
基于COⅡ基因分析鲁山柴鸡遗传多样性
2020-06-30 13:44:30王莲哲白梦彩李冰冰
河南城建学院学报 2020年2期
王莲哲,白梦彩,毛 锐,李冰冰
(河南城建学院 生命科学与工程学院,河南 平顶山 467036)
地方家禽遗传资源是生物多样性的重要组成部分,是未来家禽品种改良和资源保护与持续发展的基础[1]。鸟类线粒体DNA(Mitochondrial DNA,mtDNA)具有母系遗传、多态性高和不易发生重组等特性,线粒体相关基因作为分子标记已被广泛用于家禽物种鉴定、系统发生、遗传多样性和种群结构等方面的研究[2-3],如利用生物体线粒体DNA中一段保守片段作为DNA条形码可以对物种进行快速准确鉴定,较为常见的是线粒体CO I、COⅡ、ND1、Cytb等保守基因[4-7]。线粒体细胞色素c氧化酶亚基II(COⅡ)基因中的部分区域由于变异速率适中及具有完善的通用引物,近年来被用作物种鉴定及群体遗传进化的理想分子标记,在物种鉴定及遗传进化分析中发挥了重要作用[8]。
河南省鲁山县隶属伏牛山脉,地处中原地区,气候温和、生态环境优良。鲁山本地柴鸡由于地理环境因素造就生殖隔绝,长期进化形成了一种稳定遗传的品系。鲁山鸡种适应性强、抗病性强、耐粗饲,觅食能力强,蛋肉品质优良,作为地方品种是宝贵而重要的遗传资源。本研究以鲁山本地柴鸡为研究对象,分析线粒体COⅡ基因序列碱基组成和多态性,并构建鲁山鸡和其他鸡种的系统发育树,探讨分析鲁山鸡遗传起源及遗传多样性特点,为地方鸡品种资源保护和开发利用提供理论支撑。
1 材料与方法
1.1 材料
随机选取平顶山市鲁山县柴鸡36只,翅下静脉取血。血样采集后添加抗凝剂,用血液DNA提取试剂盒(天根生化)提取全基因组DNA,-20 ℃保存备用。
1.2 DNA提取与PCR扩增
根据GenBank公开发表的红色原鸡COⅡ基因序列设计合成引物,上游COⅡ-F:CCACTCCCAACTAGGCTTTC,下游COⅡ-R:GACAGTAGTGAGGATCAG,引物由武汉金开瑞生物公司合成。PCR反应条件设置为:95 ℃,3 min,95 ℃变性30 s,56 ℃ 退火30 s,72 ℃延伸60 s,34个循环,72 ℃延伸5 min,12 ℃保存。片段大小669 bp。扩增产物经1.0%琼脂糖凝胶电泳检测,样品送武汉金开瑞生物公司测序。
1.3 统计与分析
测序结果通过软件Bioedit读取,双向测序校正及人工逐碱基校对。从NCBI数据库下载相关鸡种COⅡ基因,使用Clustal_X进行序列比对,MEGA6.0[9]软件采用邻接法构建系统进化树,1 000次自举检验。用DnaSP 6.0定义单倍型,同时计算核苷酸差异均数、核苷酸多样性、单倍型多样性及核酸变异位点数等。应用Network 5.0构建中介网络图(median-joining)[10]。

图1 COⅡ基因PCR扩增图
2 结果与分析
2.1 COⅡ基因序列扩增
鲁山鸡随机取样翅下静脉取鸡血,用试剂盒提取鸡血总DNA,并用特异性引物扩增COⅡ基因,经琼脂糖凝胶电泳检测。图1结果显示:COⅡ基因条带单一,片段大小在660 bp左右,符合预期。重复PCR扩增,将扩增产物送公司测序。
2.2 鲁山鸡COⅡ基因序列分析
经过PCR扩增及测序校对,剔除首尾部分低质量序列,得到CO Ⅱ序列570 bp进行分析。鲁山鸡群体CO Ⅱ全序列变异区在26~570 bp,共发现16处多态位点,分别为26、36、72、96、112、114、146、181、191、354、358、393、462、471、569、570位点,占分析位点总数的2.81%,均为简约信息位点。碱基组成分析显示4种碱基含量分别为22.7%(T)、33.1%(C)、29.9%(A)和14.3%(G),其中A+T含量(52.6%)稍高于G+C含量(47.4%),基本保持平衡。变异类型转换7处,颠换9处。16个多态位点界定为7个单倍型,分别为Hap1-Hap7。其中,单倍型Hap1有18个个体,占50%,为优势单倍型;单倍型Hap2、Hap3、Hap6各有4个个体;单倍型Hap4、Hap5、Hap7各有2个个体(见表1)。

表1 鲁山鸡COⅡ基因多态位点及单倍型
2.3 鲁山鸡单倍型网络
基于中间连接法构建的鲁山鸡CO Ⅱ 序列单倍型网络图(见图2),各单倍型以H1为中心,连接形成星状发散(star-shape)结构。其中,单倍型H1为优势单倍型,单倍型H2和H7遗传距离较远,H3、H4、H5、H6与H1遗传较近。核苷酸多样性为0.004 41,单倍型多样性为0.724。群体扩张的Tajima′s D数据为-1.142 13,该数据未达到显著水平,说明该群体依然符合中性选择,未经历大规模的种群扩张。
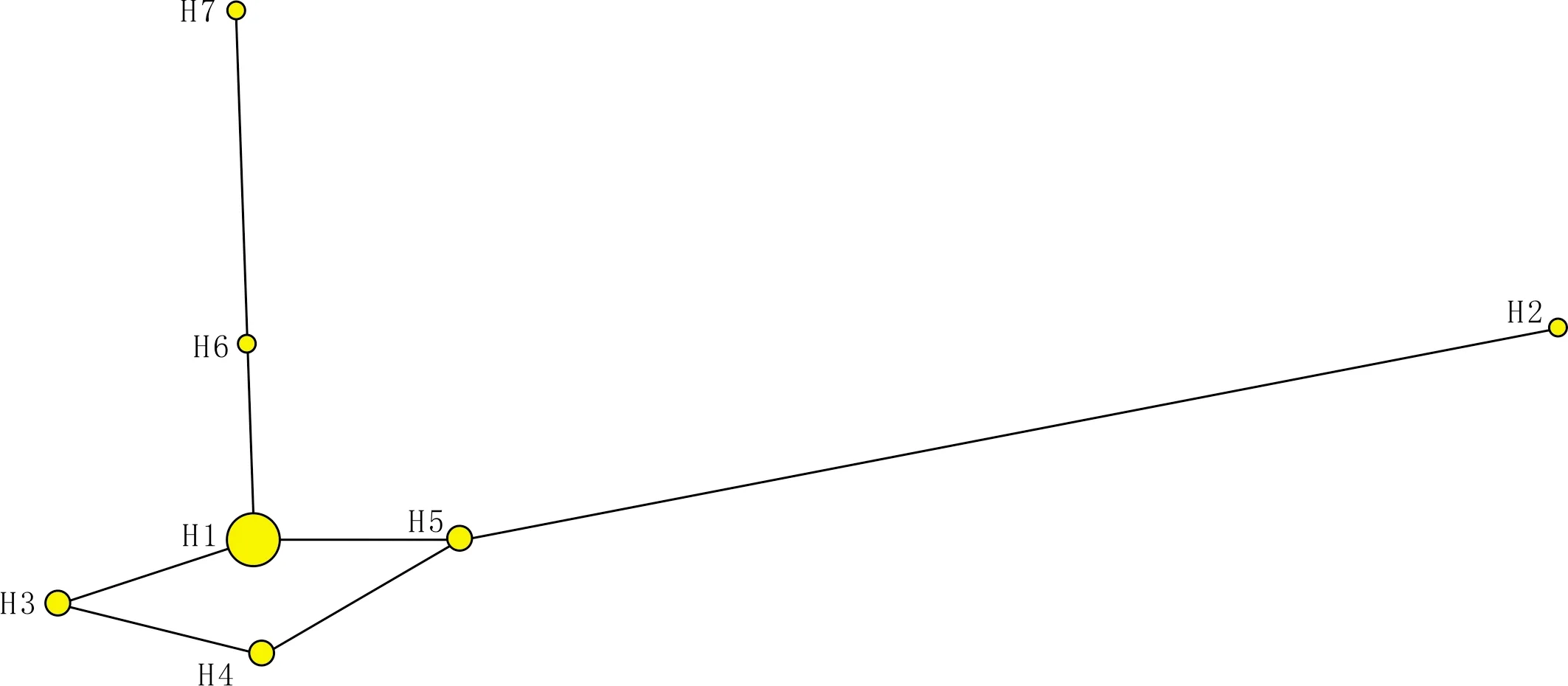
图2 鲁山鸡单倍型网络关系
2.4 鲁山鸡系统发育分析
基于鲁山鸡线粒体COⅡ基因序列分析,将鲁山鸡7种单倍型与NCBI数据库中下载的其他鸡种的COⅡ基因构建系统进化树(见图3)。结果显示:鲁山鸡7种单倍型聚在一类,且与原鸡印尼亚种聚为一类,说明其有更近的亲缘关系,在鲁山鸡进化过程中,原鸡印尼亚种贡献较大。
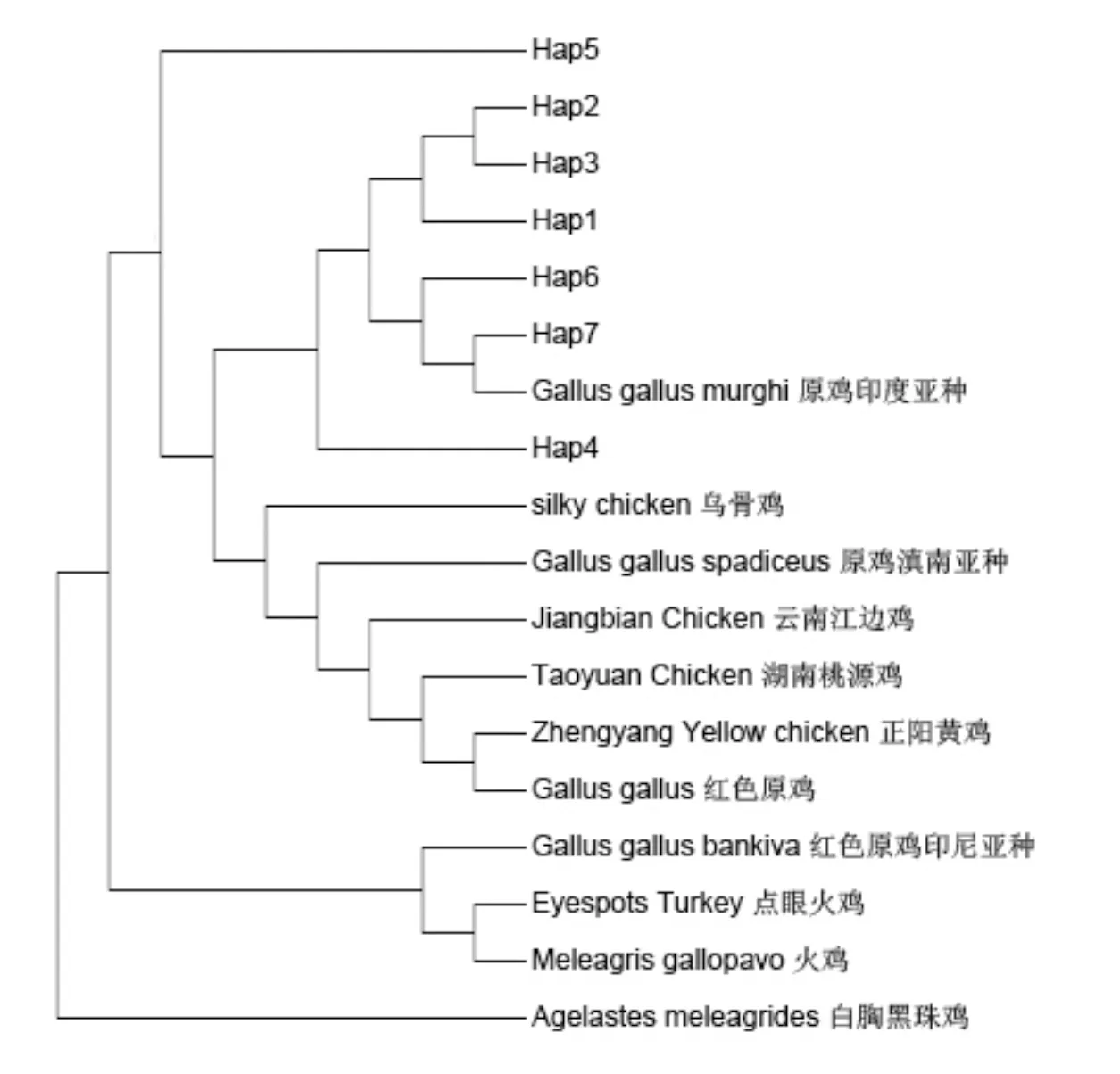
图3 鲁山鸡COⅡ基因系统进化树
3 讨论
遗传多样性是生物多样性形成的基础,是物种或其群体持续生存和环境适应的结果[11]。在农业生产中,正确认识和评价不同农副产品的遗传多样性水平,是开展保护及可持续开发利用的基础。鲁山鸡作为平顶山本地的特色品种,其养殖与资源综合利用是当地经济发展的重要支撑。通过分子标记研究地方鸡种遗传多样性有助于当地特色物种资源的保护与开发。以往有关鸡线粒体遗传多样性的研究主要集中在CO I基因[12-14],鸡COⅡ基因研究少有报道。而COⅡ基因作为新开发的分子标记已广泛应用于其他物种鉴定[15-16]。本研究中基于线粒体COⅡ基因序列分析,对鲁山鸡遗传多样性和遗传进化进行了分析。COⅡ基因扩增片段长度为570 bp,4种碱基含量中A+T含量(52.6%)稍高于G+C含量(47.4%),基本保持平衡,表明该基因碱基和密码子的使用没有一定的偏向性。36只个体共发现16处单核苷酸多态位点,7种单倍型,变异类型转换7处,颠换9处,基本平衡。结果表明CO Ⅱ基因比较保守,序列分析区分度较高,可以作为分子标记进行鸡种品种鉴定及多样性分析。
随着对外交流的日益频繁,鲁山鸡群体乱杂态势加剧,鲁山鸡作为地方品种其遗传多样性没有分析报道,其遗传进化特点也不明确,因此保护不力。本研究显示,鲁山鸡核苷酸多样性为 0.004 41,每位点核苷酸多态性指数0.006 77,平均核苷酸差异2.514。单倍型多样性为0.724,结果表现出鲁山鸡具有较高的遗传多样性[17]。该鸡种Tajima′s D数据为-1.142 13,不显著,说明该群体依然符合中性选择,未经历大规模的种群扩张[18]。
构建系统进化树是追溯物种起源的良好方法。坝上长尾鸡用此方法分析表明,追溯其与中国原鸡亲缘关系最近,中国原鸡对坝上长尾鸡进化起到重要作用[19]。系统进化显示茶花鸡祖先可能为红色原鸡,主要贡献为原鸡滇南亚种[20]。单倍型网络关系结果显示,鲁山鸡7个单倍型以单倍型Hap1为优势类型,除单倍型Hap2和Hap7进化距离稍远,其他单倍型进化距离近。系统进化分析,7个单倍型均聚于一个分支,且与原鸡印尼亚种有更近的亲缘关系。由此可见,鲁山鸡在长期的形成过程中其母系起源较为简单,其祖先可能为红色原鸡,主要贡献为原鸡印尼亚种。
4 结论
以鲁山本地柴鸡为对象,利用线粒体基因COⅡ序列,通过测序及比较分析,研究了该序列的核苷酸多态性、单倍体多态性、群体间的遗传多样性、鲁山鸡与其他鸡种的系统进化关系。结果表明:鲁山鸡核苷酸多样性为 0.004 41,单倍型多样性为0.724,表现出鲁山鸡具有较高的遗传多样性。鲁山鸡祖先可能为红色原鸡,进化过程主要贡献为原鸡印尼亚种。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
意林(2019年17期)2019-10-07 12:28:52
上海农业学报(2019年4期)2019-09-18 06:46:48
浙江农业学报(2017年4期)2017-04-26 02:28:22
新天地(2017年4期)2017-04-17 20:46:02
森林与人类(2016年10期)2017-04-11 06:52:20
浙江农业学报(2016年7期)2016-06-15 20:30:08
当代旅游(2016年3期)2016-05-13 13:45:53
