
收获时间和覆土对杜仲短周期矮化林生物量的影响
2020-07-01 02:35:24丁欢欢刘慧东杜庆鑫杜红岩刘攀峰孙志强
经济林研究 2020年2期
丁欢欢,刘慧东,张 宁,王 璐,杜庆鑫,杜红岩, 刘攀峰,孙志强
(1.国家林业和草原局泡桐研究开发中心,河南 郑州 450003;2.南京林业大学,江苏 南京 210037;3.河南颐城控股有限公司,河南 郑州 450003;4.河南省科学技术信息研究院,河南 郑州 450003;5.经济林种质创新与利用国家林业局重点实验室,河南 郑州 450003;6.国家林业局杜仲工程技术研究中心,河南 郑州 450003)
高密度的短周期速生树种矮林可为生物质能源、燃料、医药及其他工业领域快速提供所需原材料[1-2]。例如,用于提供生物质能源和造纸原料的杨树Populusspp.、柳树Salixspp.和桉树Eucalyptusspp.的短周期矮林模式[3-5],以及供提取天然药用成分的叶用银杏Ginkgo biloba矮林模式[6-7]。杜仲Eucommia ulmoides是中国特有的贵重中药材和橡胶工业原料树种,杜仲胶产业化目前所面临的主要瓶颈问题之一是资源不足[8-9]。近年来,围绕杜仲高密度短周期矮林种植模式,学者们从多个角度开展了系统研究。Zhu 等[10]报道,在同一时期(10 a 以内)叶用林模式的杜仲叶片、树皮和茎的总生物量和次生代谢物积累量显著高于杜仲传统乔木栽培模式。有研究结果表明,对杜仲矮林进行叶面喷施外源激素,不仅可提高叶片、树皮等的产量,还显著提高了杜仲胶的产量,第1 轮伐期胶产量182.90 kg/hm2,为对照的5.01倍[11-12],第2 轮伐期的叶片和树皮杜仲胶理论总产量最高达到391.00 kg/hm2,是对照的1.42 倍[13]。高密度短周期杜仲矮林模式有望成为杜仲胶产业化资源林培育的主要模式之一。
有学者指出,短周期矮林的收获方式和收获时间等会影响到收获的成本和效益,以及后续的生物量[14]。Santiago 等[15]的研究结果表明,收获季节对短周期轮作桉树和杨树树桩的存活和生长活力均有相当大的影响,建议在冬季收获,与Souza 等[1]得出的结论一致。Souza 等[1]指出,桉树和棉白杨Populus deltoides冬季收割比夏季收割其萌条存活率高,而收割方式对其根蘖再生无影响。Hytonen[16]的研究结果也表明,在7月底或8月初砍伐异花柳树(exotic willows)会对其存活、高生长和生物量产生不利的影响。Sims 等[17]报道,在新西兰北岛中部,短期轮作的桉树通常应在10—11月的春季收获。与叶用银杏短周期矮林主要供工业化提取有效成分类似,杜仲矮林也是每年进行收割以获得叶片和树皮,来提取有效成分和杜仲胶。何文广等[18]通过对杜仲叶林模式6—10月生物量动态变化的调查分析,建议以杜仲叶片为原料的加工产业,应在9月采摘叶片。目前,关于不同收获时间对杜仲短周期矮林后续生长及生物量影响的研究未见报道。
本研究中,以2015年营建的杜仲矮林为对象,2017年在单一密度的杜仲矮林中设定不同的收获时间和标准样株进行收割平茬,2018年系统调查样株的发芽时间、生长指标和生物量,并进一步比较不同种植密度杜仲矮林地上部分生物量,旨在确定杜仲矮林模式适宜的收获时间和种植密度,为杜仲矮林的产业化利用和机械化收割提供参考。
1 材料与方法
1.1 试验地概况
试验地位于中国林业科学研究院经济林研究开发中心孟州基地(112°42′58′′E,34°51′38′′N)。该基地南临黄河,属黄河滩地,大陆性暖温带季风气候,年降水量约549.9 mm,年平均气温14.3 ℃,无霜期224 d,具有中部平原地区黄河故道沙地典型立地特征。沙壤土,土壤(0 ~40 cm 土层)的理化性质:pH 8.67,有机质含量4.07 g/kg,总氮含量0.03 g/kg,有效氮含量18.79 mg/kg,有效磷含量4.80 mg/kg,有效钾含量156.62 mg/kg。
1.2 试验设计
杜仲矮林于2015年3月营造,总面积 3 000 m2。采用宽窄行栽植模式,宽行距1 m,窄行距0.5 m,窄行内株距分别为0.2、0.4、0.6、0.8和1.0 m,栽植密度分别相当于66 667株/hm2(D1)、33 333 株/hm2(D2)、22 222 株/hm2(D3)、 16 667 株/hm2(D4) 和13 333 株/hm2(D5)。每年春季平茬。从第2 轮伐期开始,对栽植密度D3、D4、D5 的植株均保留4 个萌条,栽植密度D2 的植株留2 ~3 个萌条,栽植密度D1 的植株保留1 个主干。
2017年春季平茬后,栽植密度D1 处理组按完全随机区组试验设计,设定4 个收获时间,每个时间点收获时分成2 组,一组平茬后将根桩覆土(记为C),另一组不覆土(记为NC)。按收获时间,设置8 个处理,分别为8月30 日(C1 和NC1)、9月15 日(C2 和NC2)、9月30 日(C3和NC3)和10月15 日(C4 和NC4)。2017年秋季,按上述设定的时间,对每个处理进行收割并随机平茬10 株,重复3 次。每个收获时间点合计平茬60 株。所有植株均进行标记。
1.3 生长指标和生物量的测定
2018年春季,观察标记样株并逐一记录初始发芽时间。5月3 日,测定初始苗高和初始基径,随后每隔20 d 测量1 次苗高(cm)和基径(mm),至9月6 日,共计120 d。适时浇水、除草,试验期间不施肥。2018年,逐一收获并称量各样株的全部叶片、树皮和茎的鲜质量。将采集的叶片、树皮和茎(取中间20 cm 长的茎)在65 ℃条件下烘干后,再分别称取其质量。采用差重法计算含水率。
为比较不同栽植密度下生物量的差异,10月15 日在各栽植密度的林分中随机选取5 株植株,按上述方法测量叶片、树皮和茎的鲜质量,然后分别计算单位面积叶生物量、单位面积皮生物量、单位面积茎生物量。
1.4 数据统计分析
采用单因素方差分析方法检验不同收获时间的植株各生长性状的差异,并进行Duncan 多重比较。运用Student-T检验方法对同一收获时间的覆土与未覆土组次年初始和最终苗高,以及叶片、树皮和茎生物量进行差异性检验。此外,采用单因素方差分析方法比较不同密度下的生物量差异,并进行Duncan 多重比较。采用SPSS 软件(20.0 版)进行数据分析。
2 结果与分析
2.1 收获时间和覆土对次年植株发芽时间的影响
不同处理组中样株的初始发芽时间和持续时间略有差异(图1)。10月15 日收割的苗木次年整体上发芽最早,无论平茬后根桩覆土与否,均在次年3月14 日发芽。明显的区别在于,8月30 日和9月30 日收割的苗木次年发芽时间较10月15日收割的苗木整体上约晚1 周,而9月15 日平茬后根桩覆土苗木发芽时间较10月15 日收割的苗木约晚10 d。8月30 日、9月15 日和9月30 日收割后根桩覆土的苗木,整体上比未覆土苗木发芽持续时间长,如9月15 日平茬的苗木,覆土组发芽时间从3月17 日持续至4月3 日,而未覆土的苗木发芽时间较为集中,3月22—24 日,仅3 d 时间。 8月30 日、9月30 日和10月15 日平茬的苗木,覆土组最晚发芽时间各有差异,其中最晚为4月1日,而未覆土组时间相同,最晚均为3月27 日。
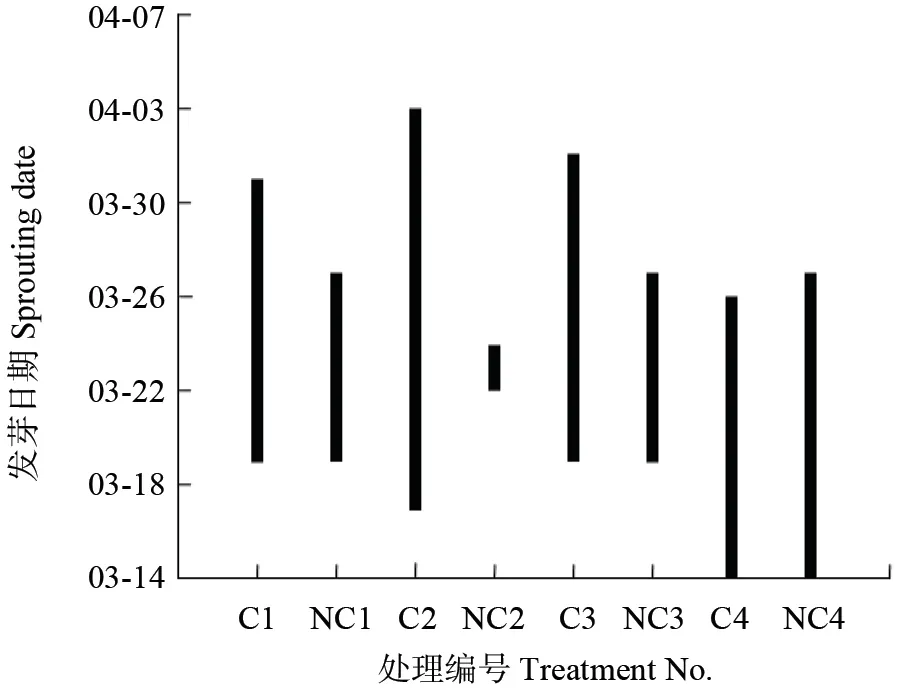
图1 不同收获时间下覆土与未覆土组杜仲次年发芽时间Fig.1 Sprouting dates in E.ulmoides harvested on different dates in mulching and non-mulching treatments
2.2 收获时间和覆土对植株生长指标的影响
2.2.1 对植株苗高的影响
在不同收获时间点,覆土组初始苗高0.42 ~ 0.47 m,最终苗高2.36 ~2.53 m,各收获时间的苗高间均无显著差异;未覆土组初始苗高0.41 ~ 0.50 m,最终苗高2.25 ~2.43 m,各收获时间的苗高间均无显著差异(图2)。整体上,覆土后的最终苗高略高于未覆土组,其中9月30 日平茬后覆土的苗高高于其他3 组。
T检验结果显示,8月30 日收获的样株,覆土和未覆土(C1 和NC1)组的初始苗高有显著差异(T=-2.194,Df=18,P=0.042),其余收获时间点,无论是否覆土,初始苗高和最终苗高均无显著性差异。
2.2.2 对植株基径的影响
不同收获时间点,覆土组初始基径4.71 ~ 5.33 mm, 无显著差异; 最终基径14.68 ~ 17.56 mm,差异显著(F=2.863,P=0.05)。其中,覆土组10月15 日处理(C4)的植株基径最大,其次为8月30 日、9月30 日和9月15 日处理。未覆土处理组植株的初始基径4.59 ~5.18 mm,无显著差异;最终基径13.95 ~16.79 mm,差异显著(F=2.964,P=0.046)。9月30 日处理(C3)的植株基径最大,其次为10月15 日、8月30 日和9月15 日处理。整体上,覆土组最终基径均值(16.07 mm)略高于未覆土组(15.52 mm)(图3)。

图2 不同收获时间下覆土与未覆土组杜仲苗高生长曲线Fig.2 Growth curves of seedling heights in E.ulmoides harvested on different dates in mulching and non-mulching treatments

图3 不同收获时间下覆土与未覆土组杜仲基径生长曲线Fig.3 Growth curves of ground diameters in E.ulmoides harvested on different dates in mulching and non-mulching treatments
2.3 收获时间和覆土对植株生物量的影响
不同收获时间下,覆土组叶片(F=3.255,P=0.034)、树皮(F=5.265,P=0.004)和茎(F=4.108,P=0.014)生物量呈显著性差异,而未覆土组无显著性差异(P>0.05)(表1)。不论覆土与否,9月30 日(C3)平茬苗木叶片的次年生物量最大,覆土组为3 433.35 kg/hm2,未覆土组为 3 457.20 kg/hm2。在10月15 日收获时,覆土组树皮和茎的次年生物量最大,分别为1 269.70 和 7 776.97 kg/hm2,其次为9月30 日,生物量分别为1173.69 和7 720.93 kg/hm2,2个处理组间的生物量无显著性差异。9月30 日收获的处理中,未覆土组树皮和茎的次年生物量最大,分别为 1 169.52 和8 321.09 kg/hm2(表1)。
T检验结果显示,8月30 日平茬后,叶片生物量在覆土与未覆土组间差异显著(T=-2.250,Df=10,P=0.048),而10月15 日平茬苗木的树皮生物量在覆土与未覆土组间差异极显著(T=3.341,Df=17,P=0.004),其他处理组间无显著性差异 (表2)。
2.4 栽植密度对次年植株生物量的影响
不同栽植密度下同一时间收获的杜仲叶片(F=9.201,P<0.001)、树皮(F=18.749,P< 0.001)和茎(F=15.773,P<0.001)的生物量均值存在极显著差异。根据叶片、树皮和茎生物量由高到低排列,各栽植密度依次均为D2、D1、D4、D3、D5。其中,栽植密度D1 和D2 的杜仲茎和叶片的生物量之间无显著差异,但显著高于其他3 个栽植密度,栽植密度D2 杜仲树皮的生物量显著高于其他4 个栽植密度。栽植密度D2 杜仲的叶片、树皮和茎生物量均值分别为4 656.71、 1 967.28 和15 121.02 kg/hm2;栽植密度D1 杜仲叶片、树皮和茎生物量均值分别为3 982.07、1 633.01 和13 037.27 kg/hm2。 栽植密度D5杜仲的叶片、树皮和茎生物量最低,其均值仅为 2 432.05、929.69、和7 561.77 kg/hm2(图4)。

表1 不同收获时间下覆土与未覆土组杜仲生物量†Table 1 Biomass in E.ulmoides harvested on different dates in mulching and non-mulching treatments
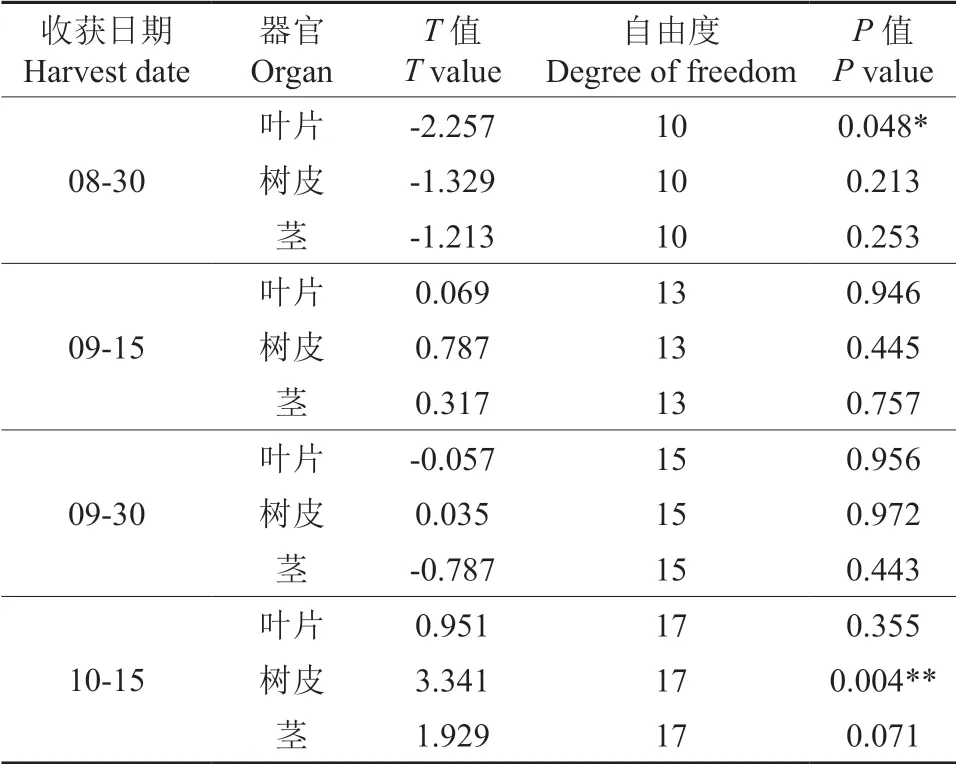
表2 同一收获时间下覆土与未覆土组杜仲生物量T 检验 结果†Table 2 T test result of biomass in E.ulmoides harvested on the same date in mulching and non-mulching treatments
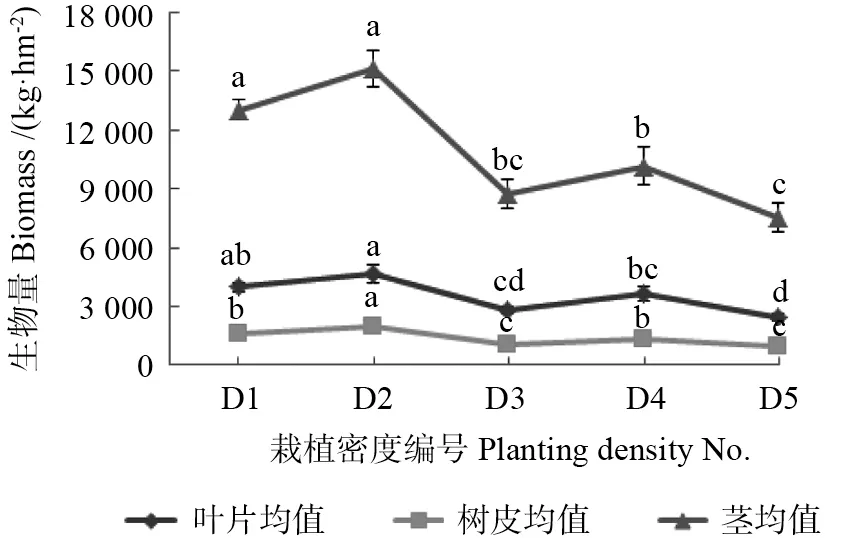
图4 不同栽植密度下杜仲各器官的生物量Fig.4 Biomass of different organs in E.ulmoides under different planting densities
3 结论与讨论
本研究结果表明,收割时间显著影响杜仲矮林次年植株的发芽时间及整体发芽的持续时间(图1)。尽管同一时间点收割后根桩覆土与未覆土的最终苗高之间差异不显著,但整体上覆土后的苗高比未覆土的略高(图2),最终的基径表现与苗高相类似(图3)。对不同时间点收割后根桩进行覆土,次年叶片、树皮和茎的生物量均存在显著或极显著差异,其中,2017年9月30 日及其之后收割的矮林,根桩覆土后次年叶片、树皮和茎的生物量普遍较其他处理高(表1);而不同时间点平茬后根桩未覆土,2018年收获时,各器官生物量的差异均不显著。另外,栽植密度66 667 和 33 333 株/hm2条件下,杜仲的叶片、树皮和茎生物量显著高于其他3 个栽植密度(22 222、16 667和13 333 株/hm2)(图4)。
封顶是杜仲生长发育规律之一[19]。在高密度平茬建园的条件下,实生苗和‘华仲11 号’杜仲的幼树均存在封顶与二次梢现象[20]。本研究中,自5月3 日开始测量初始苗高,从120 d 的生长曲线可知,高密度下的杜仲苗木顶端优势明显,表现出较强生长特性。9月30 日前平茬的杜仲并未完全停止生长(田间可见有些平茬后的苗木再次萌芽,特别是8月30 日平茬的苗木,但冬季受冻后死亡),此时萌条会消耗根部营养[21],从而影响次年的萌发和生长。10月15 日杜仲已经完全停止生长,此时进行平茬,不会引起再次萌发,根部的营养不会丧失,因此,该时间点平茬的苗木次年萌芽最早。针对桉树矮林的最佳收获时间,有学者强调,收获时要避免冬季低温对幼嫩萌条的伤害,或避免夏季干旱期因缺水导致无法萌发或生长[17]。同时,Blake[22]经研究证实,在夏季收获时,桉树的新萌芽数和萌条长度均有所下降。Santiago 等[15]的研究结果表明,在冬季收获的根桩可观察到更大的萌蘖,可能归因于在收获时储存在根系中糖类储备的高可用性,例如,在冬季具有明确休眠期的胡杨Populusspp.有更好的平茬反应率和最大的萌芽活力。
对平茬后的根桩进行覆土有利于保持水分[23]。覆土后,尽管次年萌芽持续时间较长,但苗木的苗高普遍较高,生物量得以整体提高。若平茬时间早于9月30 日,因苗木未完全进入休眠而消耗部分养分,导致其次年的生长受到影响,从而降低了生物量积累。因此,应尽量待杜仲矮林苗木进入休眠期再进行收割。
刘慧东等[24]在其研究报道中强调,每年为获得更高的叶片、树皮和茎产量,短周期杜仲矮林栽培模式应选择更高的种植密度,如66 667 株/hm2等,或留置6.7 个/m2以上的茎。马顺兴等[25]在其研究报道中提到,杜仲短周期矮林在每年9—10月轮伐1 次,第3年开始低密度下根桩可保留3 ~5个主茎。Zhu 等[26]经研究发现,在多茎条件下,茎干高度可达2 ~3m。本研究中,在33 333 株/hm2(D2)的栽植密度下,每个根桩留置3 个主枝。因此,不难理解本研究中66 667 和33 333株/hm2栽植密度下杜仲的总体生物量远高于其他各栽植密度的生物量(图4)。
综上所述,在不同时间点收获杜仲矮林后,从次年萌芽时间、生长指标和生物量积累,以及栽植密度综合考虑,应尽量待苗木进入休眠期进行收割,即9月30 日以后进行收割。应及时进行平茬后根桩的覆土作业。为便于机械化收割,杜仲短周期矮林最好采用宽窄行栽植模式,宽行距1 m, 窄行距0.5 m,窄行内株距0.4 m,栽植密度相当于33 333 株/hm2,每个根桩留置2 ~3 个主枝,使年生物量达到最大。
本研究中调查时间仅为1 个生长季,并未对不同收获时间的杜仲短周期矮化林情况进行3 ~5 a的重复观测和调查。宋海云等[27]在8—9月设置了5 个时间点,研究了不同日期采收的澳洲坚果其必需氨基酸的差异性分布情况,结果表明采摘时间会影响澳洲坚果的营养品质。故下一步将研究不同收获时间杜仲短周期矮林叶片和树皮中有效活性成分和杜仲胶的变化,并结合本试验结果进行重复性检验,以生物量、有效活性成分和杜仲胶为主要指标,得出杜仲短周期矮林的最佳收获时间。
猜你喜欢
童话王国·文学大师班(2022年3期)2022-04-19 14:01:36
江西农业大学学报(2020年6期)2021-01-08 02:35:50
中成药(2018年8期)2018-08-29 01:28:14
小哥白尼(野生动物)(2018年4期)2018-08-01 03:41:14
现代园艺(2018年1期)2018-03-15 07:56:21
小溪流(画刊)(2017年7期)2017-08-09 19:49:55
农业知识(2017年9期)2017-04-10 07:07:58
陕西画报(2016年1期)2016-12-01 05:35:28
童话王国·文学大师班(2016年6期)2016-11-10 06:10:33
小雪花·小学生快乐作文(2015年9期)2015-09-29 23:06:01
