
5-氨基乙酰丙酸和乙烯利对东北春玉米源库碳平衡的调控效应
2020-07-02 09:47:48李瑞杰唐会会王庆燕许艳丽霖闫董志强张凤路
作物学报 2020年7期
李瑞杰 唐会会 王庆燕 许艳丽 王 琦 卢 霖闫 鹏 董志强,* 张凤路
1,# 河北农业大学农学院 / 省部共建华北作物改良与调控国家重点实验室 / 河北省作物生长调控实验室, 河北保定 071001; 2,# 中国农业科学院作物科学研究所 / 农业农村部作物生理生态与栽培重点开放实验室, 北京 100081
东北春玉米区是我国重要的玉米主产区之一,播种面积占全国的30%左右[1]。该区域耕地以黑土为主, 土壤肥力高, 是我国玉米单产最高的区域之一, 然而由于生育期内环境实际平均气温低于玉米适宜生长温度, 且初霜期早, 低温冷害频发, 进一步限制了东北春玉米单产的提高[2]。有研究表明, 低温能够造成作物生育期延长, 源-库代谢失调, 光合产物及淀粉积累量降低, 花粉败育率提高, 干物质和产量显著降低[3]。此外低温还可以加速玉米叶片衰老, 降低籽粒的灌浆速率, 降低产量[4]。而光合产物主要以蔗糖的形式通过韧皮部输送到库端, 与蔗糖代谢的密切相关酶主要包括蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)和转化酶[5], 低温造成蔗糖代谢相关酶的活性降低, 淀粉积累减缓, 籽粒灌浆速率下降,千粒重降低, 产量降低[6]。因此, 增强低温冷害条件下玉米功能叶与籽粒间蔗糖代谢运输能力、促进灌浆对缓解东北春玉米低温冷害胁迫, 提高玉米单产水平具有重要意义。
5-氨基乙酰丙酸(5-ALA)作为叶绿素合成的关键前体物质, 可以促进叶绿素合成, 增强叶片的光合作用, 促进干物质积累, 提高产量, 在低温[7]、弱光[8]、盐碱[9]等逆境条件下, 表现尤为明显。外源5-ALA 处理可以提高甜瓜幼苗可溶性糖的含量, 增强植株抗冷性和耐弱光性[10]。5-ALA 可以提高低温胁迫下辣椒叶片中脯氨酸和可溶性糖含量, 显著提高植株生长量[11], 叶面喷施25 mg L-15-ALA 可以诱导葡萄中蔗糖转化酶基因的表达, 促进蔗糖代谢,增加可溶性固形物含量, 改善果实品质[12]。喷施含有5-ALA 的植物多糖类复合制剂, 能明显提高小麦旗叶蔗糖含量, 促进养分向籽粒的转移, 提高吐丝后干物质对产量的贡献率[13]。本课题前期研究表明5-ALA 处理能够改善玉米的群体结构, 提高灌浆后期LAI, 延缓叶片衰老[14], 然而5-ALA 的化学性质不稳定, 在高温、碱性和光照等环境下易分解, 单剂使用在大田中作用效果并不稳定。乙烯利(ETH)在大田生产中常作为一种生长延缓剂广泛使用, 能够增强绿豆芽下胚轴蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)和酸性转化酶(AI)活性, 促进蔗糖向下胚轴的转运和分解[15], 还能够增强橡胶树叶片中蔗糖代谢酶的活性, 提高胶乳产量[16]。乙烯利在pH<3.5 的溶液中化学性质稳定, 能为5-ALA 提供稳定的酸性环境, 并形成稳定的复合物, 因此将5-ALA 与ETH复配可能在调控大田玉米源-库碳代谢方面具有重要的应用价值, 然而相关的研究鲜有报道。因此,本研究探讨不同5-ALA 和ETH 复配剂组合对东北春玉米源-库碳代谢及其产量的调控效应, 以期为解决东北春玉米因低温冷害造成源库不协调而导致的减产问题提供一种新的技术途径, 保障该区域春玉米高产稳产。
1 材料与方法
1.1 试验地概况
中国农业科学院吉林省公主岭试验站(43°29'55″N, 124°48'43″E)年平均降雨量594.8 mm,有效积温≥10℃) 2600~2800℃, 试验田土壤为黑钙土, 0~20 cm 耕层土壤含有机质26.7 g kg-1、全氮1.4 g kg-1、 速 效 氮 155.3 mg kg-1、 速 效 磷34.4 mg kg-1、速效钾184.2 mg kg-1, pH 5.8。2018、2019年玉米生长季气温和降雨量如图1。
1.2 试验材料与设计
试验以中单909 为材料, 将5-氨基乙酰丙酸(5-ALA)(设置0、11.25、22.50 和33.75 g hm-24 个浓度梯度)和乙烯利(ETH)(设置0、450 和900 mL hm-23 个浓度梯度)复配, 共计12 个处理(表1), 在拔节期(V6)叶面混合喷施处理, 对照区喷施等量清水。试验小区均为8 行区, 行距为60 cm, 小区长12 m,小区面积为57.6 m2, 采用随机区组试验设计, 3 次重复, 基施尿素412.5 kg hm-2、磷酸二铵225 kg hm-2、氯化钾132.4 kg hm-2, 施肥量N∶P2O5∶K2O = 230.5∶103.3∶79.5 (kg hm-2), 其他田间管理同当地大田生产。2018年4月29日播种, 10月1日收获, 2019年5月4日播种, 10月3日收获。
图1 2018年和2019年公主岭试验站春玉米生长季日累计降雨量(柱形)和日平均气温(折线)Fig. 1 Daily total rainfall distribution (bar) and daily mean air temperature (line) during spring maize growth stage at Gongzhuling Experimental Station in 2018 and 2019
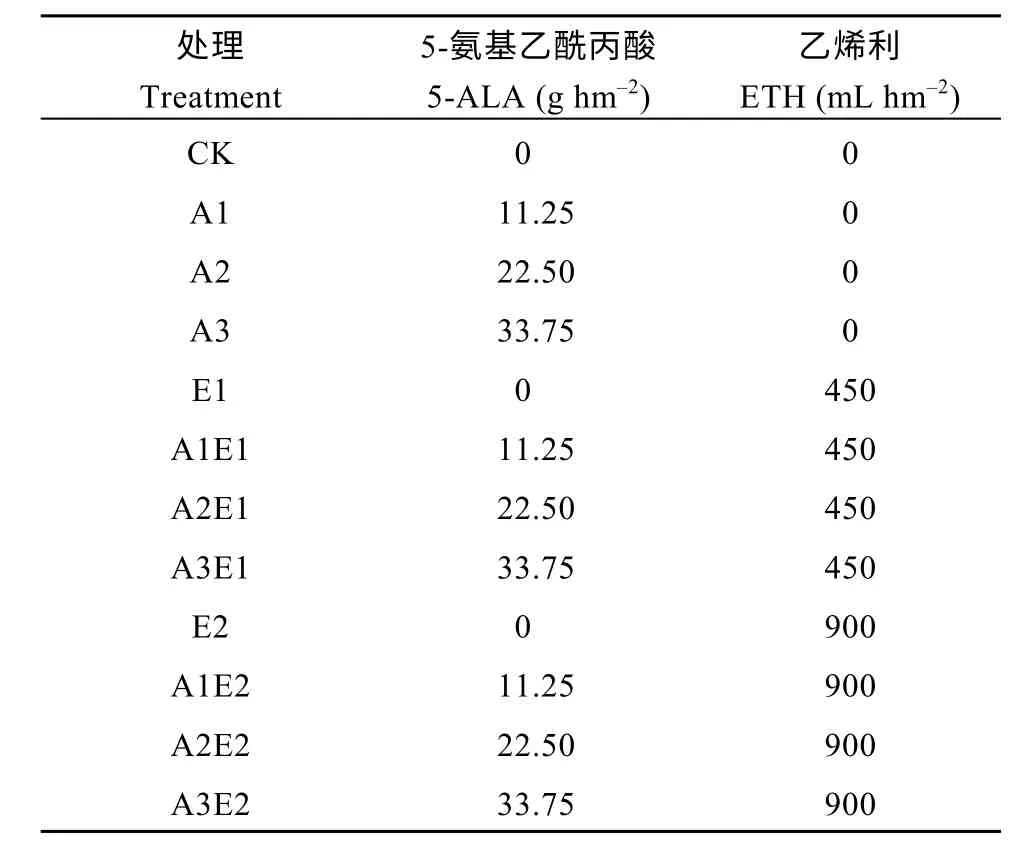
表1 2018年和2019年不同试验处理下5-氨基乙酰丙酸和乙烯利使用量Table 1 Amount of 5-aminolevulinic and ethephon used under different treatments in 2018 and 2019
1.3 测定的项目及方法
1.3.1 产量及产量构成因素 玉米成熟后, 取小区中部10 m2收获测产, 选取代表性果穗20 个, 调查穗部性状(穗长、秃尖长、穗粗、穗粒数和千粒重),测定出籽率和含水率, 并按14%质量含水量折算玉米单产。
1.3.2 籽粒灌浆速率模拟 在玉米吐丝期, 选取长势、穗型基本一致的植株挂牌标记。自吐丝后10 d起, 每隔10 d 在每小区标记的植株上取3 个果穗,每穗取中部籽粒100 粒, 于105℃烘箱中杀青30 min,80℃烘至恒重后称重。参照陈传永等[17]方法, 以吐丝后天数(x)为自变量, 每隔10 d 测得的百粒重(y)为因变量, 用Logistic 方程y=a/(1+be−cx) 模拟籽粒灌浆过程, 其中a为终极生长量,b为初值参数,c为生长速率参数, 灌浆速率最大时的生长量Wmax=a/2,达到最大灌浆速率的天数Tmax= (lnb)/c, 最大灌浆速率Gmax= (c×Wmax)/2, 活跃灌浆天数P=6/c。
1.3.3 蔗糖与蔗糖代谢关键酶活性 分别在9 展叶期(V9)、大喇叭口期(V12)、吐丝期(VT)、吐丝后10 d (VT+10)、吐丝后20 d (VT+20)、吐丝后30 d(VT+30)、吐丝后40 d (VT+40), 取玉米最新展开叶(吐丝前)和穗位叶(吐丝后)于液氮中保存。参照李光彦等[18]的方法测定功能叶蔗糖含量、蔗糖磷酸合酶(SPS)活性和蔗糖合酶(SS)合成方向酶活性。参照赵智中等[19]的方法并稍加改进, 将样品研磨后加入内含0.05 mol L-1蔗糖的pH 4.7(AI)/pH 7.0 (NI) 0.08 mol L-1的乙酸-K3PO4缓冲液和酶提取液, 在30℃下水浴反应10 min, 沸水浴5 min 终止反应。用高温杀死的酶液做对照。冷却, 加1 mL 3,5-二硝基水杨酸试剂, 沸水浴5 min, 冷却至室温在540 nm 波长下比色。以每克植物鲜样在1 h 内反应生成的蔗糖含量(mg)表示蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)活性, 以每克鲜样在1 h 内反应生成的果糖含量(mg)表示酸性转化酶(AI)、中性转化酶(NI)和蔗糖合酶(SS)分解方向活性。
1.4 试验数据处理
采用Microsoft Excel 2016 整理数据和制作图表,SPSS 18.0 软件统计分析, 以LSD (P<0.05)检验平均数间的差异显著性。
2 结果与分析
2.1 5-ALA-ETH 复配剂处理对玉米功能叶蔗糖合成的影响
2.1.1 5-ALA-ETH 对玉米功能叶蔗糖含量的影响
如图2 所示, 单施5-ALA 处理可以提高生育期内功能叶的蔗糖含量, 其中A1 处理增幅最大, 在VT+10 至VT+40 比对照平均增加26.6%, 而A2 处理在V9 至V12 平均增加15.3%。单施ETH 处理降低了蔗糖含量, 其中 E1 处理平均降低 20.2%。5-ALA-ETH 复配剂处理中, A2E1 处理提高了V9 至V12 和VT+30 至VT+40 叶片蔗糖含量, 比对照平均分别提高5.0%和14.3%, 降低了VT 至VT+20 叶片的蔗糖含量, 比对照平均降低14.4%, 而A2E2 处理生育期内比对照平均降低25.6%。
2.1.2 5-ALA-ETH 对功能叶蔗糖合酶(SS, 合成方向)活性的影响 如图3 所示,单施5-ALA 处理降低了生育期蔗糖合酶SS 的活性,其中A2 和A3 处理VT 至VT+10 平均比对照分别降低67.8%和67.7%。单施ETH 处理降低了生育期内功能叶SS 活性, 其中E1 和E2 处理VT 至VT+10 分别降低21.7%和40.7%。5-ALA-ETH 复配剂处理降低了生育期内SS 活性, 其中A1E1 和A2E1 处理比对照平均分别降低21.8%和26.1%, 而A1E1 处理在V9 至V12 平均提高19.3%,而A2E1 处理在V9 提高73.4%, A2E2 处理平均降低3.9%, 而VT+10 至VT+20 平均提高47.8%。
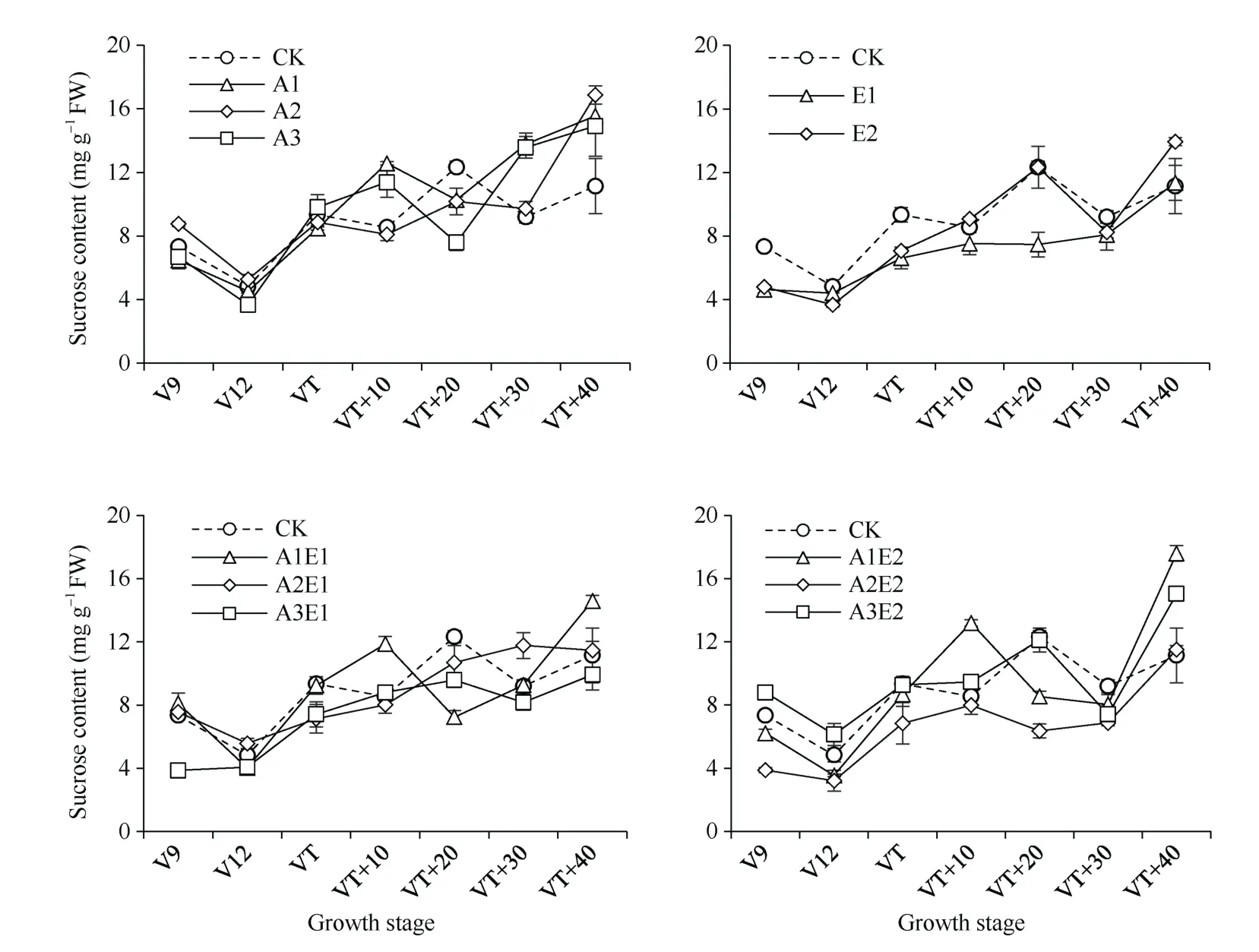
图2 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对玉米功能叶蔗糖含量的影响Fig. 2 Effects of 5-ALA, ETH, and 5-ALA-ETH on sucrose content in functional leaves of maize in 2018

图3 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对蔗糖合酶(SS, 合成方向)活性的影响Fig. 3 Effects of 5-ALA, ETH, and 5-ALA-ETH on the activity of sucrose synthase (SS, synthetic direction) in 2018
2.1.3 5-ALA-ETH 对功能叶蔗糖磷酸合酶(SPS)活性的影响 如图4 所示, 单施5-ALA 处理提高了生育期内SPS 活性, 其中A1 和A2 处理吐丝后比对照平均分别提高65.3%和80.3%。单施ETH 处理提高了生育期内SPS 活性, 其中E1 和E2 处理吐丝后分别提高12.2%和48.3%。5-ALA-ETH 复配剂处理均提高了吐丝后叶片SPS 活性, 其中A1E1、A2E1、A3E1、A1E2、A2E2 和A3E2 处理在VT+10 至VT+40比对照平均分别提高73.2%、12.2%、30.8%、28.2%、39.3%和48.5%, 且A2E1 处理在V9 至V12 比对照平均提高9.9%, 而A1E1 和A3E1 处理分别降低4.9%和3.3%。

(图4)
图4 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对蔗糖磷酸合酶(SPS)活性的影响Fig. 4 Effects of 5-ALA, ETH, and 5-ALA-ETH on the activity of sucrose phosphate synthase (SPS) in 2018
2.2 5-ALA-ETH 复配剂处理对籽粒蔗糖分解的影响
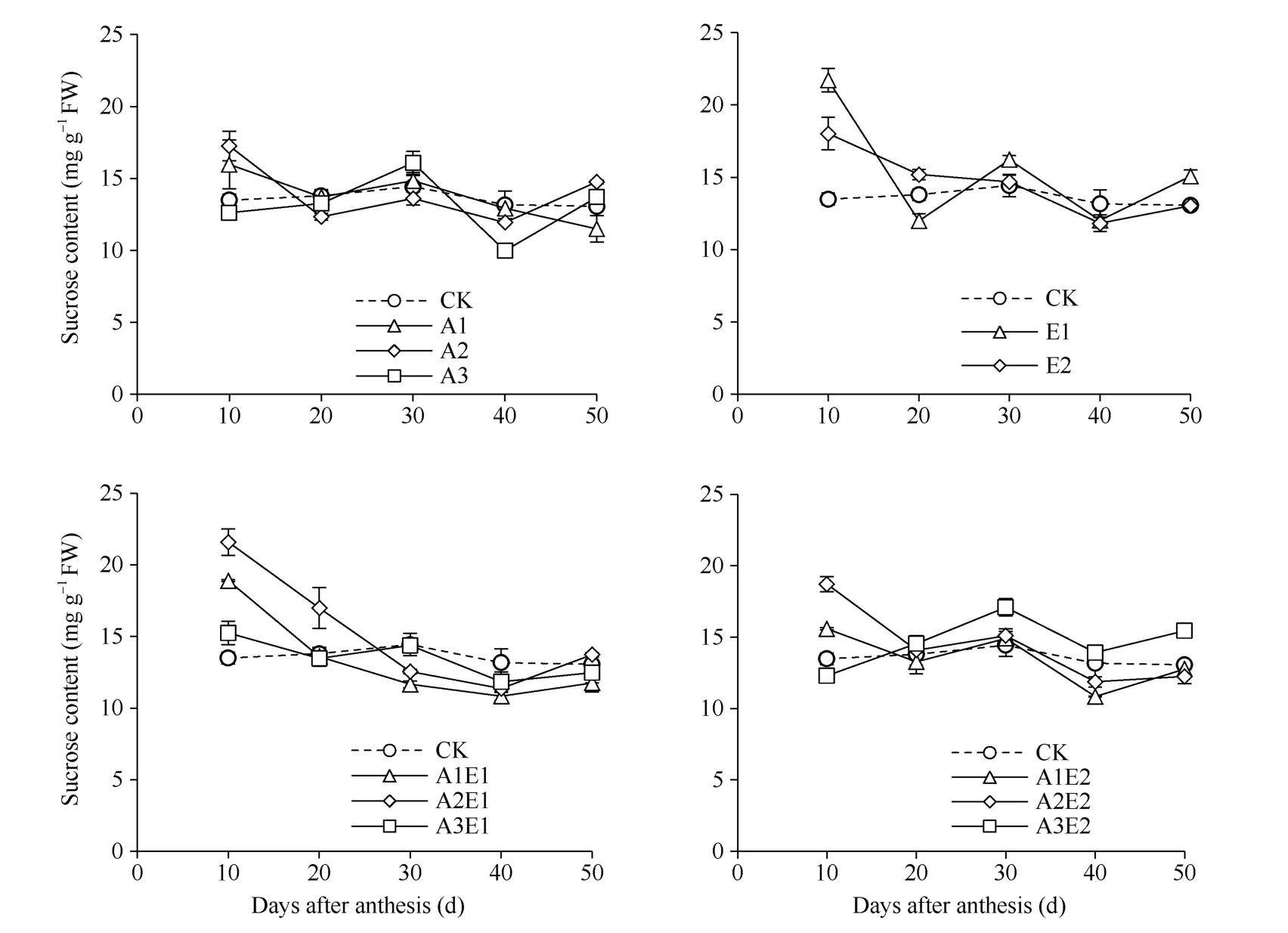
图5 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对籽粒蔗糖含量的影响Fig. 5 Effects of 5-ALA, ETH, and 5-ALA-ETH on sucrose content in grains in 2018
2.2.1 5-ALA-ETH 对籽粒蔗糖含量的影响 如图5 所示, 单施5-ALA 处理可以提高灌浆前期籽粒中蔗糖含量, 其中A1 和A2 处理灌浆10 d 比对照分别提高18.4%和27.9%。单施ETH 处理可以提高灌浆前期籽粒中蔗糖含量, 其中E1 处理灌浆10 d 至20 d 平均提高23.5%。5-ALA-ETH 复配剂处理可以提高灌浆前期籽粒中蔗糖含量, 其中A2E1 处理在灌浆10 d 至20 d 比对照平均提高41.4%, 30 d 至50 d平均降低16.0%, 而A3E2 处理在灌浆20 d 至50 d比对照平均提高12.0%, 而灌浆10 d 平均降低9.0%。
2.2.2 5-ALA-ETH 对籽粒蔗糖合酶(SS)活性的影响
如图6 所示, 单施5-ALA 处理提高了灌浆前期蔗糖合酶SS (分解方向)活性, 其中A2 和A3 处理灌浆10 d 至20 d 比对照平均分别提高18.7%和41.0%,而A2 处理30 d 至50 d 降低8.2%。单施ETH 处理, 相比对照平均提高17.2%, 其中E1 处理平均提高33.6%,灌浆10 d 至30 d 平均提高35.4%。5-ALA-ETH 复配剂处理提高了灌浆前期SS 分解方向活性, 其中A2E1和A2E2 处理在灌浆10 d 至20 d 比对照平均分别提高12.5%和14.4%, A3E2 处理灌浆20 d 提高65.5%, 而A1E1 处理灌浆20~30 d 降低44.5%。
2.2.3 5-ALA-ETH 对籽粒酸性转化酶(AI)活性的影响 如图7 所示, 单施5-ALA 处理降低了酸性转化酶(AI)活性, 其中A1 和A3 处理比对照平均分别降低27.6%和18.9%, 而A2 处理提高18.1%。单施ETH 处理, E1 和E2 处理比对照平均分别提高46.5%和16.0%。5-ALA-ETH 复配剂处理提高了灌浆前期酸性转化酶(AI)活性, 其中A1E1 和A3E1 处理灌浆10 d 至30 d 比对照平均分别提高54.2%和34.9%,A2E1 和A3E2 处理灌浆10 d 分别提高52.8%和34.9%, 而A1E2 处理在灌浆10 d 至20 d 平均降低16.7%。
2.2.4 5-ALA-ETH 对籽粒中性转化酶(NI)活性的影响 如图8 所示, 单施5-ALA 处理降低了中性转化酶(NI)活性, 其中A1、A2 和A3 处理比对照平均分别降低38.3%、21.9%和48.5%。单施ETH 处理, E1和 E2 处理平均分别降低 20.0%和 15.0%。5-ALA-ETH 复配剂处理中, A1E1、A2E1、A3E1 和A2E2 处理相比对照平均分别提高14.9%、7.0%、13.4%和12.1%, 其中A3E1 处理在灌浆10 d 至30 d提高16.0%, A1E1 和A2E2 处理灌浆20 d 至50 d 平均分别提高38.0%和32.0%, A2E1 处理灌浆10 d 提高24.1%, 而A1E2 和A3E2 处理在灌浆10 d 分别降低68.9%和63.1%。
2.3 5-ALA-ETH 复配剂处理对籽粒灌浆速率模拟的影响
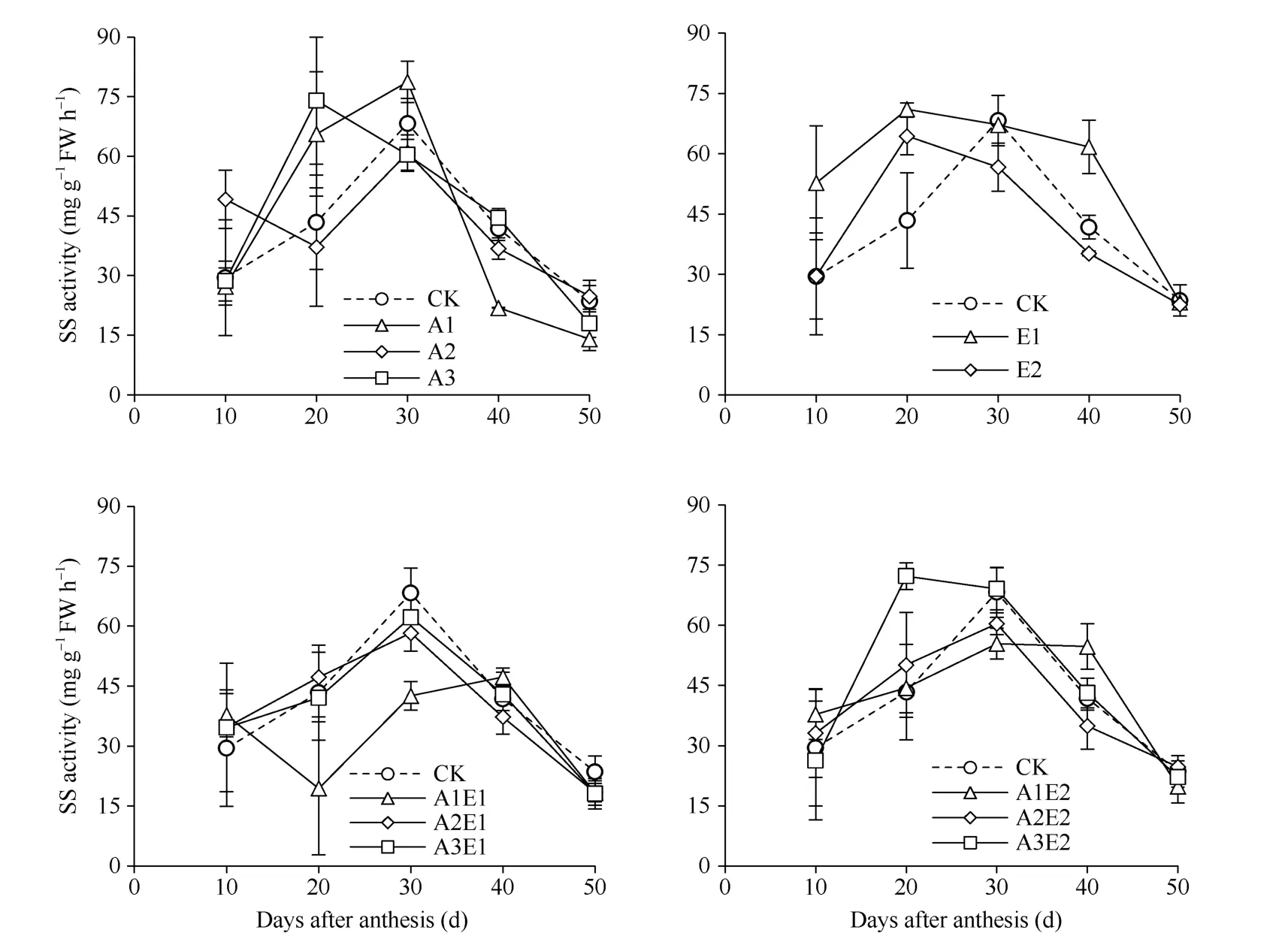
图6 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对籽粒蔗糖合酶(SS, 分解方向)活性的影响Fig. 6 Effects of 5-ALA, ETH, and 5-ALA-ETH on the activity of sucrose synthase (SS, cleavage direction) in 2018
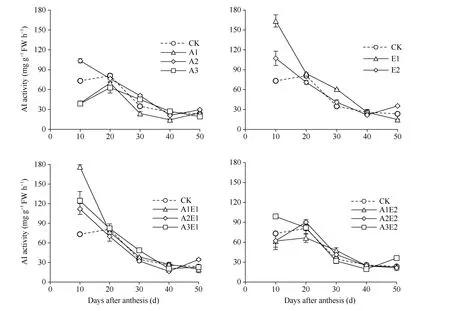
图7 2018年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对酸性转化酶(AI)活性的影响Fig. 7 Effects of 5-ALA, ETH, and 5-ALA-ETH on the activity of acid invertase (AI) in 2018

图8 2018年5-ALA、ETH 单剂和5-ALA 和ETH 复配剂处理对中性转化酶(NI)活性的影响Fig. 8 Effects of 5-ALA, ETH, and 5-ALA-ETH on the activity of neutral invertase (NI) in 2018
如表2 所示, 单施5-ALA 处理提高了玉米籽粒终极生长量a, 其中A2 处理相比对照提高6.2%, 且最大灌浆速率Gmax和灌浆速率最大时的生长量Wmax分别提高11.8%和6.2%, 而活跃灌浆天数P缩短4.9%。单施ETH 处理降低了籽粒终极生长量a, 其中E1 和E2 处理分别降低2.9%和2.0%。5-ALA-ETH复配剂处理中, A1E1、A2E1 和A2E2 处理a相比对照分别提高7.5%、1.5%和4.5%, 且A1E1 处理Wmax和P分别提高7.5%和3.5%, 而达到最大灌浆速率天数Tmax推迟2~3 d, 而A2E1 处理Tmax提前0~1 d, 且Gmax比对照提高3.5%。
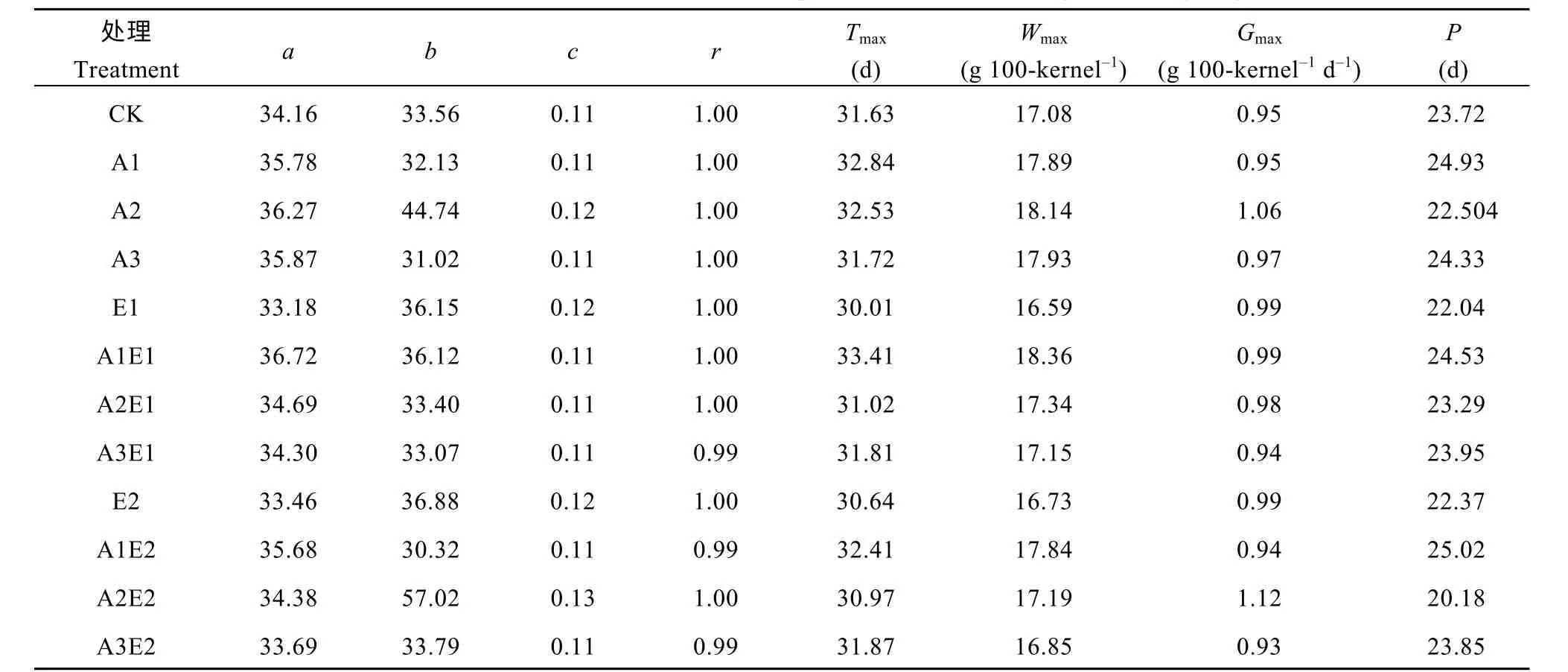
表2 2018年5-ALA、ETH 单剂以及5-ALA-ETH 复配剂处理对玉米籽粒灌浆特征参数的影响Table 2 Effects of 5-ALA, ETH, and 5-ALA-ETH on characteristic parameters of maize at grain-filling stage in 2018
2.4 5-ALA-ETH 复配剂处理对籽粒产量及其构成因素的影响
如表3 所示, 2018、2019年单施5-ALA 处理均提高了玉米产量, 其中2018年A2 处理比对照提高2.5%, 且穗长、穗粒数分别提高1.8%和4.7%, 秃尖长缩短41.7%, 而2019年A3 处理产量降低1.4%。单施ETH 处理, 2018、2019年E1 处理下产量分别提高2.9%和0.4%, 且2018年E1 处理穗长显著提高6.0%, 秃尖长显著缩短 52.4%, 穗粒数提高 8.2%,而E2 处理两年产量比对照分别降低2.2%和1.0%。5-ALA-ETH 复配剂处理中, A2E1 处理在2018、2019年均达到小区最高产(12,497.1 kg hm-2和14,078.9 kg hm-2), 与对照相比分别显著增产4.8%和5.4%,且在2018年穗长、穗粒数比对照分别提高4.7%和8.6%, 秃尖长显著缩短58.3%, 2019年穗长显著提高5.4%, 穗粒数提高6.2%, 秃尖长显著缩短50.4%。
3 讨论
3.1 5-ALA-ETH 复配剂处理对玉米功能叶蔗糖合成代谢的影响
玉米功能叶是玉米进行光合作用固定碳源的主要器官, 蔗糖作为光合作用的重要产物及其运输的主要形式参与代谢[20], 光合组织中蔗糖含量的多少直接影响籽粒中糖分含量, 近而影响淀粉含量[21],因此功能叶中蔗糖代谢在“源”端供应能力方面起关键作用。然而东北地区玉米季低温冷害频发, 其中灌浆中后期相比适宜温度平均降低7.5℃, 严重影响了灌浆速率和产量的提高[2]。有研究表明, 在低温[22]、弱光[10]条件下, 叶面喷施一定浓度5-ALA, 能够提高幼苗叶片的可溶性糖含量, 增强抗冷性、耐弱光性; 葡萄果实膨大期用5-ALA 处理, 5 d 后果实含糖量迅速增加[23]。本研究结果表明, 单施5-ALA 处理降低了生育期功能叶蔗糖合酶合成方向活性, 却提高蔗糖磷酸合酶活性, 提高生育期内功能叶的蔗糖含量, 其中A1 处理增幅最大。单施ETH 处理, 提高吐丝后SPS 活性, 但明显降低SS 活性和蔗糖含量。5-ALA-ETH 复配剂处理提高了吐丝后功能叶蔗糖磷酸合酶活性, 其中A2E1 处理提高了灌浆后期叶片蔗糖含量, 并且花前蔗糖磷酸合酶活性也高于对照, 提高花前蔗糖含量, 而降低了吐丝后20 d 内蔗糖含量。有研究表明, 叶片蔗糖含量的变化主要包括来自源器官的合成和向库器官的运输, 光合产物在花前主要作用于器官建成, 吐丝后主要供给籽粒的形成[24], 而A2E1 处理下吐丝后20 d 内叶片蔗糖含量降低可能与吐丝后功能叶蔗糖向籽粒运输有关, 但也有研究认为蔗糖含量增加会反向抑制蔗糖生成[25], 因此上述猜测需要在籽粒中进一步考证。综上, 5-ALA-ETH 复配剂处理主要通过提高SPS 活性促进功能叶蔗糖的合成, 提高花前功能叶蔗糖含量, 增强源端合成能力, 促进灌浆中后期功能叶蔗糖的积累, 增强灌浆后期玉米叶片的抗低温能力。
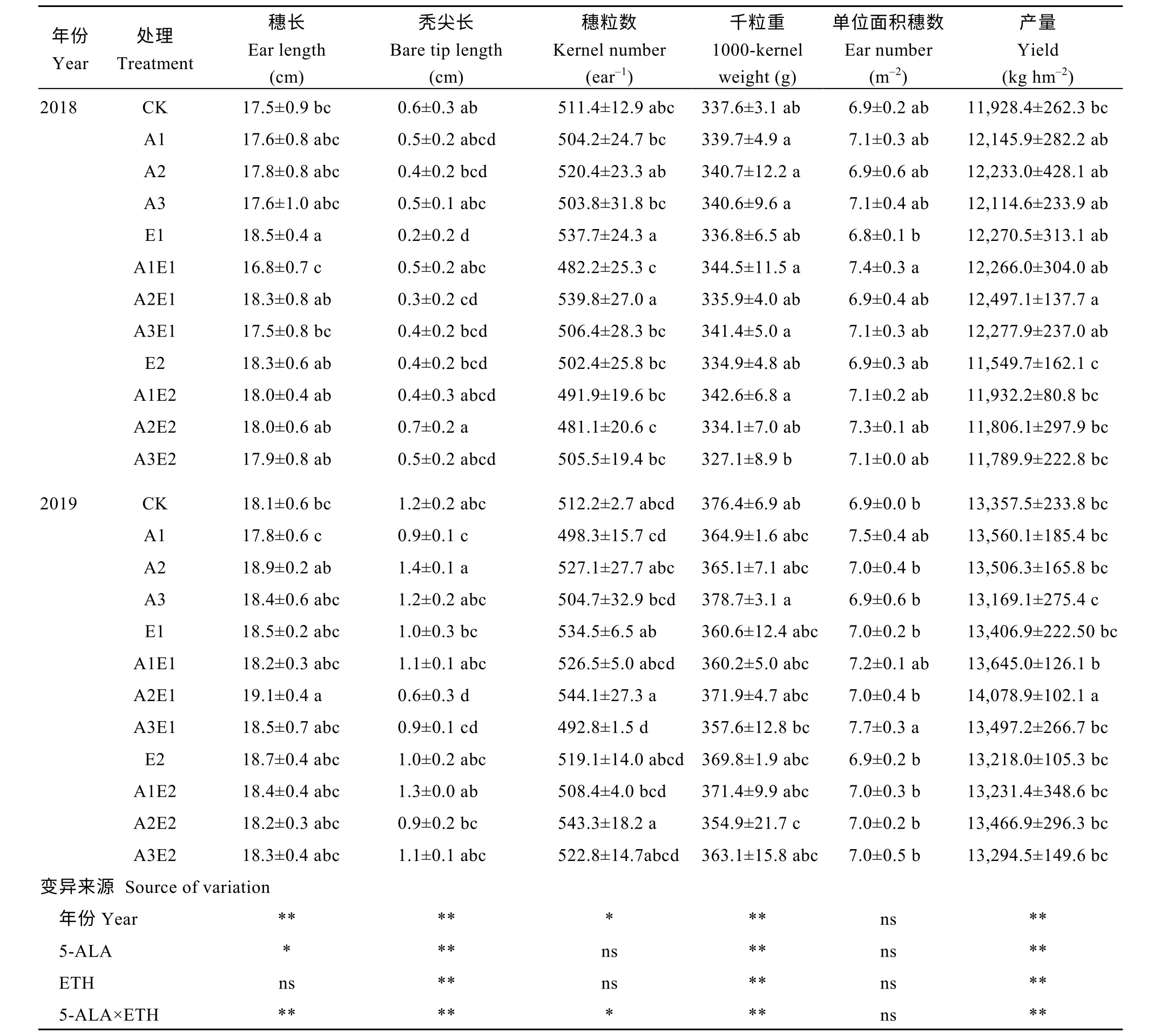
表3 2018年和2019年5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理对玉米产量及其构成因素的影响Table 3 Effects of 5-ALA, ETH, and 5-ALA-ETH on yield and yield components of maize in 2018 and 2019
3.2 5-ALA-ETH 复配剂处理对籽粒蔗糖分解方向活性的影响
作物高产不仅需要高的光合生产力, 还需要光合产物及时的运输分配, 籽粒中蔗糖积累过程包括来自源端的转运、自身合成以及降解[26]。Blum 等[27]研究认为, 小麦在逆境条件下, 茎秆中积累的碳水化合物能否有效运转、分配至籽粒, 对其产量形成起重要作用。本研究结果表明, 单施5-ALA 和ETH均提高灌浆前期籽粒蔗糖含量, 并且复配剂处理优于各单剂处理。小麦灌浆过程氮素通过增强蔗糖代谢相关酶活性来提高弱势籽粒重量[28]。籽粒中蔗糖经SS(分解方向)和转化酶(AI、NI)分解, 为淀粉合成提供原料[18]。籽粒中蔗糖降解可以反映“库”强度的高低[29], 而SS 能够催化蔗糖降解, 促进干物质的分配和储存, 可作为库强的指标[30]。转化酶作为蔗糖从韧皮部卸出的主要驱动力, 能够在源端和库端形成蔗糖梯度, 加速蔗糖向库端运输和分配[30-31]。本研究结果表明, 5-ALA、ETH 单剂和5-ALA-ETH 复配剂处理均可以提高灌浆前期籽粒SS (分解方向)活性, 增加库强。与之不同, 5-ALA 单剂处理AI 和NI 活性均明显降低, ETH 单剂处理NI 活性降低,但A2E1 复配剂处理AI 和NI 活性提高, 最终籽粒灌浆30 d 至50 d 蔗糖含量比对照降低。因此, A2E1处理一方面能够提高蔗糖分解方向SS 活性, 增大库容, 另一方面能够提高转化酶活性, 加快蔗糖向库端的转运。
3.3 5-ALA-ETH 复配剂处理对玉米籽粒灌浆和产量的影响
籽粒灌浆速率是决定玉米最终产量的重要因素[32]。有研究表明, 叶面喷施低浓度的5-ALA 可明显提高水稻籽粒的灌浆速率, 提高籽粒的起始生长势, 提早灌浆高峰[33]。小麦拔节期叶面喷施5-ALA,可以提高小麦灌浆中前期灌浆速率, 增加成穗数,提高产量[34]。乙烯利单剂处理降低籽粒灌浆起始生长势、缩短灌浆时间, 千粒重下降, 产量降低[35], 但也有研究表明乙矮合剂(主成分是乙烯利)能够提高高密度条件下玉米穗粒数, 增加产量[36]。本研究中,2018年5-ALA 单剂处理提高最大灌浆速率, 增加了穗长、千粒重和产量; 而2019年, 5-ALA 单剂处理增产效果不明显。单施ETH, 能够提高玉米穗长和穗粒数, 缩短秃尖长, 其中E1 处理2018年穗粒数明显增加, 秃尖长显著缩短, 产量提高, 而E2 处理2018、2019年产量均低于对照处理。5-ALA-ETH 复配剂处理中, A2E1 处理增产效果最佳, 且优于各单剂处理, 可以提早灌浆高峰, 提高最大灌浆速率,增加穗长和穗粒数, 显著缩短秃尖长, 且2018、2019年玉米产量均显著提高。
4 结论
5-ALA 与 ETH 可形成稳定的复合物, 其中A2E1 处理(5-ALA-22.50 g hm-2+ETH-450 mL hm-2)相比单剂处理和对照, 一方面通过提高蔗糖磷酸合酶活性, 促进功能叶蔗糖合成, 提高源强; 另一方面, 促进蔗糖由源向籽粒库的转运和降解, 大幅度提高吐丝后0~20 d 籽粒的蔗糖含量、籽粒蔗糖合酶(SS, 分解方向)、酸性转化酶(AI)以及中性转化酶(NI)活性, 增强籽粒库活性, 并提高籽粒最大灌浆速率,增加穗长和穗粒数, 显著缩短秃尖长, 显著提高产量。因此, 5-ALA-ETH 复配剂处理能够协调玉米功能叶和籽粒间的蔗糖代谢平衡, 促进光合产物向籽粒转运和降解, 加速灌浆, 对解决东北地区低温冷害造成源库代谢失调问题实现玉米增产稳产具有重要意义。
猜你喜欢
作物学报(2022年11期)2022-08-31 09:11:54
石油化工自动化(2022年3期)2022-07-13 02:46:12
热带作物学报(2021年9期)2021-11-08 11:55:35
核农学报(2021年3期)2021-02-22 08:47:58
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
茶叶科学(2018年5期)2018-10-17 01:56:42
化工管理(2017年36期)2017-03-07 06:59:17
热带农业科学(2015年2期)2015-04-13 04:12:06
现代检验医学杂志(2014年3期)2014-02-02 02:42:24
中国神经精神疾病杂志(2013年1期)2013-03-11 20:23:37
