus 6.0 软件对黑质酪氨酸羟化酶(tyrosine hydroxy⁃lase,TH)免疫阳性纤维光密度(optical density,OD)进行统计分析。
1.3 跑台训练方案
造模手术后24 h,参照Tajiri 等[7]运动方案(11 m/min ,30 min/day,5 day/week)对 Control+Ex 组和 PD+Ex 组大鼠进行4 周跑台运动干预,并根据第7 天APO诱导旋转行为测试结果,剔除不符合PD 模型标准的PD+Ex 组大鼠。训练期间将Control 组与PD 组大鼠置于跑台旁侧,排除设备和环境对大鼠的干扰。
1.4 主要实验仪器
MF-5362 型颅骨钻(Silite Corporation,加拿大),68002 型标准型双臂脑立体定位仪(瑞沃德,深圳),微量注射器(Hamilton,瑞士),DSPT-202 型动物跑台(段氏,杭州),Detect 凝胶成像系统(百泰克生物,北京),CFX96 Real-Time PCR 仪(Bio-Rad,美国),Rota-Count-8 旋转计数器(Columbus,美国),冰冻切片机(Thermo ,Microm International GmbH ,德国),奥林巴斯荧光显微镜(BX53,Olympus,日本)。
1.5 主要实验试剂与配制
主要实验试剂:6 羟基多巴(6-OHDA,Sigma,美国),L-抗坏血酸(Sigma,美国)阿朴吗啡(APO ,Sig⁃ma,美国),冷冻切片包埋剂(SAKURA,日本),Triton X-100 :(中杉金桥,北京),鼠抗A2AR单克隆抗体(San⁃ta Cruz Biotechnology,美国),兔抗D2DR、TH 多克隆抗体(Abcam,香港),荧光素化山羊抗鼠/抗兔二次抗体(Molecular Probes,美国),SCH58261(Abcam,香港),Quinpirole(QUIN,Sigma,美国),总RNA 快速提取试剂盒、逆转录试剂盒、Real-time PCR 试剂盒(百泰克生物,北京)。
主要试剂配制方法:6-OHDA 试剂配制:2 mg 6-OHDA 与0.2 mg 抗坏血酸溶解于1 mL 生理盐水(0.9 %)中,EP 管分装,锡纸包裹-80℃保存;APO 诱导旋转试剂配制:2.5 mgAPO 加入0.01g 抗坏血酸溶于10 mL 生理盐水(0.9 %)后混匀,全程避光、冰上配制;10 %水合氯醛:将10 g 水合氯醛溶于100 mL 蒸馏水中。
1.6 A2ARR//DD2DDRR实时荧光定量PPCCRR检测
检索Genbank 获得目的基因A2AR、D2DR 与内参βactin 基因real-time PCR 检测的引物序列(见表1),上述引物由北京奥科鼎盛生物公司合成。4 周运动干预结束后24 h,腹腔注射10%水合氯醛(0.35 g/100 g)麻醉大鼠,开颅取脑,迅速分离右侧纹状体背外侧部,提取总RNA,逆转录合成cDNA,进行荧光定量PCR 分析。根据PCR反应信号强度Ct值计算目的基因相对表达量,用2-△△Ct法计算目的基因相对于内参基因的mRNA 相对表达量,单个样本△Ct=三个复孔平均基因Ct-内参Ct,△△Ct=单个样本△Ct-假手术组△Ct 均值。

表1 基因引物序列(5’- 3’)
1.7 A2ARR//DD2DDRR蛋白表达水平的免疫荧光检测
4 周运动干预后24 h,10%水合氯醛(0.35 g/100 g)麻醉大鼠,固定、灌流、取脑、置于4%多聚甲醛中固定。将脑组织切片(40 μm)按顺序分为6 组,分置于6孔培养板内,漂浮于0.1 mol/L PB(pH=7.4)中,3%山羊血清、0.5%Triton X-100和0.1 mol/L PB(pH=7.4)封闭液中孵育 1 h,加入 A2AR 抗体(1∶50)、D2DR 抗体(1∶300)置4℃冷藏过夜。次日,PB 清洗3 次后,移入含Al⁃exa 488 荧光素化的山羊抗鼠二抗(1∶500)和Alexa 594 荧光素化的山羊抗兔二抗(1∶500)溶液中,室温条件下孵育 1 h,再用 0.1 mol/L PB 清洗 3 次,贴片,暗处风干,滴加50%甘油,加盖玻片完成标本制作。用荧光显微镜观察图片,从各组纹状体背外侧部区域和黑质致密部的相同层面选取5 张切片,统计阳性细胞数量,取平均值进行统计。
1.8 A2AR拮抗剂和D2DDRR激动剂干预行为测试
造模手术后第 14、28 天对 PD 组和 PD+Ex 组大鼠进行药物诱导旋转行为测试,方法如下:A2AR 拮抗剂SCH 和D2DR 激动剂QUIN 分别溶于二甲基亚砜(DMSO)中备用。QUIN 组和Ex+QUIN 组腹腔注射生理盐水(2 mL/kg),15 min 后腹腔注射QUIN(0.1 mg/kg);SCH+QUIN 组和 Ex+SCH+QUIN 组腹腔注射 SCH(2 mg/kg),15 min后腹腔注射QUIN(0.1 mg/kg);药物注射完毕后用旋转仪记录大鼠旋转圈数。
1.9 统计学分析
实验数据用SPSS 20.0 软件进行统计分析,结果以(±s)表示。采用双因素重复测量分析药物诱导旋转行为测试结果;采用Person 相关性检验分析纹状体A2AR-D2DR 表达量与药物诱导旋转圈数的相关性;采用方差分析(One-Way ANOVA)与Duncan 检验对RT-PCR、免疫组化、免疫荧光实验各组间均值进行比较,显著性水平为P<0.05。
2 结果
2.1 PPDD大鼠模型鉴定及行为学评价
APO 诱导旋转行为检测结果发现,第 7、14、28 天Control 组与Control+Ex 组大鼠均无明显旋转行为;注射6-OHDA 大鼠(n=116)中向健侧旋转圈数与损毁侧旋转圈数差值>100 r/30 min 共 76 只,PD 成模率约为65.5%,剔除其余未达标大鼠。第 7,14,28 天,PD 与PD+Ex 组大鼠净旋转圈数均显著高于Control 组(P<0.001),第7、14天PD+Ex 组净旋转圈数较PD 组有所下降但不具有显著性(P>0.05),第28 天PD+Ex 组净旋转圈数较PD组明显减少(P<0.05)(表2)。

表2 各组大鼠术后APO诱导旋转圈数变化(r/30 min)
TH 为DA 合成限速酶,存在于DA 神经元胞浆内,本研究采用免疫组化染色标记TH 的方法判定黑质DA能神经元数量和存活率。6-OHDA 损毁侧黑质致密部TH 染色如图1A~D 所示。Control组黑质 DA 神经元胞体染色深,数目多,纤维丰富,细胞边界清晰,形态规则(图1A);Control+Ex 组黑质DA 神经元数目较多,染色也略比Control 加深(图1B);PD 组黑质DA 神经元数目明显减少,胞体和纤维稀少(图1C);PD+Ex 组DA 神经元数目也较稀少,但部分着色较深,突起稍多(图1D)。各组损毁侧与健侧黑质TH 阳性表达如图1E 所示。PD 组和 PD+Ex 组较 Control 组明显减少(P<0.001),PD+Ex组较PD组明显增多(P<0.05)。
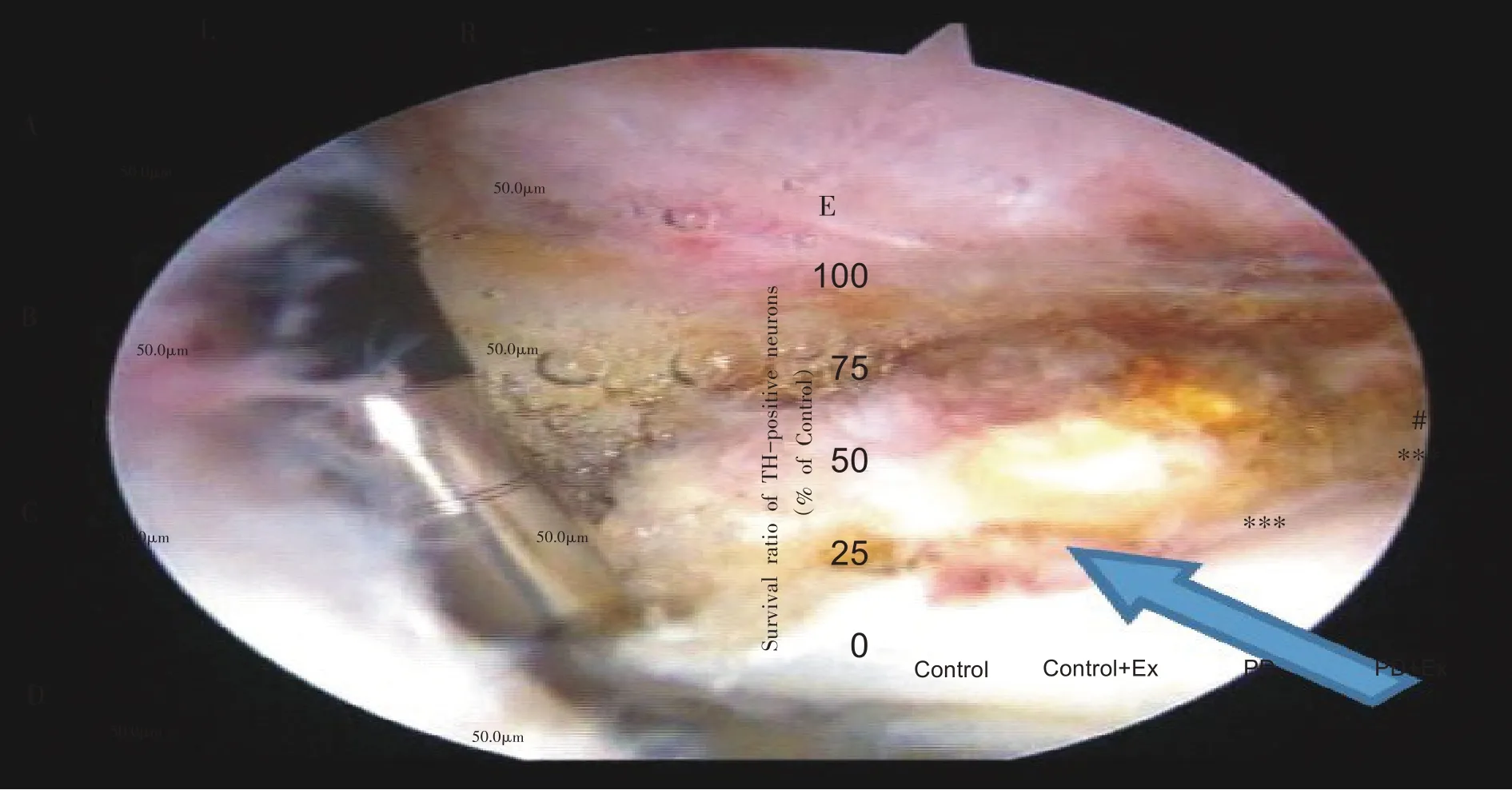
图1 各组大鼠黑质双侧TH阳性表达(n=6)
2.2 纹状体A2AR mmRRNNAA和D2DR mmRRNNAA表达水平的检测结果
各组大鼠纹状体背外侧部A2AR mRNA 检测结果显示,与Control 组比较,Control+Ex 组表达未出现显著变化(P>0.05),PD 组和 PD+Ex 组表达量均显著上调(P<0.01),但PD+Ex 组却较PD 组显著下调(P<0.05),如图2A 所示。纹状体D2DR mRNA 检测结果显示,与Control组相比,Control+Ex 组表达量有上调趋势,但未出现显著性变化(P>0.05),PD 组和PD+Ex 组出现显著下调(P<0.01),但PD+Ex 组却较PD 组出现上调,差异具有显著性(P<0.05),如图2B所示。

图2 大鼠纹状体A2AR和D2DR mRNA相对表达量的比较(n=8)
2.3 纹状体A2AR/D2DR蛋白表达水平的检测结果
各组大鼠纹状体背外侧部A2AR 及D2DR 蛋白表达水平如图3 所示,C 图中绿色荧光代表A2AR 蛋白表达(a),红色荧光代表D2DR 蛋白表达(b),黄色荧光代表A2AR 与 D2DR 共表达(c)。各组大鼠纹状体 A2AR 及D2DR 蛋白阳性细胞个数统计结果表明,与Control 组相比,PD 组纹状体A2AR 阳性细胞数量显著增加(P<0.01),PD+Ex 组较PD 组有所下降(P<0.05),但仍显著高于Control 组(P<0.01);PD 组纹状体 D2DR 阳性细胞数量显著减少(P<0.01),PD+Ex 组有所回升(P<0.05),但仍然低于Control 组(P<0.01);与Control 组比较,PD组纹状体A2AR-D2DR 共表达的细胞数量显著增加(P<0.01),PD+Ex组较PD组下降(P<0.05,图3D-F)。

图3 大鼠纹状体A2AR及D2DR蛋白表达水平比较(n=8)
2.4 A2AR拮抗剂和D2DR激动剂诱导PD大鼠旋转行为检测结果
造模手术后第14、28 天,药物诱导PD 大鼠旋转行为检测结果如图4 所示。造模手术后第14 天,与QUIN组相比,SCH+QUIN 组大鼠向健侧旋转圈数显著增多(P<0.01),Ex+ QUIN 组大鼠向健侧旋转圈数无明显变化(P>0.05)。与Ex+QUIN 组相比,Ex+SCH+QUIN 组大鼠向健侧旋转圈数显著增多(P<0.01);与SCH+QUIN 组相比,Ex+SCH+QUIN 组大鼠向健侧旋转圈数显著降低(P<0.01)。造模手术后第28 天,EX+QUIN组大鼠向健侧旋转圈数显著低于QUIN 组(P<0.05,图4B)。各组大鼠28天旋转圈数较14天均有增加的趋势(图4C)。
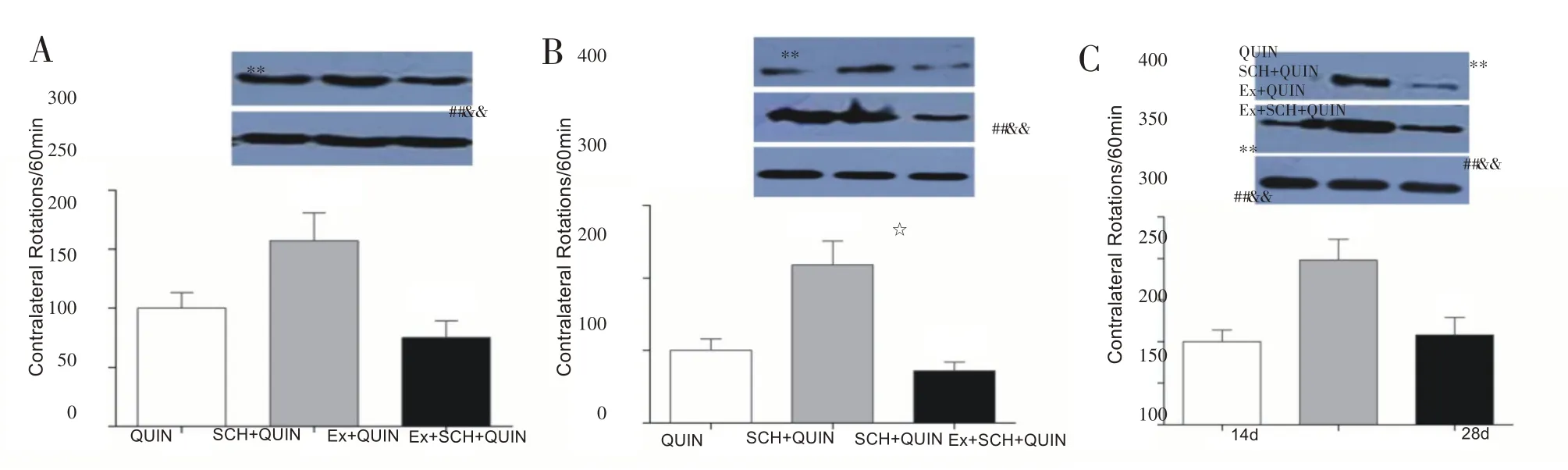
图4 大鼠向健侧旋转圈数的比较(n=8)
2.5 纹状体A2ARR--DD2DDRR 表达量与药物诱导旋转圈数的相关性分析
为评价纹状体A2AR-D2DR 表达水平与PD 大鼠行为障碍之间的关联性,本研究将PD 大鼠纹状体A2ARD2DR 阳性细胞数与药物诱导旋转圈数进行Person 相关性检验,结果显示,PD 与PD+Ex 组大鼠注射SCH58261 和Quinpirole 后向健侧旋转的圈数与A2ARD2DR阳性细胞数成正相关(P<0.01),如图5所示。

图5 PD大鼠纹状体A2AR-D2DR表达与药物诱导旋转行为的相关性
3 讨论
3.1 6--OOHHDDAA 药物对大鼠黑质DDAA 能神经元的毒性损伤作用
PD 是以运动功能障碍为主要临床特征的神经系统疾病,主要表现为运动不能、迟缓、肌强直、姿势步态不稳、静止性震颤及不同动作协调模式之间转换能力降低等[8]。PD 发病比较隐匿,当上述症状出现时黑质DA 能神经元丢失已达60%,因此,临床上将运动功能评分作为判断PD 患者DA 能神经元存活率的参考依据[9]。6-OHDA 为选择性儿茶酚胺类神经化学损毁剂,进入体内后易氧化生成羟自由基和醌类等神经毒物,特异性地损毁黑质DA能神经元,引起DA生成减少,黑质-纹状体DA系统功能退变,产生与PD患者相类似的一系列运动功能障碍,如损毁侧运动减少、身体向损毁侧偏斜、四肢平衡及协调性降低等[10]。因此,6-OHDA大鼠是目前公认的一种用于PD 病理学和药理学研究的理想PD动物模型。
已有研究证实,6-OHDA 注射后24 h出现黑质DA神经元变性,2~3 天后出现纹状体DA 含量减少,成模后 14 天 DA 含量约减少 80%~90%[11]。6-OHDA 染毒导致PD 大鼠纹状体中等多棘神经元(medium spiny neurons,MSNs)突触后膜上 D2DR 超敏,此时若给予D2DR 激动剂或促DA 释放剂等药物可诱导大鼠出现异常向健侧旋转的行为[12]。鉴于此,本研究在注射6-OH⁃DA 药物7 天后,采用APO 诱导大鼠旋转行为,并以30 min 内大鼠旋转次数大于100 r 作为判断标准,成功复制PD 大鼠模型,成模率约为65.5%。TH 免疫组化染色观察发现,PD 组和PD+Ex 组大鼠黑质致密度DA 能神经元着色较深,数量明显减少,胞体和纤维稀少,说明6-OHDA药物造成了DA能神经元不同程度损伤。
3.2 运动对PPDD模型大鼠黑质DDAA能神经元的保护作用
长期从事规律的体育活动能够有效降低PD 发病率[13]。太极拳、拳击、探戈、抗阻力练习等运动康复手段对PD 患者行为功能异常产生积极影响[14,15]。自主转轮运动、强迫跑台运动、游泳及综合运动训练等方法在改善啮齿类PD 动物行为功能障碍方面也积累了一些有效数据[16,17]。本实验室前期研究证实,4 周运动干预显著改善了PD 大鼠前肢运动不对称性[18]、四肢协调性和自主活动能力[19]。本研究采取Tajiri 等[7]跑台运动干预方案,运动干预介入时间选择在6-OHDA 注射后24 h,证实早期运动干预达到缓解6-OHDA 药物对黑质DA 能神经元毒性损伤的效果。另有研究证实,运动可降低PD 模型大鼠黑质DA 能神经元的死亡率,上调纹状体TH 阳性表达[20],与本研究结果一致。本实验室前期研究还发现,4 周跑台运动干预通过上调PD 大鼠纹状体D2DR 表达,抑制黑质DA 能神经元放电频率和爆发式放电活动,改善黑质-纹状体DA 神经通路的功能[21,22];其原因与运动的神经保护作用减轻了 6-OHDA 药物对黑质-纹状体通路DA 能神经元的毒性损伤,增加了DA合成与分泌,进而调节了黑质-纹状体DA能神经传递过程有关[23]。
3.3 运动调节纹状体A2ARR//DD2DDRR 表达改善PPDD 大鼠的行为功能障碍
纹状体是基底神经节直接与间接运动调节通路的起始部位,其神经元构筑约95%为GABA 能MSNs,分两类亚型:即表达D1DR 的GABA 能强啡肽神经元和表达 D2DR 的 GABA 能脑啡 肽神经 元[24]。 A2AR/D2DR 共表达于纹状体D2-MSNs 突触后膜[25],均属G 蛋白偶联受体,对腺苷酸环化酶(AC)产生互为拮抗作用;激活A2AR 兴奋纹状体-苍白球通路GABA 能神经元、激活间接通路,起到抑制运动的作用[26];相反,激活D2DR 抑制纹状体-苍白球通路GABA 能神经元、抑制间接通路,起到易化运动的作用[27]。
Katia 等[28]发现PD 患者血浆中腺苷及其代谢物浓度高达5.1 ± 0.4 μM,而健康人仅为2.0 ± 0.2 μM;腺苷浓度大幅度增高易使A2AR活性增强、密度增加,可刺激下游cAMP 生成增多,兴奋纹状体-苍白球通路GA⁃BA 能神经元、过度激活间接通路,是引起PD 患者运动障碍的脑内分子机制之一。PD 动物纹状体D2DR 表达下调,易感性增强[29];A2AR 激动剂可显著削弱6-OHDA大鼠纹状体MSNs突触后膜上超敏的D2DR 与其激动剂的亲和力,抑制D2DR 活性[30]。由此推测,PD 大鼠纹状体D2DR 信号减弱与A2AR 过度激活有关,纹状体A2AR与D2DR 表达异常是导致直接与间接通路调节失衡,引起PD大鼠运动功能障碍出现的主要原因之一。
本研究结果显示,4 周运动干预下调了PD 大鼠纹状体A2AR mRNA 和蛋白表达,上调了D2DR mRNA 和蛋白表达,并下调A2AR-D2DR 表达。Costa 等[31]研究发现,1 次/周、3 次/周和 7 次/周跑台运动均可下调老年大鼠海马A2AR 蛋白表达。上述研究结果说明,运动干预起到类似A2AR 拮抗剂的作用。咖啡因隶属于非特异性A2AR 拮抗剂,作用于纹状体D2-MSNs 突触后膜调节A2AR 表达,抑制AC 和cAMP 生成及纹状体-苍白球通路GABA 能神经元的兴奋性,改善了PD 大鼠的行为功能[32]。近些年来A2AR 拮抗剂已作为PD 病人药物治疗的新靶点[33]。流行病学研究显示,人群原发性PD 发病率与咖啡因摄取量呈负相关,6-OHDA大鼠给予2周低剂量咖啡因干预后自主活动能力明显改善[34]。本研究采用A2AR拮抗剂和D2DR激动剂药物干预进一步证实,运动通过抑制纹状体A2AR对D2DR的拮抗作用,减弱胞内AC-cAMP-PKA 信号转导通路活性,抑制D2-MSNs过度激活,促进黑质-纹状体DA能突触传递的可塑性,进而调节直接/间接通路失衡状态,达到改善PD 行为功能障碍的治疗效果。
4 总结
早期运动干预可减轻6-OHDA 药物对大鼠黑质DA 能神经元的毒性损伤,调节PD 模型大鼠纹状体A2AR 及D2DR 基因转录及蛋白表达,改善PD 模型大鼠的行为功能障碍。本研究初步证实纹状体A2AR/D2DR是运动纠正PD 大鼠基底神经节直接与间接通路失衡的重要细胞分子靶点,其机制可能与运动的神经保护作用改善了PD 大鼠黑质-纹状体DA 能通路的突触可塑性有关。采用全细胞膜片钳与光遗传技术实现对细胞膜分子的精准操控,进一步阐明纹状体A2AR/D2DR在运动纠正PD 基底神经节直接与间接通路失衡中的作用机制将是本研究团队今后的工作重点。
猜你喜欢
中国体育科技(2022年1期)2022-03-10
中国康复(2021年6期)2021-11-30
青岛大学学报(医学版)(2021年5期)2021-11-17
科学与财富(2020年24期)2020-10-27
浙江医学(2020年9期)2020-07-01
青岛大学学报(医学版)(2020年2期)2020-06-08
首都体育学院学报(2019年5期)2019-10-18
家庭医药·快乐养生(2018年9期)2018-09-27
中国医药导报(2018年18期)2018-09-26
恋爱婚姻家庭·养生版(2018年12期)2018-01-15
