
土壤生物与可持续农业研究进展
2020-07-02 01:23张卫信申智锋邵元虎时雷雷刘胜杰史楠楠傅声雷
生态学报 2020年10期
张卫信,申智锋,邵元虎,时雷雷,刘胜杰,史楠楠,傅声雷
1 黄河中下游数字地理技术教育部重点实验室;河南大学环境与规划学院, 开封 475004 2 中国科学院西双版纳热带植物园, 勐腊 666303
生态文明已成为我国国家发展战略之一,而可持续农业是生态文明建设的必然要求和重要内容。2015年5月农业部等印发了《全国农业可持续发展规划(2015—2030年)》,分析了目前我国农业发展取得的成就和面临的严峻挑战,指出农业关乎国家食物安全、资源安全和生态安全,并把我国分成了农业优化发展区、适度发展区和保护发展区。2017年9月,国家又出台了《关于创新体制机制推进农业绿色发展的意见》,认为:“推进农业绿色发展,是贯彻新发展理念、推进农业供给侧结构性改革的必然要求,是加快农业现代化、促进农业可持续发展的重大举措,是守住绿水青山、建设美丽中国的时代担当,对保障国家食物安全、资源安全和生态安全,维系当代人福祉和保障子孙后代永续发展具有重大意义”;2017年11月,中国科学院相关专家建议在雄安新区成立国家绿色先进农业研究院[1]。可持续农业正迎来重要的发展契机。
土壤是农业发展的基础,而土壤生物是土壤健康的关键;农业的可持续发展,必须充分发挥土壤生物的积极作用。土壤生物之间、生物与非生物因子之间的复杂和多维的互作关系,是生态学的前沿领域和难点,也是与可持续农业相关的基础研究的重要突破口。农业的可持续发展是人类生存和发展的基础,以高投入维系高产出的工业化农业面临不断增加的经济和环境压力[2]。最近50年,土壤生物多样性对生态系统服务的基础性贡献得到日益重视[3]。土壤生物多样性在抑制土传病害,保障清洁健康的大气、水体和食物等方面都有重要的作用;通过不断优化管理措施进而提高生态复杂性和土壤生物多样性的稳健性(Robustness)关乎人类福祉,但这一潜力远未被充分发掘[4]。如何充分发挥农业生态系统各组分间天然的协作关系[5],提高整个系统的资源总量及其能量、水分和养分利用效率,降低能量内耗及养分淋失,是可持续农业发展的迫切需求。土壤动物和土壤微生物是土壤结构和肥力的重要驱动力,也是生态系统内部平衡的重要调控者。打开土壤“黑箱”,被喻为陆地生态系统研究的“最后的前沿”[6]。
土壤生物与可持续农业已成为后工业化农业的重要研究方向和重大挑战。可持续农业事实上已经超出了农业领域,成为人类社会文明发展的重要内容。本文仅从农业生产本身涉及的科学和技术层面梳理土壤生物和可持续农业的学科发展历史,总结土壤生物与农业管理措施的关键联系,探究土壤生物在农业生态系统中的生态功能;最后,对如何充分发挥土壤生物在现代农业体系中的作用进行了思考。我们认为土壤生物群落、土壤有机质和土壤结构等的演变是影响可持续农业的关键因素;所以本文针对的科学问题为:它们三者对不同农业模式如何响应?土壤生物如何调控土壤有机质、土壤结构和食物网内的生物互作,进而促进可持续农业?
1 研究历史概述
1.1 可持续农业的提出
工业革命以来,人口的持续增长、地球资源的快速消耗以及工农业化学残留的大量累积等,对人类生存环境造成了空前的破坏。为实现“自然平衡、经济平衡、社会平衡”的伟大目标,可持续发展的思想应运而生[7]。20世纪80年代,发达国家率先提出了可持续农业(Sustainable agriculture)的发展战略[8]。1991年,联合国粮农组织把可持续农业的基本内涵概括为:“通过重视可更新资源的利用,更多地依靠生物措施来增进土壤肥力,减少石油产品的投入,在发展生产的同时保护资源改善环境并提高食物质量,实现农业的持续发展”[9]。
可持续农业是在传统农业,工业化农业,及以有机农业﹑生物农业和生态农业等为代表的替代农业模式的基础上,贯彻可持续发展的思想的基础上形成的。农业生产模式的发展是人类社会发展的重要内容,有相似的发展规律。两者都是基于对原有模式的部分否定,否定之否定,进而螺旋式演进的(表1)[10-13]。具体说来,原始农业极度依赖自然,生产力极端低下,但人类活动对自然的干扰也小[10];工业化之前的传统农业,人的主导作用明显加强,对自然的影响较大,生产力明显提高但仍然“靠天吃饭”[11];工业化农业过度依赖人类及其创造的各种工具,短期内生产力高,但对自然资源的消耗过大,并深刻地改变了自然环境,可持续性差[12];替代农业从各自的角度出发,希望找到对自然影响较小的现代农业生产方式,但仍存在短期内生产力偏低等诸多不利因素[12-13]。只有发挥各种农业生产模式的优点,积极应对其缺点,才可能真正实现农业的可持续发展。可持续农业与原有农业模式并非截然不同,它是所有农业生产模式的集大成者。可持续农业可以定义为:“综合运用现代科学和管理的理论和技术,充分发挥农业生态系统中“植物-土壤-其他生物”天然的协作关系,提高资源利用效率,减少外部资源投入,并长期维持较高的种群或群落生产力和产量的现代农业体系”。

表1 农业发展的主要阶段及其主要特征
作为可持续农业的重要代表,最近20年,有机农业在全球范围内获得了快速发展。在1999年至2016年间,全球有机农产品的年销售额已经增长了6.2倍,达到897亿美元;有机农业用地10年间增加了近一倍,到2016年达到5780万公顷[14]。Reganold 和 Wachter[15]在NaturePlants从生产力、环境影响、经济效益和社会福祉等4个方面对有机农业做了综述。该研究发现从总体上看,有机农业的单位面积产量稍低于传统农业,但它拥有利润更丰厚、对环境更友好及食品安全性更高等优势。不过,当前有机农业仍处于起步阶段,其体量仅占全球农业用地的1.2%。有机农业单产偏低的不足,是制约其发展的瓶颈之一,也恰恰是土壤生物可能发挥积极作用的地方。
1.2 土壤生物与可持续农业的学科发展
可持续发展的概念框架中,并不是一开始就充分考虑了土壤生物的重要贡献。这可以从土壤生物学和可持续发展相关的专业期刊的创建时间清晰地看到。3种主要的土壤生物学相关期刊早在可持续发展的概念提出之前就已经创刊,而真正意义上的可持续农业的专业期刊,直到1990年才正式出版。但土壤生物在近年来得到了格外的关注(表2)。2014年,Elsevier 发行了第一本以食物网为研究焦点的期刊FoodWebs;2016年,又发行了Rhizoshere以促进有关植物和土壤的关键连接点的研究,发挥根际圈在维持生态系统生产力的可持续性和食物供给安全及应对气候变化等领域的积极贡献[16-17]。有意思的是,可持续发展相关的期刊,其关注的主题很少直接涉及土壤生物,但土壤生物相关的期刊,则往往会明确关注可持续农业。
土壤生物在生态系统物质循环和能量传递中的贡献得到日益重视。但是,土壤生物在可持续农业中的关键地位没有得到足够的认可,人们对土壤生物多样性的保护意识也很低[18];而在实践中应用土壤生物去调控农业生态系统过程,更是刚刚开始尝试。美国科学院关于21世纪的可持续农业的报告,只提到了有机农业对土壤生物的利好作用(如提高了蚯蚓和土壤微生物等的生物量),以及生物防治在有机农业中的应用,却没有提及如何利用土壤生物多种多样的功能促进可持续农业的发展[19]。2016年出版的GlobalSoilBiodiversityAtlas一书,虽然在科普层面促使人们更多地关注土壤生物[20],但在科学研究上并无太多的贡献。以 Elsevier 的文章为例,2015年之前,土壤生物与可持续农业相关文章少于50篇每年,而2015年之后则有明显的增长,每年发表的文章接近80篇(图1)。值得一提的是,从1998年至2016年,由 Clive A. Edwards 和 Stephen R. Gliessman 主编的系列丛书AdvancesinAgroecology陆续出版了21本可持续农业相关的专著。其中两部著作:MicrobialEcologyinSustainableAgroecosystems[21]和SoilOrganicMatterinSustainableAgriculture[22]从不同角度阐述了土壤生物在可持续农业中的作用。可见,最近20年土壤生物与可持续农业相关研究已经开始得到重视。
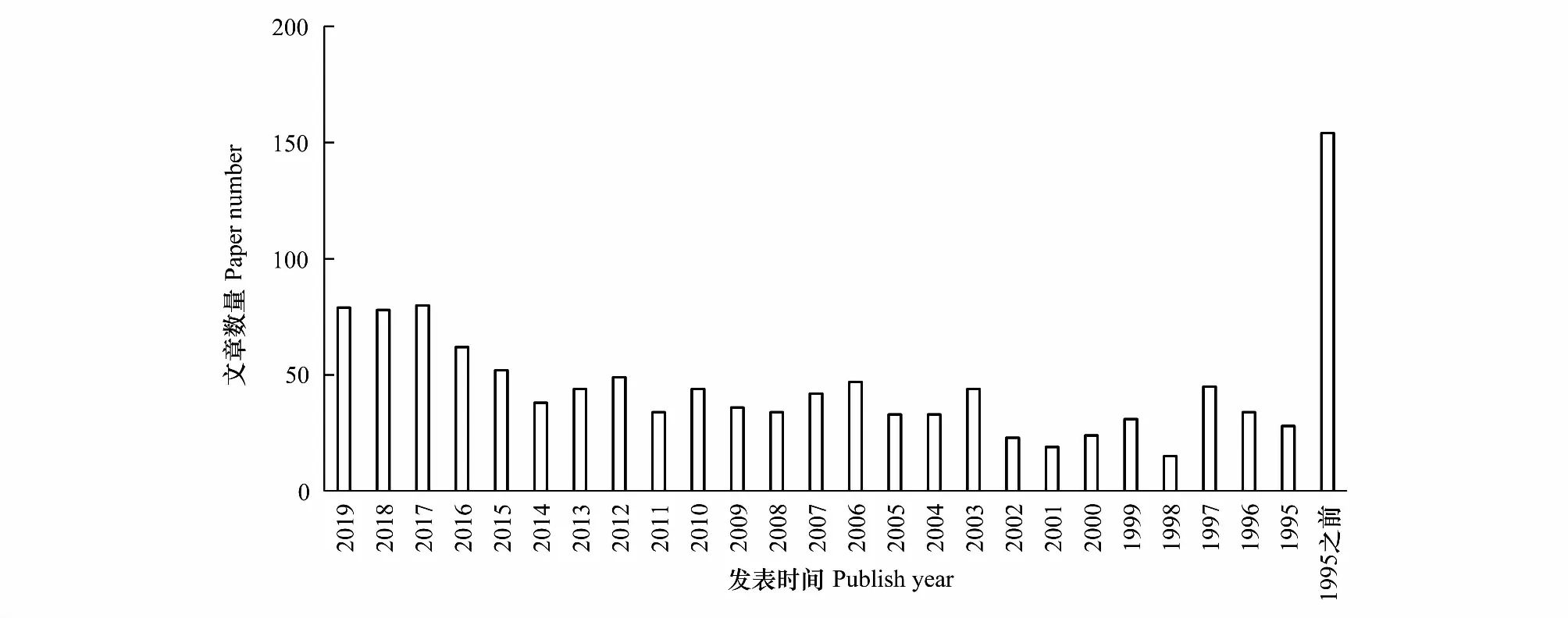
图1 Elsevier 刊发的与土壤生物与可持续农业紧密相关的文章数量Fig.1 The number of papers that closely related to soil biota and sustainable agriculture published in Elsevier journals以“soil fauna AND soil biota AND sustainable agriculture” 为检索词,全文检索,检索时间为1974年1月至2019年7月29日
国内的农业生态学在20世纪80年代兴起,90年代开始快速发展,至2005年每年发表的中文文章即已超过1000篇[23],但是,土壤生物在可持续农业中的作用长期未得到重视。直到2013年,土壤生物学科发展被纳入国家自然科学基金委员会-中国科学院学科发展战略研究项目[24]。中国农业科学院基于中国农田土壤肥力长期试验网络,探究了长期施肥、秸秆还田及保护性耕作等农田管理措施对土壤肥力、土壤结构和pH以及作物产量等的影响,部分工作也涉及到农田土壤微生物和酶活性以及土壤动物群落对施肥的响应[25-26]。从CNKI的检索结果来看,国内土壤生物与可持续农业相关的研究集中在土壤肥力及于此相关的土壤养分和有机质、土壤微生物和酶活性等方面,而土壤动物研究仅占1.97% (图2)。并且,多数研究集中于对农业生态系统中土壤动物群落的种类组成和生物量数量特征及其对耕作和管理措施的响应上[27-30]。值得注意的是,国内学者已基于长期小区试验研究了蚯蚓对农田有机碳动态及土壤微生物等的影响,发现蚯蚓活动可增强土壤团聚作用,改变土壤微生物群落,提高养分有效性并影响光合产物在植物-土壤系统中的分配[31-34]。但是,土壤动物野外控制试验仍十分缺乏,且仍然很少探究土壤动物食性、肠道微生物及粪粒等与其生态功能密切相关的特征及其可能的影响因素。2015年,《中国科学院院刊》也组织出版了土壤与生态安全专刊,综述了土壤物理学[35]、土壤化学[36]和土壤微生物学[37]等的研究现状与展望,及土壤与可持续农业生产领域的现状与战略[38-40]。但是,该专刊并未包含土壤动物学研究的内容;土壤动物及其在农业中的作用研究,在国内还需要更多的关注。以蚯蚓为例,在土壤科学英文文献中,“Earthworm”一直属于排名前20位的高频关键词,它在1986—1995年,1996—2005年和2006—2014年间的文献中分别出现54次,258次和299次;但是,“蚯蚓”却从未成为土壤科学中文文献的高频关键词[41]。2017年,《中国科学院院刊》又组织出版“科技促进农业供给侧结构性改革”专刊,围绕秸秆综合利用[42]、土壤肥力调节[43]、都市现代农业[44]和生态农牧场建设[45]等可持续农业主题开展了深入的讨论。但是,该专刊仅对微生物资源(如微生物肥料)的利用有所涉及,没有对土壤动物及整个土壤食物网在我国“绿色革命”[1]实践中应有的贡献进行深入讨论。2018年2月,《中国科学院院刊》再次组织了“土壤与可持续发展”专题,针对土壤资源保护、土壤污染防治等方面存在的问题探讨了相关解决方案;但仅仅简短地提及了土壤生物多样性的保护[46]。目前,我国土壤动物与土壤污染治理相关研究还处于起步阶段,主要围绕蚯蚓及其蚓触圈的贡献展开[47-50]。总之,土壤生物与可持续农业正迎来学科发展的重要契机,但是,土壤生物特别是土壤动物在可持续农业生产中的积极作用应该得到进一步的发挥。

表2 土壤生物和可持续农业相关英文期刊的发展脉络

图2 CNKI中土壤生物与可持续农业相关的研究主题分布格局(1979.6—2019.03)Fig.2 CNKI-based analysis of the distribution pattern of research topics on soil biota and sustainable agriculture during periods of Jun 1979 to Mar 2019此图为以“农田,施肥,秸秆,有机肥,土壤动物,土壤微生物”为检索词在CNKI中全文检索所得数据
2 土壤生物与可持续农业研究概述
可持续农业与土壤过程的关键联系大体包括三方面:即1)土壤结构,与根系活动、土壤生物群落特征和土壤有机质含量等紧密相关。2)病虫害,与土壤食物网内部及植物-土壤生物的互作有关。3)土壤肥力,与土壤有机质含量、土壤生物群落特征及土壤理化环境等有关。可持续农业研究的重要内容是探究农业模式对土壤生物群落特征、土壤结构和土壤有机质及三者的互作过程的影响。并且,分析土壤生物对土壤结构、病虫害和有机质动态的调控作用,以揭示土壤生物在可持续农业中的贡献,构建因地制宜的现代农业模式。
2.1 农业模式对土壤生物群落及相关生态过程的影响
2.1.1土壤生物群落对农业管理措施的响应
有机农业和免耕等替代农业模式和传统耕作模式对土壤生物群落及相关的地上地下生态过程的影响差别很大。建于1978年的瑞士长期农业实验 (DOK trial, Switzerland),对2种有机农业系统和2种传统农业系统进行了长期系统的比较研究。Mäder等[5]在Science发表文章,初步总结了持续21年的有机和传统农业措施对作物产量、土壤结构和土壤食物网特征的影响。结果表明,有机农业增加了土壤团聚体的稳定性、土壤微生物生物量、菌根侵染率、蚯蚓数量、蜘蛛等捕食者数量,提高了微生物多样性并降低了微生物代谢熵。相应地,有机农业措施使有机质分解更彻底、养分淋失风险更低、微生物对资源的利用效率更高;有机农业系统的外部养分输入比传统耕作系统低34%—51%,但产量仅低了20%,而且其生产单位干物质所需的能量也比传统耕作系统低20%—56%。Birkhofer等[51]在该实验平台的研究进一步表明:有机农业系统及施用有机肥(可以是传统农业系统)提高了土壤微生物生物量及有机质含量,促进了食细菌线虫和蚯蚓的生长,以及捕食者数量的增长,进而促进了养分循环和害虫控制。相反,施用化肥和杀虫剂,则削弱了自然的捕食作用对害虫的控制,并减弱了系统内部的生物循环。Birkhofer等[52]在专著中进一步总结了瑞士DOK实验发现:经过27年的野外实验,农田土壤革兰氏阴性菌因施用无机肥而受抑制,因施用有机肥而增加;菌根真菌的侵染率在传统耕作的农田中明显降低,结果导致在传统耕作的农田中土壤革兰氏阳性菌居于主导地位。与传统认识不同的是,该研究发现大个体的土壤动物,如线蚓、蜘蛛和地表甲虫,对耕作强度的响应明显弱于细菌、真菌和线虫等小个体类群,认为可能与后者的移动性差有关。
植物-土壤生物互作对耕作模式响应明显。Coleman 等[53]通过14CO2标记实验发现:在传统耕作的玉米田中,小型节肢动物的14C丰度很快就接近根系的14C丰度,即快速地获得了玉米根系输入的碳;但是,在免耕的玉米田中,小型节肢动物对根系碳的利用要少很多。他们认为,上述结果可能说明在免耕系统中小型节肢动物从未被标记的有机质(如凋落物层的真菌)获得了大量的食物,而相反,在传统耕作系统中,它们不得不更加依赖作物根系。该研究还发现耕作方式会改变地上地下植食者的联系。在传统耕作的玉米田中,蝗虫对玉米植株的取食作用并不影响线虫对根系来源碳的利用;但是,在免耕系统里中等强度的蝗虫取食作用可以促进线虫对根系来源碳的利用,而且杂草的根系可能对土壤动物的生存也有十分重要的意义。
耕作措施对土壤微生物多样性和土壤动物多样性的影响规律并不一致。一方面,土壤微生物与土壤动物的响应不同。Coleman 等[54]发现农田中细菌多样性甚至可以明显高于农田附近的林地,这可能与伴生的高多样性的杂草有关,但是保护性耕作往往可以提高土壤动物多样性。Callaham Jr. 等[55]也发现随着干扰强度的增加(阔叶林、松林、草地和耕地)大型土壤动物多样性逐渐下降。传统耕作系统向保护性耕作系统的转变,也可提高跳虫多样性及跳虫的密度[56]。另一方面,不同土壤微生物类群对耕作方式的响应也不同。Moore和de Ruiter[57]发现土壤耕作过程对真菌能流通道的不利影响要远大于其对细菌能流通道的潜在不利影响。我们以为,传统耕作系统中真菌通道的相对削弱可能是农田细菌多样性升高的原因之一。可见,土壤生物多样性的驱动因子并不简单,有待深入研究。最后,由于耕作措施复杂多样,免耕或保护性耕作的“内涵”不统一,其对土壤食物网结构和功能的影响,还很难得到一般性的结论。例如,在阿根廷主要农区之一的Pampas地区,为减少抗除草剂大豆的生产成本,实行的免耕措施伴随着大量的除草剂的使用,这与联合国粮农组织(FAO)倡导的免耕方式有很大不同,导致研究结果不好与其他研究进行比较[58]。
2.1.2代表性土壤动物类群对农业管理措施的响应
蚯蚓个体大,食性复杂,几乎参与了所有地上地下关键过程,在土壤食物网中常有“牵一发而动全身”的特殊地位[59];线虫数量多,分布广,可以与微生物和植物构成一个完整的微食物网[60]。所以,利用蚯蚓和线虫群落特征的变化反映土壤食物网特征随耕作措施的变化规律,有独特的优势。Liang等[61]研究了中国东北20年长期施肥样地的线虫变化,发现施有机肥使土壤线虫数量明显增加,且该土壤食物网以细菌分解通道为主。叶成龙等[62]也发现施用有机肥的麦田中细菌分解途径处于土壤腐屑食物网的主导地位。刘婷等[63]则发现在我国南方稻麦轮作体系中,施猪粪2年后土壤线虫多样性显著提高,但线虫通路比值未变。Culman等[64]比较了美国堪萨斯长期未施肥但长期割草并移走的多年生草地与长期依赖外部高输入维持的作物大田中线虫食物网特征。发现相对于作物大田,草地土壤中真菌分解通道明显加强,植食性线虫数量更少,且食物网的复杂性和稳定性更高;说明草场土壤肥力更高和生物群落更复杂,也部分解释了为何该草场经历70多年的草料收获仍然可以维持很高的生产力。我们以为,该天然草场得以可持续利用的重要原因,可能是收获草料的时候,并未破坏土壤地被物层,而后者是健康的土壤食物网的重要保障。当然,若线虫群落结构太过退化,其对耕作方式的指示作用也可能不准确。例如,Minoshima 等[65]发现免耕提高了土壤微生物生物量并促进了土壤食物网真菌通道的发展;然而基于线虫群落表征的土壤食物网结构未明显变化,而后者可能由于长期耕作已使高营养级的线虫消失殆尽所致。
蚯蚓是另一个与耕作措施紧密相关的代表性类群。在斯洛伐克,耕地的蚯蚓数量和生物量仅为永久草地蚯蚓的一半[66]。Briones 和 Schmidt[67]对全球40多国在1950—2016年间开展的耕作强度对蚯蚓群落大小和结构影响研究进行了Meta-analysis分析。可以看到,欧洲和北美的研究最多,其次是非洲和大洋洲,而亚洲相关的研究最少。该研究发现:免耕或保护性耕作可使蚯蚓数量和生物量增加至传统耕作农田的1—2倍;而且,免耕中常配套使用的草甘膦(Glyphosate)并未显著影响蚯蚓种群大小;而蚯蚓的活动的增强,反过来可作为“自然之犁”而改善土壤结构及养分循环。传统耕作的农田中,杀虫剂的大量使用、食物的短缺及可能的机械伤害,使蚯蚓生存条件恶化,种群偏小[68],进而限制了蚯蚓对作物生长的积极作用。蚯蚓的种群大小还可能受到植物分泌物和农药的综合影响。Ashworth 等[69]发现,连续种植棉花,可能因为过多施用杀虫剂或分泌大量的棉籽酚而使蚯蚓群落大小降至该地平均水平的50%以下,而连续种植大豆或玉米、或玉米大豆轮作等则能维持较高的蚯蚓数量;并且,棉花-玉米轮作或大豆-棉花轮作可消除种植棉花对蚯蚓的不利作用。河北曲周盐渍化改造区农田及山东恒台高产农田生态系统蚯蚓群落结构特征的研究结果也表明合理的投入特别是农田有机物投入对蚯蚓种群维持及其演替过程十分重要[30, 70-71]。可见,适当地改进耕作措施,可以调控蚯蚓群落大小;深入探究其中的机制,对发挥土壤生物的积极作用,意义很大。蒋高明等[72-73]利用秸秆养牛、牛粪堆肥回田等措施提高了土壤生物多样性,蚯蚓数量从16 条/m2增至317条/m2;该团队还比较了施牛粪堆肥和化肥对小麦-玉米轮作系统的影响,发现单施化肥不仅降低了土壤有机质和总氮含量,且对蚯蚓不利,但是添加牛粪堆肥可明显减轻因施化肥而产生的副作用。
2.1.3土壤生物、土壤结构和土壤有机质对农业模式的响应速度
同样建于1978年的美国佐治亚 Horseshoe Bend实验,对免耕和传统耕作系统中的土壤结构、土壤生物和有机质等对耕作模式的响应速度进行了比较研究。结果表明:土壤微生物群落、土壤结构对免耕的响应快于土壤有机质的累积。随着免耕的实施,土壤微生物群落结构呈非线性的变化,且这种响应非常迅速。相对于传统耕作的农田,30年的免耕措施可提高0—5 cm表土的团聚体的稳定性,而农田附近自然演替的森林可提高 0—28 cm 土壤团聚体的稳定性;但是仅仅表土中的土壤有机质因免耕或自然森林演替而提高[74-75]。可见,土壤有机质的积累要比土壤结构的改善需要更长的时间。这也可能是导致土壤养分循环效率和作物产量的提高相对滞后于土壤食物网的变化[76]的原因之一。当然,整个土壤生物群落对耕作方式的响应仍然需要较长的时间。Adl等[77]对美国东南部的免耕4—25年的棉花田的土壤生物多样性进行了比较。结果发现,只有免耕8年以上的棉田,其土壤生物多样性和多度才比较接近未受干扰的生态系统。土壤生物、土壤结构和土壤有机质的逐次恢复过程可以给我们几点启示:1)恢复土壤有机质是可持续农业的难点;2)评价农田土壤质量需要综合考虑土壤生物、土壤结构和土壤有机质的变化;3)通过因地制宜的秸秆还田和施有机肥等措施提高土壤有机质含量,仍然是可持续农业管理的重要突破口。
2.2 土壤生物在可持续农业中的作用
2.2.1土壤生物对土壤结构的影响
土壤生物对土壤结构和相伴的水分动态等的调控作用很早就被关注[78-79]。土壤动物,特别是被称为“生态系统工程师”的蚯蚓[80-81],可以通过取食、掘穴、排泄和搬运等活动改变土壤的孔隙度、渗透性和团粒结构,形成生物活性及土壤理化特性迥异的各类“资源岛”和关键界面[82-84]。近年来,土壤微生物对土壤结构的影响也得到重视。Six 和 Paustian[85]总结了耕作方式对团聚体的影响,同时强调了蚯蚓和真菌对团聚体形成的重要作用。但是,最新的一项Meta-analysis研究则认为,相对于土壤动物来说,细菌和真菌对土壤团聚体形成的作用更为重要[86]。该研究还发现,虽然有时候土壤动物和真菌不利于土壤团聚体形成,但土壤生物总的多样性对土壤团粒结构的形成有重要的意义。
土壤动物对土壤结构的影响显而易见。以蚯蚓和马陆为例,它们在旱作农田中十分常见,其活动对土壤结构及相关过程均有重要的影响。蚯蚓被达尔文喻为“大地之犁”[87]。蚯蚓及其蚓触圈(Drilosphere)对土壤结构有深刻的影响[59, 88]。蚯蚓活动导致的生物扰动(Bioturbation)甚至被认为对亚马逊黑土(Amazonian Dark Earths)的形成也有很大的贡献[89]。蚯蚓活动在多数情况下倾向于降低土壤容重,但少数情况下也会提高土壤容重[90]。在大棚蔬菜土壤中养殖蚯蚓4个月,即可提高土壤孔隙度10%[91]。表栖类(Epigeic)蚯蚓的取食过程,可以显著改变地表凋落物层的厚度甚至是林下植被组成[92],进而影响土壤过程。深栖类(Anecic)蚯蚓体型大,可以挖掘连接到地表的垂直洞穴从而有利于水分流通和气体扩散[93]。然而,当土壤含水量很高的时候,有些内栖类(Endogeic)蚯蚓的活动还可能转而降低土壤的孔隙度及透气透水状况(图3)。

图3 南美岸蚓对土壤结构和水分动态的影响(申智锋供图)Fig.3 The impacts of earthworm (Pontoscolex corethrurus) on soil structure and moisture dynamic (photo by Zhengfeng Shen)
Snyder 和 Hendrix[94]认为马陆是未来可用于促进生态恢复的重要土壤生物类群。有两方面的特性使马陆在土壤改良和生态恢复中可能起着重要的作用。其一,马陆对土壤结构的改良作用。类似于传统的“生态系统工程师”(蚯蚓、蚂蚁和白蚁),许多马陆也有掘穴能力[95-96],在土壤团聚体的形成方面起着明显的促进作用[95,97]。Fujimaki等[95]在28天的培养实验中发现日本最常见的马陆Parafontarialaminata幼体能显著增加2 mm以上土壤团聚体的形成。巴西学者DaSilva等[97]的培养实验也发现热带常见的马陆Glyphiulusgranulatus能在短期内(28天)促进2.0—4.76 mm团聚体的形成。时雷雷等[98]的马陆培养实验也发现,大型马陆山蛩能在土壤中掘穴和活动(图4),在土壤中形成大型的孔隙,促进土壤的通气透水性能。Bowen和Hembree[96]详细地研究了美国两种山蛩类马陆在土壤构建巢穴的结构,这些结构非常复杂,能在很大程度上改变土壤的结构。山蛩类马陆在温带和热带地区广泛分布,并且数量巨大,因此,人为引进这些马陆,可能是生态恢复过程中土壤结构改良的一种重要途径。其二,马陆能大量取食凋落物,由于其同化效率低,大部分凋落物转化为粪球并成堆排放在凋落物和土壤表面,可促进土壤腐殖质的形成和改良表层土壤的物理、化学和生物性质。Frouz等[99]发现,在采矿废弃地恢复的中期,马陆等大型分解者动物的出现,其粪球的大量产生能在土壤表面形成一个发酵层(Fermentation layer)。这个发酵层有较高的生物活性,能改变土壤的化学和生物学特性,促进土壤的恢复。王梦茹等[98]在鼎湖山常绿阔叶林地表也观察到大量成堆的马陆粪球,其形态和周围土壤明显不同(图5);这种现象在热带和温带森林皆广泛存在 。

图4 蜷曲于土壤中的马陆(张洪芝拍摄)Fig.4 The curled-up millipede in soil (photo by Hongzhi Zhang)

图5 鼎湖山亚热带次生常绿阔叶林马陆及其排泄的粪球堆(张洪芝拍摄)Fig.5 The millipedes and their fecal pellets in the subtropical secondary evergreen broadleave forest in Dinghushan Mountains (photo by Hongzhi Zhang)
微生物对土壤结构的影响近年来也日益得到关注。Lehmann 等[86]对183篇文献进行了整合分析,结果认为数量占绝对优势的细菌和真菌对团聚体形成的贡献最大;其中细菌对大团聚体(Macroaggregates)和微团聚体(Microaggregates)的形成都有明显的促进作用,但真菌主要影响大团聚体的形成。该研究还发现个体小的土壤生物(宽度< 100 μm)对团聚体形成的贡献显著大于个体大的土壤生物(宽度> 100 μm);而且,非运动型细菌(Non-motile bacteria)对团聚体的形成的贡献显著大于运动型细菌(Motile bacteria)。
球囊菌门真菌及其与植物形成的菌根(Mycorrhiza)对土壤团聚体形成的积极作用最受关注。菌根是土壤真菌与陆地植物根系形成的一种互惠共生体[100],对植物生长常有明显的促进作用。早在3.5—4.5亿年前,菌根真菌就与古老的陆生植物形成了共生关系[101-102]。由于菌根真菌无法进行光合作用,必须从宿主植物中获取光合产物并转化为自己的生物量。据估测,植物光合作用固定的C有4%—26%转移至菌根真菌,由此固定的土壤有机C含量可达54—900 kg/hm2,占土壤总有机C的15%[103]。此外,球囊霉素土壤相关蛋白(Glomalin-related soil protein, GRSP)是丛枝菌根真菌产生并分泌到根外的糖蛋白,GRSP随着丛枝菌根真菌的衰亡和降解而释放到土壤中,最高可占土壤总C的5%[104],并在土壤中可维持6—42年;它们是土壤主要有机物质之一,同时也是维持土壤团粒结构的重要有机分子,故具有维持土壤结构和土壤肥力的功能[105]。同时,菌根真菌的菌索和外延菌丝在土壤中形成庞大的菌丝网,且其总长度远远超过宿主的根系总长度,扩大了宿主植物根系吸收面积和范围,因此菌根真菌能帮助植物从土壤中吸收磷以及其他矿物元素[106-107],并因此提高了植物耐盐能力[108]。
近年来,菌根真菌的土壤改良效应研究取得了重要的进展。Asmelash等 (2016) 综述了菌根真菌在退化土地恢复中的潜在作用:有大量的科学证据表明菌根真菌可以改善土壤质量,提高地上及地下生物多样性,提高水分和养分胁迫土壤中乔木和灌木幼苗的成活率、生长及定殖过程[109];今后,在进一步开展对菌根真菌的资源、功能、作用和调控机制等方面进行深入的研究的同时,需要加强开发可用于野外森林生态系统管理和恢复的廉价的菌剂。菌根真菌和土壤动物在土壤改良过程中的协同作用,也值得关注。
2.2.2土壤生物的肥力调节功能
土壤肥力(Soil fertility)是土壤支持健康生态系统生产力的综合能力,其内涵早已从最初的物理和化学肥力拓展至“生物-物理-化学”肥力,称之为“地力”更为恰当[110-112]。将土壤养分循环的理论知识应用于可持续农业的尝试由来已久[113]。作为主要的分解者及生物固氮的主体,土壤微生物无疑是土壤肥力的基础调节者之一[114-115];土壤动物则可以通过取食作用(Trophic effect)和非取食作用(Non-trophic effect)调控土壤微生物及其主导的生物化学过程[60]。显然,土壤生物对土壤肥力的调控过程并不局限于其对生物固氮的贡献。通过土壤生态学理论的不断完善,优化调控土壤食物网中土壤动物和微生物的互作过程的途径,进而保障养分供给和作物生产力,是可持续农业研究的重要方向[21]。1986年,为了应对传统耕作模式引起的地力衰退和环境污染等问题,荷兰科学家提出以综合农业体系(Integrated farming)代替传统农业体系,并正式启动了荷兰耕地农业系统土壤生态学项目(Dutch Programme on Soil Ecology of Arable Farming Systems)[116]。该项目开展的早期研究认为蚯蚓和原生动物的活动对微生物矿化有机质过程有关键的促进作用。但是,也有研究认为土壤微生物多样性在农业土壤可持续利用方面有重要的潜在作用[117]。应该注意,得益于其庞大的数量和快速的周转,细菌在有机质分解及与之相连的能量流动和养分循环过程中可能起着基础性的作用[118]。在Horseshoe Bend 农田生态系统中的研究也发现:在冬春季节,免耕和传统耕作农田中细菌的生物量和代谢活动都远高于真菌[119]。蚯蚓及其与其他土壤生物的相互作用可以通过调控土壤结构和有机质的分解过程而影响土壤肥力,相关的研究也相对充分,故可以从蚯蚓与土壤肥力的关系中大概把握土壤生物对土壤肥力之贡献。Lemtiri 等[120]认为可以将蚯蚓群落变化作为土壤肥力和质量的生物指示物,以评估土壤管理措施的效果。
蚯蚓对土壤有机质动态及养分循环的影响与耕作措施及蚯蚓-微生物-植物三者的互作关系紧密相关。其一,蚯蚓和植物的有效协作,可以促进生态系统对碳的固定。蚯蚓的活动可能促进植物生长[121-122]并将大量光合产物输入土壤[123];同时,植物的存在也往往有利于蚯蚓的生存,后者又可能进一步促进植物的生长,形成良性循环。其二,蚯蚓-植物-微生物的协作,可更加优化土壤碳和养分动态,促进农林生态系统的可持续发展。虽然土壤微生物可能与植物根系竞争氮,但是它们可能通过对氮的利用时间的分异而实现互利共赢,同时根系和土壤微生物的紧密合作,也减少了土壤氮的流失[124]。蚯蚓的存在促进了土壤无机氮的含量,并促进土壤微生物对氮的持留[125-126],客观上加强了上述植物-土壤微生物之间的互利关系。有意思的是,最近在广东鹤山的研究表明,环热带区的广布种蚯蚓南美岸蚓(P.corethrurus)的活动可明显促进土壤铵态氮的累积,且这些铵态氮无法直接被菌根真菌侵染率较低(< 30%)的植物(Dicranopterisdichotoma)吸收利用,但是接种菌根真菌(Rhizophagusintraradices)则可以帮助植物有效利用这些因蚯蚓活动而产生的铵态氮[127]。另外,蚯蚓还可能促进根瘤形成过程和固氮植物生长[128]及增加土壤磷的活性并缓解植物和土壤微生物对磷的潜在竞争[129]。如此一来,植物根系、土壤微生物、蚯蚓和菌根真菌等形成了相互协作的体系,可以显著减少养分的损失并提高其利用效率。反之,若是植物和土壤生物之间配合不好,则可能造成养分的固定或淋失[127,130-132],降低生态系统对养分的利用效率。其三,耕作措施对土壤生物在土壤碳和养分循环中的作用也有重要影响。例如,Fonte和Six[133]在南美开展的野外中宇宙实验发现,单纯添加不同质量的有机质对玉米田土壤性质和玉米生长影响很小,但是,添加蚯蚓(P.corethrurus)并辅以植物残体,可有效提高土壤大团聚体中微团聚体的有机质含量。而且,虽然蚯蚓可提高玉米对添加外源氮的吸收,但也可能加大土壤有效磷的淋失。可见,合理地进行蚯蚓及有机质管理,对提高热带农林生态系统的生态服务功能有重要的潜在价值。Fragoso等[121]也发现表栖类和深栖类蚯蚓的生存对地表凋落物有很强的依赖性,所以植物秸秆或动物粪肥等的合理回田,可促进农田中蚯蚓群落的恢复,进而提高土壤肥力和作物生产力。Coleman 等[54]估计在一公顷农田中,蚯蚓活动每年可以影响数吨的易分解的有机质;故将保护性耕作(以增加蚯蚓数量)和添加高质量的作物残体相结合,对农田生态系统健康可能会有事半功倍之效。
此外,蚯蚓堆肥技术(Vermicomposting)通过蚯蚓对“有机-无机”物质的大量取食,以及蚯蚓与肠道微生物的协同作用形成了一个宏大的“蚯蚓-有机质-微生物”有机融合的蚓触圈(Drilosphere),为高效地转化有机质并改善土壤结构创造了绝佳的条件。Chaoui等[134]发现,蚯蚓堆肥产生的蚓粪和蚯蚓堆肥产物可显著地提高小麦对磷和钾的吸收,并且与普通堆肥或化肥相比,产生盐胁迫的可能性更小。蚯蚓堆肥技术还可以明显促进其他微生物肥料的使用效果。Song等[135]在番茄-菠菜轮作体系中的研究发现:单独施植物根际促生菌(Plant growth-promoting rhizobacteria, PGPR)对土壤性质和作物生长并无明显影响,但是,蚯蚓堆肥存在时,根际促生菌可使土壤微生物生物量碳氮明显升高,并不同程度地提高番茄和菠菜的产量及其维生素C的含量。可见,若能充分发挥蚯蚓堆肥及其与有益微生物的协同作用,或可极大地减少化肥的使用。Lavelle等[136]很早就提出通过调控蚯蚓群落以保持土壤质量的思路。蚯蚓活动将地表凋落物转入土壤,并同时促进了有机质矿化和腐殖质化过程[136-137],进而深刻影响了土壤结构及土壤养分释放过程,为满足植物的养分需求奠定基础。但是,农田生态系统中蚯蚓群落面临多种物理和化学过程的胁迫,蚯蚓活动锐减。通过合适的管理措施恢复蚯蚓群落或寻找并引入耐受性强的蚯蚓种类,是发挥蚯蚓作用的关键。我们推测,若能将整个生态系统优化成一个蚯蚓堆肥系统,或许就建成了一个真正的可持续农业系统。
需要注意的是,土壤生物及土壤食物网对生态系统的资源总量、资源利用效率和生产力的影响错综复杂,以至于任何一个所谓的“具体”问题都包含了很多的子问题,导致短时间内难以找到有效的办法予以解决。例如,我们仍然需要大量的基础研究才可能知道如何通过管理有机质的输入及提高微生物活性等途径提高养分的可利用性[21]。在此,我们试着对土壤生物在碳和养分循环方面的“角色”及其相互关系进行梳理(图6),以期尽可能地把大问题分解成一个个可能被独立研究的具体问题。总的来说,土壤中碳和养分的稳定供给,是维持农林生态系统物质循环和生产力的关键。一方面,土壤生物可以调控可利用碳和养分“蛋糕”的大小。土壤动物和微生物可以通过影响光合作用效率或凋落物输入量及其分解过程等调控土壤可利用碳总量,进而影响土壤生物自身的群落大小及活性,最终影响生态系统总固氮量和有效磷总量等。另一方面,土壤生物可以影响可利用资源的“命运”。土壤微生物的快速生长可以迅速将碳和养分固持于微生物生物量中,避免意外的资源淋失;然后,微生物在各种生物或非生物“干扰”(如蚯蚓或线虫的取食或干湿交替等)的作用下,又可将原先固持的养分释放回土壤;这些养分或重新固持于微生物生物量中、或保存于各类团聚体中、或直接供植物吸收利用。最后,土壤食物网调控的“养分释放”与植物的“养分吸收”能否协调一致,将最终影响整个生态系统对养分的利用效率。如果可利用的碳氮磷浓度升高太快,植物和微生物来不及或因为某些原因无法及时利用,则将造成资源的 “闲置”或损失。我们认为,理想的可持续农林生态系统具有这样的特征:管理措施得当,能量和养分供给充足,生物固氮适时适量;可利用的碳和养分主要存在于某种缓存库中,譬如海绵里的水,适度活跃却不易流失,但植物或土壤生物需要的时候,总是能从海绵里“挤出”所需的资源。蚓粪的特性类似于这样的资源缓存库,而菌根真菌等则类似于植物赖以“挤出”养分所凭藉的武器。

图6 土壤生物对农业生态系统土壤肥力的调节作用概念图Fig.6 A conceptual framework showing how soil biota may regulate soil fertility in agricultural ecosystems
2.2.3土壤生物的病虫害防治功能
土壤食物网中复杂的生物互作关系为病虫害防治提供了重要基础。在农林生态系统中,通过合适的管理措施充分发挥本地或引入的病虫害的天敌进行有效的生物防治,很早就被关注[138]。生物防治可以减少农药的使用,减少食物中农药残留,是有机农业的重要内容[139]。土壤食物网中生物多样性极高,长期的进化过程可能使很多土壤动物和微生物具有独特的病虫害防御体系,这也是建立高效、经济的病虫害防治技术体系所必需的宝贵的生物资源。
Sánchez-Moreno 和 Ferris[140]以线虫群落结构为研究对象,发现土壤对病虫害的抑制能力与土壤食物网动态和农业管理措施是紧密相关的;自然生态系统具有更长的食物链和更复杂的土壤食物网,进而可以有效地抑制植物病原线虫的种群扩张;而干扰明显的农田生态系统则失去了这种优势。叶成龙等[62]也发现化肥、有机肥和秸秆配施可以增加食细菌线虫的丰度,抑制植食线虫垫刃属和螺旋属的繁殖。当然,从调控整个土壤食物网的角度去提升农林生态系统的抗病虫害能力,目前多停留在理论探讨或“知其然,不知其所有然”的阶段。但是,蚯蚓及蚯蚓堆肥、昆虫病原线虫及微生物农药等方面的基础研究较多,并且已有不同程度的成功应用。
蚯蚓及蚯蚓堆肥对病虫害的抑制作用原因复杂,有些内在机制并未完全理清。胡艳霞等[141]发现蚯蚓粪对黄瓜苗期土传病害有明显的抑制作用。Teng等[142]发现环热带区广布种南美岸蚓(P.corethrurus)有抑制香蕉血病的潜力。南美岸蚓的存在还可显著抑制对植物有害的大个体植食性线虫,但是,若有较多的外源氮输入时,蚯蚓对这些线虫的抑制作用消失了[143]。可见,蚯蚓对植食性线虫的影响过程还受全球变化因子的调控,其内在机制仍有待深入研究。蚯蚓活动还可通过调节植物激素平衡而促进植物抵御病害。Puga-Freitas等[144]发现蚓粪中含有植物生长素相关的信号分子,因为蚯蚓活动刺激了植物根际促生细菌合成植物生长素,激活了植物对病害的防御机制,进而促进了粳稻和黑麦草的生长。
相对来说,基于微生物的病虫害防治措施则是在揭示了某种抗病虫害的生物机制后,再开发出相应的产品,防治效果也更加直接。给植物接种某些土壤微生物,可以改变植物组织的化学组成,进而调控有害昆虫的种群动态。在相关基础研究取得突破时,可能实现为植物“定制”抗病虫害的微生物接种方案[145]。最著名的案例是1938年苏云金芽孢杆菌(Bacillusthuringiensis)被用作高效且环境友好的生物杀虫剂,如今已经发展出一个基于芽孢杆菌属的生物杀虫剂、生物杀真菌剂、生物肥料等的大家族,实现了商品化[146]。相应地,Bt 棉等农产品也取得了巨大了经济和生态效应。Bt棉在高效抗虫的同时,其本身未发现对土壤微生物及酶活性有明显不利影响[147],也未明显改变根际土壤线虫的群落特征[148];Bt玉米产生的Cry蛋白对蚯蚓、线虫、原生动物、真菌和细菌也没有明显的毒性[149-150]。Lu 等[151]分析了中国北方6个省区36个地点从1990到2010年间的Bt 棉生态系统及周围区域的害虫动态。结果表明:随着Bt棉的种植,广谱性杀虫剂用量减少,Bt 棉区的瓢虫、草蛉和蜘蛛等广谱性捕食者的数量明显增加,而蚜虫等害虫数量则明显下降。重要的是,该研究还发现这些捕食者一定程度上抑制了Bt 棉区附近的玉米、花生和大豆等的害虫,说明Bt作物可能带来额外的生态效益。另一个比较成功的案例,是利用线虫肠道共生菌对昆虫的毒性防治蝼蛄等草坡和作物害虫。昆虫病原线虫在植物保护中的应用得到了较多的重视,商业化的线虫制剂已成功用于抑制农业害虫[152-154]。另外,在设施农业中,“强还原土壤修复法”被成功用于应对作物连栽引起的严重的土传病害,巧妙地利用了薄膜覆盖及土壤微生物对易利用碳的持续代谢形成的高温厌氧环境来杀灭病原菌,并改善土壤结构和肥力[155]。
当然,生物防治的效果仍受限于多变的立地条件、相对不足的基础研究以及从业者参差不齐的管理和技术素养[156]。无独有偶,综合害虫管理法(Integrated pest management, IPM)自1960s 年代提出以来,已经是发达国家主要的作物保护策略,但是在发展中国家的应用仍极其有限[157]。需要将生物防治和综合害虫管理法等结合起来,才能切实提高农林生态系统病虫害的防治效果。此外,通过分析捕食者肠道微生物的DNA[158]或比较捕食者与潜在的被捕食者的中性脂肪酸(NLFAs)组成等,结合野外的生物种群调查数据,确定捕食者、被捕食者或其他可能的替代食物之间的相互关系,是制定病虫害综合防治方案的重要科学基础。
总之,土壤生物群落及土壤食物网在可持续农业模式中土壤结构改善、土壤肥力提高和化肥农药减施等方面大有可为。例如,探究蚯蚓等关键土壤生物的生态功能,可以促进生物肥料及生物土壤改良剂等的生产应用;探究土壤食物网内部及其与植物之间的互作机制,可以促进环境友好的高效生物农药的开发,推动生物防治的广泛应用;探究土壤食物网特征的变化规律,可以建立更完善的生态系统健康的生物指示体系。
3 难点与展望
目前,土壤食物网特征与生态系统服务之间的直接联系或作用机制并不清楚[159]。这是难以对农田系统进行精准管理以充分发挥土壤生物在可持续农业中的关键作用的重要原因。
3.1 难点
将土壤食物网结构与功能的基础理论与方法运用于可持续农业的宏大实践,不可能一蹴而就。因为方法论和技术的局限,很多重要关系或过程以及它们的相对贡献难以厘清,包括取食与被取食,资源现存量、周转速率与累积量,生物与非生物作用,直接与间接影响等[160]。具体来说,有以下几方面困难。
3.1.1所测非真实(量化不准)
土壤生态系统是众多连续的、交织的和随时空变化的过程的集合体,故很难将其分解为独立的过程并量化研究,因为任何的“分解”尝试都可能改变系统的本质属性,导致量化失真。虽然,同位素示踪技术在一定程度上弥补了还原论研究方法的缺陷,但“量化不准”依然是土壤生态学研究的重要难题。“量化不准”的原因大致有以下几种:其一,背景噪音太大。如土壤N2通量的测定,野外土壤总有机碳氮的短期变化等。其二,所研究过程为连续过程的一个短暂的过渡态。如 “根系碳”进入土壤即被微生物迅速利用转化,若脱离土壤进行收集测定,又显然与真实环境不符,所测值定有偏颇,导致“根系碳”的量一直未能准确测定;并且在同位素示踪实验中也难以确定“根系碳”的13C丰度,进而影响后续的“根系碳”分配的计算。其三,所测参数有明显的时空变化。例如往往在短期内发现蚯蚓的存在明显刺激CO2排放,但蚯蚓最终可能降低CO2释放的总量;若仅关注了蚯蚓前期的效应则必然以为蚯蚓不利于碳的固存[137]。其四,所测过程与其他过程相互依存,无法分开。例如蚯蚓肠道微生物与土壤微生物既有竞争又相互补充,导致两者对碳氮过程的贡献不好独立评估。
3.1.2“关系”非真实
土壤食物网各组分及其与外部环境因子的关系错综复杂,很多“关系”是多个过程的综合作用,根据相关分析或者结构方程模型等数学方法得到的土壤生物类群之间或土壤生物与其环境因子(如碳氮资源)的“关系”,有很大的不确定性。Shao等[161]运用结构方程模型分析八角林土壤食物网的特征,发现基于真菌的食物链相对完整,而基于细菌的食物链各成员间关系更加“松散”。这种“松散”可能说明食细菌线虫同时受到其他因子(如捕食者)的调控,或是细菌同时受其他因子(如碳氮资源)调控,故并不能直接理解为“关系”弱。例如,他们发现食细菌线虫数量与细菌生物量没有明显的关联,但这并不意味着两者没有关系,因为食细菌线虫必然要取食细菌。并且,虽然14C的示踪实验表明仅有约0.1%的微生物生物量碳进入线虫网络[162],但是,线虫通过刺激细菌群落进而对生态过程的间接影响仍不可忽视[163]。另外,很多研究都只有土壤生物各类群的即时数量或生物量数据,完全没有考虑生物的世代周转速率,结果必然低估世代周转速率较高的类群的生物量。若土壤生物的“关系”是基于对土壤生物在单位时间和空间内生物量的累积量的分析而得到,当更加真实。但目前对于多数土壤生物,特别是小型土壤动物和微生物来说,其准确的生物量累积量仍不得而知。
3.1.3“效应”非永恒
在较长的时间尺度(如植被演替)上,土壤食物网结构及其生态功能会发生明显的变化。随着植被和土壤的发育,土壤生物群落组成以及各类群的相对比例,甚至是代谢特征和繁殖策略都可能发生明显变化,是故脱离了生态系统所处的发育阶段而讨论土壤生物和食物网生态功能,意义不大。例如,固氮植物及其根瘤菌在森林演替早期主要的作用在于促进生态系统氮的累积,但在土壤氮含量较高的成熟森林中,它们的主要贡献可能转而利用氮促进植物对磷的获取[164]。
3.1.4缺乏有效的野外土壤生物群落特征的监测手段
土壤生物生活在土壤“黑箱”中,用常规的监测手段,无法了解土壤生物的动态。目前,对土壤生物群落的调查,依然主要依靠直接挖土手拣,或采集土壤然后用干湿漏斗法分类,或用陷阱法诱捕等传统方法。这些方法费时费力,对样地破坏大,采样效率不高,所以基本上无法做到长期的连续观测。这直接导致我们对多数土壤生物的生活史特征,种群周转速率等关键信息知之甚少,进而影响了前述各种“关系”估算的准确性。
3.1.5很难在生态系统尺度上量化土壤食物网的贡献
已有的研究以室内的微宇宙实验为主,且往往仅针对一个或少数几个土壤生物类群。即使是蚯蚓这样的大型土壤动物,在野外条件下控制其群落大小也困难重重,坚持多年的长期实验更是寥寥无几。但野外大田中植物、土壤和土壤生物都具有高度的时空异质性,室内实验极大地简化了土壤食物网成员及其与环境之间的互作关系,所以这些小尺度的简化实验所得的结果对野外只有参考意义,理论上并不能直接用于指导农业生产。当然,很多室内实验发现的作用过程和机理,在野外仍然在起作用,但其净效应则可能远不如室内。例如,微宇宙中培养蚯蚓,常常很明显地看到蚯蚓降低土壤微生物生物量,但是,在野外则不一定,因为微宇宙的空间一定,资源有限,蚯蚓对土壤微生物的影响被放大,而野外土壤异质性高,空间大,土壤微生物存在大量的“避难所”,故蚯蚓对土壤微生物的净效应可能明显削弱,但这也并不否定蚯蚓在野外依然在调控土壤微生物。
3.2 展望
土壤食物网和可持续农业研究是土壤生态学、生物学和农学等多学科交叉的前沿领域。自20世纪中叶开始,积累了丰富的研究成果,同时提出了更多的科学问题。大量新兴技术的出现,为解决这些科学问题提供了契机。
3.2.1从差异中找同一性规律
土壤生态学研究,除了需要努力融入重要的生物学、生态学基础理论,如能量守恒、中性理论、生态位理论、生态代谢和化学计量学理论等,还需要对土壤生态学自身的规律有总体的把握。
首先,需要了解植物-土壤系统中的主要“关系”。植物-土壤系统中的“关系”大体可归纳为五类[160]:1)某些组分为其他组分或过程提供基础条件,即起“平台”作用。只要“平台”还在,作用就能维持,其作用大小与构成平台的组分本身的生长状况关系不大。例如,华南地区桉树人工林地表长满了芒萁,并不像人们以为的那样因为竞争养分而不利于桉树的生长,反而促进了整个系统的养分循环,为土壤食物网营造了一个良好的生境[165-166],但是,这种利好作用可能与芒萁本身的生长情况无关。2)某些组分对其他组分或过程有明确的正效应或负效应。例如,土壤肥力越高,土壤微生物生物量越大;但周转越慢。3)某些组分通过一种途径抑制其他组分或过程,又通过另外的途径促进该组分或过程。例如,蚯蚓取食过程可能降低土壤微生物生物量,但蚯蚓的代谢活动又可以为土壤微生物提供可利用的碳源,进而刺激土壤微生物的生长[137]。4)某些组分可明显影响其他组分或过程,但这种作用很难被直接观测。由多过程控制的组分的变化(如前述的食细菌线虫与细菌的关系)常常如此,特别是在野外条件下。5)物质和能量在某些组分内部循环。例如,土壤细菌获得易分解碳氮资源后,可能通过生长-死亡-再生长过程将大部分资源限定在细菌群落或少数紧密相关的类群(如病毒)的内循环中。了解上述主要的“关系”,可以对土壤生态学过程有总体的把握。
其次,需要了解重要生物或过程的基本量度。从基本的生物学规律出发,提出问题、设计实验、解读数据,可以事半功倍。例如,了解细菌是单细胞原核生物可以帮助理解细菌为何有“响应快、不稳定、对资源的利用能力和效率相对低”等特点;相反,真菌是多细胞真核生物,则有“稳定性强、对资源的利用能力和效率较高”等特点。最近有报道认为,自然生态系统中糖类(单糖双糖)占土壤有机质的比例约为10%[167]。这个比例可能是生态系统健康的一个关键表征。糖类活性高,是生态系统即时可利用能量的量度,所以糖类在总有机质中的比例过高过低都不适宜。土壤中留存的糖类相当于“货币”,比例过高,可能说明生态系统活力不足,周转过慢,类似于“经济过冷”;相反,土壤中留存的糖类比例过低,可能说明生态系统代谢太快,消耗过大,可能难以持久,类似于“经济过热”。土壤生物及土壤食物网对糖类代谢的调控作用,应该是未来的一个重要研究方向。另外,真菌细菌生物量比也被认为与有机质分解和养分循环等重要生态学过程紧密关联[168]。类似的重要量度还有:土壤微生物生物量碳氮磷占土壤总碳、总氮和总磷的比例;主要土壤生物类群的C∶N 比、N∶P比和C∶N∶P比等;细菌、真菌和主要土壤动物类群的生物量周转速率;微生物残体和代谢产物对土壤有机质的贡献百分比;根系来源碳持留于土壤的比例;凋落物来源碳对土壤有机质的贡献比例;生态系统中“生物固氮”占氮来源的比例;主要土壤生物类群的资源利用效率(生物量与呼吸量比);蚯蚓粪年产量;土壤孔隙度年变化及土壤生物的贡献;随着植被演替,纤维素、木质素及植物次生代谢物含量的比例的变化;捕杂食者/食微者比例及捕杂食者/植食者比例等随纬度或植被演替的变化等等。以上列举的很多量度,目前并不清楚,也正是大有可为的地方。
最后,需要了解重要的生物和非生物标记物。若特定的生物类群或过程,可形成或积累特定的生物和非生物的物质,正是某种同一性规律的集中体现。土壤生态学研究受困于土壤“黑箱”,发现并运用生物和非生物标记物以巧妙地揭示土壤食物网中的物质循环和能量传递过程,尤为重要。目前已经广泛运用的标记物有:生物细胞膜的磷脂脂肪酸(PLFAs),可以大致量化活体土壤微生物群落结构及主要类群的生物量[169];中性脂肪酸(NLFAs),作为细胞的储藏物质可以帮助分析土壤生物的食物来源[170];氨基糖,作为微生物残体的重要组分可帮助区分真菌和细菌在碳氮转化过程中的贡献[171];球囊霉素等菌根真菌的分泌物可促进团聚体形成,是菌根真菌影响土壤碳持留的重要途径[105]。每一个新的标记物的发现,都可能极大地推动整个土壤生态学的发展。
3.2.2发展和完善关键的实验和分析手段
土壤生态学的很多重要科学问题缺乏真正有效的研究手段,这也正是整个学科发展的瓶颈所在。首先,需要优化技术和试验方案,准确量化土壤生物群落主要特征。其一、土壤生物的食性。传统的肠道内容物检测法,只有大型土壤动物才方便实施,而且只能确定哪些东西被土壤动物摄入肠道,但不能确定是否为其食物;另外,要判断肠道内容物的成分,需要很长时间的经验积累,所以适用面很窄。测定土壤生物的13C和15N自然丰度,并与其可能的食物的13C和15N丰度做比较,则可以方便地确定土壤生物的食性范围(如植食者、枯食或腐食者、捕食/杂食者等等)。但是,要明确区分土壤生物具体的食物来源,则需要借助于13C和/或15N示踪实验,并比较土壤生物及其可能食物的中性脂肪酸(NLFAs)组成和含量。其二、土壤生物数量生物量年累积量。因为工作量大,对样地的破坏也大,采样不能太频繁,所以很难准确了解土壤食物网的时空动态。更重要的是,土壤生物即时数量和生物量并不能很好地反映整个土壤食物网构成。只有结合土壤生物主要类群在野外条件下种群的周转速率,计算出它们各自的数量和生物量年累积量,才能准确描述食物网各节点的能量和物质流动,进而评估土壤食物网的整体生态功能[118]。其三、土壤生物生活史和“物候”等的实时长期监测。了解主要土壤生物的生活史、繁殖策略、体型大小等关键特征及其时空变化规律,是从深层次研究土壤食物网内部组分及其与外部环境之间互作机制的关键。例如,蚯蚓的肠道过程及其粪便对蚯蚓的生态功能至关重要,不了解各生态类型蚯蚓的粪便的年产量、在土壤中的分布、碳氮磷含量及其稳定性等,就无法确切评估蚯蚓在生态系统中的贡献。
其次,需要优化技术和试验方案,准确量化土壤生物对碳和养分循环的贡献。土壤动物对凋落物和土壤有机质(包括其中的原生动物和微生物)的取食过程,往往可以加快有机质的周转,同时释放出养分。养分释放的多少,大致可以由土壤动物、微生物、凋落物和土壤有机质等的碳氮磷比的差异计算而得。但是,非取食过程对养分循环的贡献不能直接算出,只能通过特定的实验处理来间接评估。后者又受制于土壤生物群落结构的调控手段。考虑将微生物磷脂脂肪酸、氨基糖和宏基因组分析与同位素示踪技术结合(PLFA-SIP,氨基糖-SIP和DNA-SIP等)揭示与碳氮循环过程相关的土壤生物及其相对贡献。
最后,需要优化野外土壤生物群落结构调控技术。虽然化学驱离法在室内微宇宙实验中较好地控制了土壤生物群落结构[172],但野外的效果常不能持久,而且对环境不友好。目前,比较成功的案例是应用物理驱除/隔离的办法控制部分大型土壤动物类群。例如,电击法驱离蚯蚓[173]并配合魔术贴防止蚯蚓横向迁徙[174];网罩法控制蜘蛛[175];曲面隔离法控制蚂蚁[176]等。另外,利用不同孔径的网袋可以一定程度上量化不同大小的土壤动物对凋落物分解的贡献,但网袋改变了水热条件并可能引起土壤生物活性的变化。
3.2.3贯彻整体观的研究方法论
首先,需要在植被和土壤发育动态中探究土壤食物网特征及其生态功能。揭示土壤食物网及其对生态系统的效应在不同时空尺度上的变化规律,是全面理解土壤食物网生态功能的前提。土壤食物网与植被及土壤发育可能互为驱动力。已有的研究,集中在对处于特定演替阶段的生态系统内部组分的转化过程以及不同纬度带生态系统中土壤生物组成和数量特征等方面,而且多数并未基于整个食物网的变化开展工作[177]。
其次,需要在流域或区域的尺度上探究土壤生物与可持续农业的关系。以有机农业为代表的可持续农业与传统农业的一个根本区别,在于前者强调以系统的观点审视农业生态系统,并以此来指导制定土地管理方案[159]。传统的生态系统或景观尺度上的研究,把焦点局限在了农场内部,忽视了宏观上的通盘考虑。在流域尺度上进行可持续发展的生态规划和景观设计,可以更加有效地实施土壤生物资源的保护和综合性利用、作物的病虫害综合防治、农业废弃物综合利用及土壤和水体污染的综合治理等。
最后,需要切实发挥多学科交叉研究的优势。一方面、可持续农业必须以多学科的深入系统的基础研究为基石。农业生态系统包含植物-土壤互作、土壤食物网内部的互作、生物-非生物因子互作,以及管理措施与土壤食物网的互作等复杂关系。可持续农业须充分考虑农业系统中的生态学和生物地球化学过程,以最大限度地发挥系统各组分间的协同作用,降低对外部资源(如化学杀虫剂、化肥等)输入的依赖,进而在充分利用资源的同时仍维持植被和土壤系统的健康。另一方面、可持续农业涉及技术、经济和社会等各方面的因素,故其天然需要以整体观的方法论为指导。例如,长期的有机农业可提高土壤质量,生物多样性,改善水质等,但这些贡献常常未能在有机农产品的产量和品质及其经济效益上得到充分体现。由于没有建立完善的生态系统服务有偿使用机制,有机农业从业者的上述贡献没有得到应有的回报,限制了积极性[159]。所以,基于技术层面的考量提出的农业经营管理模式,还需要兼顾其产生的经济和社会效益。
3.2.4以有机质管理为核心的“土壤生物-可持续农业”发展思路
土壤有机质是人类保障粮食安全和应对全球变化的基础性资源[178],是农业生态系统可持续发展的核心[159]。Coleman等[54]反复强调了土壤有机质对于可持续农业系统的重要性。他们认为土壤碳动态和土壤生物多样性是关乎农业生态系统可持续发展的两大关键因子;但是,它们之间有何重要联系?受什么因子调控?仍不得而知。Magdoff和Weil[22]也在其专著SoilOrganicMatterinSustainableAgriculture中强调了土壤有机质在可持续农业中的关键作用。该书对有机质管理对土传病害的控制,土壤和作物管理对土壤微生物的影响,有机质、蚯蚓和微生物互作在促进植物生长中的贡献等做了较深入的阐述。Kennedy等[179]认为,虽然适用于可持续农业系统的微生物管理技术尚未发展成熟,但有两个策略需要贯彻。其一,运用适宜的管理措施以提高土壤有机质。其二、通过作物轮作、免耕或放牧管理等保持植物群落的多样性,促进根系生长,减少表土淋失等。考虑到凋落物质量可能是影响凋落物分解的最重要的因子[180],可针对不同土壤类型或不同的作物特性,施用以不同作物残体为基质的蚯蚓堆肥,实现有机质的针对性管理,发挥其对土壤结构、土传病害及土壤养分动态等的积极作用。另外,可持续农业实践可能并非要建立一个持续稳定的农业生态系统,而应该以生产力的最大化和养分损失的最小化为目标。通过优化秸秆还田方式及减少土壤干扰等措施,恢复自然生态系统中真菌及细菌通道中各生物间的平衡,可能是实现上述可持续发展目标的重要基础[57]。
4 小结
可持续农业与土壤过程的关键联系在土壤有机质,土壤结构和土壤食物网。土壤生物和有机质的互作过程可以改变土壤结构、水分和养分动态,后者又反过来影响了土壤食物网结构和有机质动态,进而调控植物对光、CO2、水和养分的利用,以及植物的病虫害状况,最终影响生态系统生产力和作物产量及品质。土壤有机质管理是调控土壤结构、土壤食物网及生态系统生产力的重要支点,但也是不确定性的重要来源。土壤有机质的矿化和累积分别与短期的农田生产力及生产力的可持续性密切相关。找到土壤有机质累积和矿化的平衡点[113],揭示土壤有机质、土壤结构和土壤食物网特征的内在联系,权衡农田生态系统的短期和长期效益[13],才能确定适宜的耕作模式和有机质管理制度。通过改变耕作措施和养分管理方式可提高土壤有机质含量[181],但直接通过调控土壤食物网以促进农业可持续经营的工作十分缺乏[182]。今后,需因地制宜地细化免耕、秸秆还田,及轮作等调控措施的方案,同时以土壤生物群落特征(如线虫群落结构)反映土壤食物网动态,找到根际和腐屑食物网中土壤生物主要类群变化的驱动力,并逐步厘清食物网内部各组分及驱动力间复杂的互作关系,为最终实现有针对性的土壤食物网管理奠定基础。当然,基于系统的面上调查及农田土壤生物群落野外长期控制实验研究,揭示农业经营管理措施、土壤食物网特征及生态系统生产力三者的内在联系,是充分发挥土壤生物在可持续农业中的积极作用的关键。脂肪酸图谱、氨基糖和DNA分析等与稳定同位素技术的有效结合,为揭示土壤食物网的生态功能提供了有力的技术支撑。
致谢:感谢河南大学环境与规划学院张洪芝博士提供部分马陆照片。
猜你喜欢
红领巾·探索(2021年2期)2021-08-26
学生天地(2020年34期)2020-06-09
当代陕西(2019年11期)2019-06-24
作文小学中年级(2018年5期)2018-06-11
快乐语文(2017年9期)2017-06-15
作文通讯·初中版(2017年1期)2017-04-13
作文通讯·高中版(2017年12期)2017-02-06
爆笑show(2016年2期)2016-04-11
少儿科学周刊·儿童版(2015年6期)2015-11-24
爆笑show(2014年7期)2014-09-03
