
传染性造血器官坏死病毒Sn1203 株全基因组序列及系统进化分析
2020-06-30 05:46:34
水产学杂志 2020年2期
(中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
传染性造血器官坏死病(infectious hematopoietic necrosis,IHN)是严重威胁野生和人工养殖鲑鳟的急性、全身性传染病[1,2]。根据感染宿主、病毒株和鲑鳟鱼养殖环境的不同,IHN 造成的死亡率超过90%[3,4]。20 世纪50年代,美国华盛顿和俄勒冈州的红鲑Oncorhynchus nerka 养殖场首次报道了该病,并迅速沿太平洋海岸扩散至北加州及爱达荷州[5]。随着鱼苗及成鱼的贸易流通,目前IHN 已经蔓延到全世界多个国家,如日本[6]、伊朗[7]、加拿大[8]、韩国[9]、俄罗斯[10]、荷兰[11]等。1985年我国辽宁省的一个虹鳟鱼幼仔孵化场首次发现IHN 疫情,目前已经扩散至多个鲑鳟主养区,给我国的鲑鳟养殖造成了严重的经济损失[12]。
IHN 的病原为传染性造血器官坏死病毒(infectious hematopoietic necrosis virus,IHNV)。它由不分节段的单股负链RNA 组成,归属于弹状病毒科Rhabdoviridae,鲑科诺拉弹状病毒属Salmonid Novirhabdovirus[5,13]。IHNV 全基因组大小约为11100 nts,共编码6 个病毒蛋白,分别为5 个结构蛋白:核蛋白(N)、磷蛋白(P)、基质蛋白(M)、表面糖蛋白(G)、聚合酶蛋白(L),以及1 个非结构蛋白(NV)[14]。病毒基因组与核蛋白N 紧密连接,形成病毒粒子的核心结构后与磷蛋白P 和聚合酶蛋白L 一起参与病毒的复制[15]。目前,世界各地发现的IHNV 分别以N、G、NV 基因的部分或全部核苷酸序列,进行详细的系统进化分析[5,16,17]。
本课题组于2012年从中国东部的某虹鳟养殖场分离得到IHNV-Sn1203 病毒株,对其进行了大量研究,包括疫苗、致病机理和免疫学研究等。为了更好地了解该病毒株的分子生物学特征,对IHNV-Sn1203 病毒株的全基因组序列进行克隆及测序,并结合生物信息学分析该毒株的全基因组序列、基因组编码蛋白、基因组末端和未翻译序列、以及病毒基因同源性和系统发育,以期为我国IHNV病毒株的分子生物学特性及分子进化研究提供参考。
1 材料与方法
1.1 病毒株和细胞株
自中国东部某虹鳟养殖场分离的IHNV-Sn1203 病毒株(GenBank No:KC660147.1)经分离鉴定后保存于本实验室。实验用鲤上皮瘤细胞(epithelium papulosum cyprini,EPC)由中国水产科学研究院长江水产研究所鱼类病害教研室曾令兵教授惠赠。
1.2 引物设计
用DNAMAN 软件,比对GenBank 中收录的IHNV 基因组全序列信息。根据比对结果,利用Primer Premier version 5 软件,在IHNV-HLJ09 株(Accession number JX649101)全基因组序列的基础上,设计覆盖全基因组序列的重叠引物,用于IHNV-Sn1203 病毒株全基因组序列克隆。所有引物均由吉林省库美生物科技有限公司合成(表1)。

表1 IHNV-Sn1203 基因组克隆所用引物Tab.1 Primers sequences used in IHNV-Sn1203 genomic sequence clone
1.3 IHNV-Sn1203 病毒株全基因组序列克隆
本研究将IHNV-Sn1203 基因组序列分成5 个片段进行克隆。使用GenEluteTM总RNA 提取试剂盒(Sigma 公司),从细胞培养上清中提取IHNV-Sn1203 病毒株总RNA。以病毒总RNA 为模板,利用SuperScript III One-Step RT-PCR Platinum Taq HiFi Kit 试剂盒,及表1 中引物分段克隆IHNV-Sn1203 全基因组序列。为了保证IHNV-Sn1203基因组末端序列的准确性,采用SMARTTMRACE 试剂盒(Takara,Dalian)克隆IHNV-Sn1203 基因组序列的5’和3’末端序列。克隆所得产物经0.8%琼脂糖凝胶电泳检测后,利用凝胶回收试剂盒(Omega)纯化回收目的片段。胶回收产物连接至pMD18-T simple 载体后,送吉林省库美生物科技有限公司进行测序。
1.4 IHNV-Sn1203 全基因组序列分析及系统进化分析
IHNV-Sn1203 基因组分段克隆片段中,每个片段至少随机选择3 个阳性克隆进行测序。测序结果采用SeqBuilder 软件进行正向序列和反向序列的装配验证。利用Lasergene、MEGA 5.0 软件,采用ClustalW 多重比对法和邻位相连法(Neighbor-Joining method),分析IHNV-Sn 1203 株的全基因组序列、基因组编码蛋白、基因组末端和未翻译序列、以及病毒基因同源性和系统发育,并构建系统进化树。
2 结果与分析
2.1 IHNV-Sn1203 株的全基因组信息
IHNV-Sn1203 株的基因组全长11131 nts,共编码6 个病毒蛋白,依次为核蛋白(N)、磷蛋白(P)、基质蛋白(M)、表面糖蛋白(G)、非结构蛋白(NV)和聚合酶蛋白(L),其基因组结构与已经发表的弹状病毒及IHNV 病毒基因组结构组成相似。表2 列出了IHNV-Sn1203 株的基因组特征及预测蛋白信息,IHNV-Sn1203 的所有蛋白编码基因均被称为基因连接区的非翻译序列(UTR)分隔开(图1)。
2.2 IHNV-Sn1203 株的6 个基因分析
IHNV-Sn1203 株编码的六个基因及其组成如表2 所示。其中,N 基因的开放阅读框全长1176nts,位于基因组的175~1350nts 之间,包含391 个氨基酸残基,编码蛋白大小约为42.36kDa。N 基因的两端分别含有112nts(63~174) 的5' 非翻译区,及80nts(1351~1430)的3' 非翻译区(表2)。N 基因转录起始位点位于保守转录起始序列(CGUG,63~66nts)的下游,转录终止序列位于1415~1430nts处,其中包含推测的多腺化信号(UCUUUUUUU)。
P 基因的开放阅读框全长693nts,位于基因组的1466~2158nts 之间,包含230 个氨基酸残基,编码蛋白大小约为25.99kDa。P 基因的两端分别含有33nts(1433~1465)的5' 非翻译区,及41nts(2159~2199)的3' 非翻译区。P 基因转录起始序列位于1433~1436nts,转录终止序列位于2184~2199nts 处。
M 基因的开放阅读框全长588nts,位于基因组的2255~2842nts 之间,包含195 个氨基酸残基,编码蛋白大小约为21.86kDa。M基因的两端分别含有53nts(2202~2254)的5' 非翻译区和103nts(2843~2945)的3' 非翻译区。M 基因转录起始序列位于2202~2205nts,转录终止序列位于2930~2945nts。

表2 IHNV-Sn1203 株的基因组特征和蛋白预测Tab.2 Genomic characteristics and protein prediction of IHNV-Sn1203
G 基因的开放阅读框全长1527nts,位于基因组的2948~2998nts 之间,包含508 个氨基酸残基,编码蛋白大小约为56.37kDa。G 基因的两端分别含有51nts(2948~2998)的5' 非翻译区和42nts(4526~4567)的3' 非翻译区。G 基因转录起始序列位于4552~4567nts,转录终止序列位于2930~2945nts 处。
NV 基因的开放阅读框全长336nts,位于基因组的4596~4931nts 之间,包含111 个氨基酸残基,编码蛋白大小约为13.30kDa。NV 基因的两端分别含有26nts(4570~4595) 的5' 非翻译区和7nts(4932~4938)的3'非翻译区。NV 基因转录起始序列位于4570~4573nts,转录终止序列位于4923~4938nts处,与NV 基因的开放阅读框有9nts(4923~4931)重叠。
L 基因的开放阅读框全长5961nts,位于基因组的5016~10976nts 之间,包含1986 个氨基酸残基,编码蛋白大小约为224.73kDa。L 基因的两端分别含有75nts(4941~5015) 的5' 非翻译区和54nts(10977~11030)的3' 非翻译区。L 基因转录起始序列位于 4941~4944nts,转录终止序列位于11015~11030nts 处。
2.3 基因组末端和未翻译序列
为了实现病毒蛋白的最佳翻译,弹状病毒的开放阅读框之间都含有非常保守的非翻译区域。这些区域通常以一个假定的转录停止/聚腺苷酸化基序[UCUA/GUCU7]开始,通过U 序列的重复复制向mRNA 加入poly(A)尾巴。这一序列后紧连着一个非转录的基因间二核苷酸AC 或GC 和一个保守的假定转录起始信号CGUG。图2-A 为IHNV-Sn1203株基因间的保守UTR 结构,各基因间均含有保守的基因终止序列(Geneend,GE)、基因间序列(Intergenicregions,IG)和基因起始序列(Genestar,GS)。IHNV-Sn1203 基因组的6 个基因间的基因连接长度不同,分别为:N-P (115nts)、P-M(96nts)、M-G(156nts)、G-NV(70nts)和NV-L(84nts)。研究发现,NV 基因的终止密码子重叠在其非翻译区的GE 中,这一结果未发于其他病毒,且其具体的作用也未阐明。IHNV-Sn1203 基因组的3'leader 序列长60nt,A/T 含量为68.3%,5'trailer 序列长99nt,A/T 含量为56.6%。同其他弹状病毒类似,IHNV-Sn1203 基因组RNA 的3'端,前16 个核苷酸均与5'端核苷酸互补(如图2-B 所示),而已报道的病毒株220-90 和Ch20101008 中分别有15 个和13 个互补,这种基因组末端的互补性质对弹状病毒的复制至关重要。
已有报道表明,Kozak 序列对保证真核生物基因的转录和翻译效率至关重要,特别是起始密码前-3 位的A 碱基[18]。本文分析了IHNV-Sn1203 基因组中的Kozak 序列(图3)。结果发现:IHNV-Sn1203株6 个病毒基因的起始密码子前面均含有严格的Kozak 序列,其-3 位碱基均为A。这一结果与已报道的IHNV 220-90 株的N 基因不同(-3 位碱基为G)。
2.4 同源性和系统发育分析
2.4.1 全基因组进化分析
本研究首先分析了IHNV 同其他弹状病毒全基因组的进化关系,结果显示IHNV 与HIRRV(Hirame rhabdovirus,HIRRV)进化关系最近,其次为SHRV(Snakehead rhabdovirus,SHRV)、VHSV(Viral hemorrhagic septicemia virus,VHSV)和SVCV(Spring viraemia of carp virus,SVCV)。IHNV-Sn1203 株同Ch20101008、BJSY 和HLJ-09 聚为一簇(图4),其核苷酸同源性分别为99.67%、99.52%和99.76%;同BjLL、220-90、WRAC、lambda ZAPII 聚为一簇,同Sn1203 株的核苷酸同源性分别为96.34%、94.89%、95.41%和95.88%。(表3)
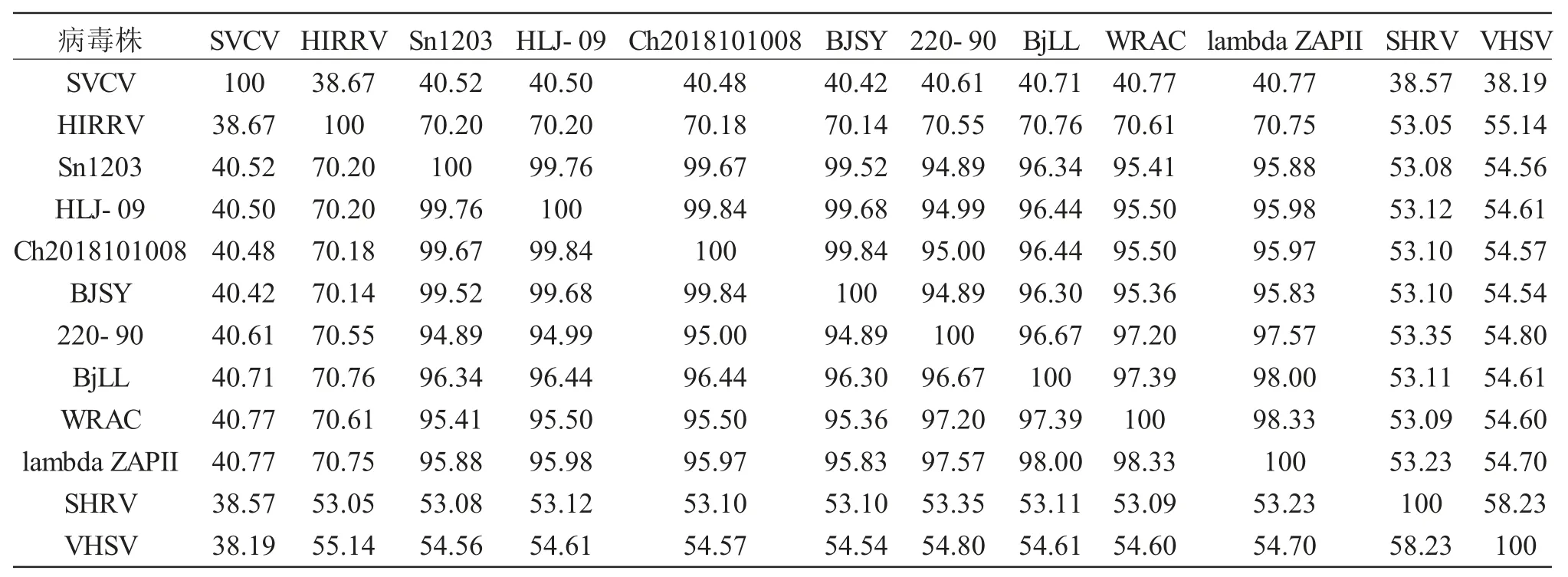
表3 IHNV-Sn1203 基因组序列同源性分析(%)Tab.3 Homology analysis of IHNV-Sn1203 genome sequence(%)
2.4.2 Sn1203 株病毒糖蛋白(Glycoprotein,G)基因进化分析
IHNV 病毒糖蛋白G 基因序列的变异系数小,又与病毒的毒力强弱直接相关,目前IHNV 的基因分型及进化分析多以G 蛋白序列为主。本研究对包括IHNV-Sn1203 株在内的100 株IHNV 的G 基因进行了系统进化分析。结果表明:IHNV 主要分为5个基因型,即E、U、M、L 和J,其中J 基因型又包含J Nagano 和J Shizuoka 两个基因亚型。系统进化分析结果表明,Sn1203 株同我国已报道的SD-12、YN-13、CJ-13、Ch20101008 和HLJ-09 等聚为一簇,共同归属于J Nagano 基因亚型,并且Sn1203 株与CJ-13 株的亲缘关系较近(图5)。
2.4.3 Sn1203 株各病毒基因进化分析
本研究同时利用GenBank 中IHNV 病毒株N基因、P 基因的全序列及L 基因、M基因和NV 基因的部分序列构建了系统进化树,如图6 所示。N 基因全序列的系统进化分析发现,Sn1203 株同LQN131107、CJ-13、zyx、HLJ-09 和Ch20101008 聚为一簇(图6-A),核苷酸同源性分别为99.32%、99.49%、99.66%、99.83%和99.83%;而中国分离株BjLL 则与美国株RB-76 及Round Butte 1 等聚为一簇,与Sn1203 株的核苷酸同源性为95.75%。P 基因全序列的系统进化分析发现,Sn1203 株同CJ-43、HLJ-09、Ch20101008 和BJSY 聚为一簇(图6-B),与HLJ-09 亲缘关系最近,其核苷酸同源性为99.86%,与BjLL 中国分离株亲缘关系最远,核苷酸同源性为94.37%。L 基因部分序列的系统进化分析发现,Sn1203 株同HLJ-09、Ch20101008 和BJSY 聚为一簇(图6-C),与HLJ-09 亲缘关系最近,其核苷酸同源性为99.80%,与BjLL 中国分离株亲缘关系最远,核苷酸同源性为97.30%。M基因及NV 基因部分序列的系统进化分析发现,Sn1203 株同CJ-13、HLJ-09、Ch20101008 和BJSY 聚为一簇(图6-D、图6-E),与HLJ-09 及Ch20101008 亲缘关系最近,M 基因核苷酸同源性均为99.49%,NV 基因核苷酸同源性分别为99.70%和99.40%,与BjLL 中国分离株亲缘关系最远。以上结果表明:Sn1203 株与HLJ-09 株及Ch20101008 的亲缘关系最近,而与中国分离株BjLL 亲缘关系最远。
3 讨论
1985年我国东北部某虹鳟鱼养殖场首次暴发了IHN 疫情,次年IHN 暴发15d 就导致该养殖场60 万尾虹鳟死亡,从此IHN 成为严重威胁我国鲑鳟养殖的主要疫病[12,19]。2012年本课题组分离得到IHNV-Sn1203 病毒株,以该病毒株为对象开展了大量的研究工作,包括疫苗、致病机理及免疫学研究等[1,13,20,21],但未详细分析IHNV-Sn1203 病毒株的基因组成、分子生物学特征等。为更清楚地了解该病毒株的分子生物学特性,本研究克隆了IHNV-Sn1203 病毒株的全基因组序列,分析了其全基因组序列、基因组编码蛋白、基因组末端和未翻译序列、以及病毒基因同源性和系统发育。
IHNV-Sn1203 病毒株全基因组长度为11131 nts,与其他诺拉弹状病毒属病毒的比对发现,在其基因组的3' 末端和5' 末端均含有与其他诺拉弹状病毒属病毒相同的保守序列:3' 末端为CAUAU,5'末端为GUAUA。在其基因组的3'末端的前16 个核苷酸均与5' 末端核苷酸互补,与已报道的220-90病毒株及Ch20101008 病毒株不同(220-90 有15 个互补,Ch20101008 有13 个互补)。所有诺拉弹状病毒属病毒基因组3' 末端的核苷酸长度基本保持一致(51~60nts),但5' 末端核苷酸长度却呈高度可变性(42~116nts)。已有的研究认为,病毒基因组3'末端和5' 末端核苷酸序列的互补性,及5' 末端均具有一定的功能意义,可能与病毒的复制密切相关,但具体机制尚不清楚[22]。对IHNV-Sn1203 病毒株各蛋白编码基因前后序列的分析发现,所有蛋白编码基因前均含有一段长度为4 个核苷酸的保守基因启动序列(CGUG,GS),蛋白编码基因后均含有一段长度为16 个核苷酸的保守基因终止序列(G/UGG/UUCUA/GUCU7,GE),所有的基因均以7个尿嘧啶(U)残基引入聚腺苷酸化信号的方式终止。在7 个尿嘧啶残基后,有2 个不参加转录的核苷酸(A/GC,IG),作为两个基因间的间隔序列。聚合酶跳过这两个核苷酸序列,进入下一个基因启动序列,开始转录下一个基因。本研究发现,IHNV 病毒株的6 个基因中,N、P、M、G 和L 基因前后均含有完整的基因启动序列和基因终止序列,而NV 基因后的基因终止序列与其编码序列有9nts(4923~4931)重叠(表2),目前关于这种序列重叠现象的功能仍未有相关报道。
对IHNV-Sn1203 病毒株全基因组系统进化分析发现,除BjLL 外,中国毒株Ch20101008、BJSY、HLJ-09 和Sn1203 均聚为一簇。IHNV-Sn1203 病毒株N 基因、P 基因的全序列及L 基因、M基因和NV基因的部分序列系统进化分析表明,Sn1203 株与中国株HLJ-09 亲缘关系最近,各核苷酸的同源性依次为99.83%、99.86%、99.80%、99.49%和99.70%。
G 基因作为病毒表面主要抗原蛋白基因,与病毒的毒力强弱直接相关[23]。目前IHNV 的基因分型及进化分析多以G 基因序列为主,本研究分析了G基因的系统进化发现,现有的IHNV 病毒株主要分为5 个基因型,即E、U、M、L、J,其中J 基因型又包含J Nagano 和J Shizuoka 两个基因亚型。其中广泛研究的毒株220-90 和WRAC 属于M基因型,Baker Lake 94 属于U 基因型,strain K 属于E 基因型,LR-73、Col-85 属于L 基因型。Sn1203 株同我国已报道的 SD-12、YN-13、CJ-13、Ch20101008 和HLJ-09 等聚为一簇,共同归属于J Nagano 基因亚型;Sn1203 株与CJ-13 株的亲缘关系较近。由G 基因的进化分析发现,J 基因型的病毒与U 基因型亲缘关系最近,猜测IHNV 病毒最初可能是通过进口鱼卵的方式由U 基因型引入,随时间的推移逐渐进化为J 基因型。因此,对进口鱼卵的监控是做好外源病毒防控的关键手段。
综上所述,本研究结果表明:Sn1203 株同我国已报道的SD-12、YN-13、CJ-13、Ch20101008 和HLJ-09 病毒株等聚为一簇,共同归属于J 基因型中的Nagano 基因亚型。本研究可为我国IHNV 病毒株的分子生物学特性研究及分子进化研究提供重要参考。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
今日农业(2021年11期)2021-08-13 08:53:24
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
遗传(2014年3期)2014-02-28 20:58:49
世界科学(2014年8期)2014-02-28 14:58:31
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
世界科学(2013年6期)2013-03-11 18:09:33
