
铅胁迫下柳树4个无性系生长和生理特性的变化
2020-06-18 08:33:12黄瑞芳教忠意施士争
江苏林业科技 2020年2期
黄瑞芳,张 忠,曹 瑶,教忠意,施士争*
(1.江苏省林业科学研究院,江苏 南京 211153; 2.江苏省农业种质资源保护与利用平台,江苏 南京 210014;3.沭阳县生产力促进中心,江苏 沭阳 223600)
蓄电池的制造以及其他涉铅工矿企业的发展,由于工艺、设备落后,管理不善等原因,造成了较重的铅污染。而铅污染不能被生物代谢所分解,在环境中滞留的时间较长,积累到一定程度就会对植物造成毒害,且能通过食物链在动物和人体内富集,对人体的肾脏、血液、神经系统等危害极大[1-2]。近年来,越来越多的研究用近超积累植物或速生、生物量大的植物来进行铅污染的修复。柳树,尤其是灌木柳,其根系发达,适应性强,耐水湿,生物量大,生长速度快,用作生物能源,不直接与食物链相联系。与草本相比,因其高积累性而金属污染修复能力强[3],有研究甚至认为柳树是重金属的超积累植物[4-5]。因此柳树是土壤污染生物修复的理想材料[6],适合用于修复低、中浓度重金属污染[7]。
植物的生长和各生理指标的变化是植物在受到铅胁迫时的自我防御机制,是判断其植物对铅胁迫耐受性的依据之一。植物的生物量是其受生长环境影响后最直观、最全面的反应,是耐铅性的重要评价因子之一[8]。生物量越大,植物体的盖度越大,吸收的重金属量也越多,修复受污染土壤的效果越好[9]。超氧化物歧化酶(Superoxide dismutase,SOD)和过氧化物酶(Peroxidase,POD)是是植物抗氧化酶系统中的重要组成成员,对机体的氧化与抗氧化平衡起着至关重要的作用,可以清除超氧阴离子自由基,抑制膜脂过氧化,在植物的抗逆过程中扮演着重要角色[10-11]。丙二醛(malonic dialdehyde,MDA)是细胞膜质过氧化的产物,其含量能够反映出机体内脂质过氧化的程度,可作为细胞损伤的指标之一。可溶性蛋白是重要的渗透调节物质和营养物质,其含量与细胞内溶质的渗透势以及原生质体内外的渗透度等都有很密切的关系,经常用作筛选抗性的指标之一。
本文研究了不同铅胁迫浓度下柳树4个无性系的生物量、SOD酶活性、POD酶活性、MDA含量和可溶性蛋白含量的变化,旨在了解铅胁迫对柳树的生长和生理特性的影响,了解柳树对铅胁迫的生理适应机制,为进一步筛选耐铅柳树品种提供理论依据。
1 材料与方法
1.1 试验材料
杞柳(Salixintegra)、簸箕柳(S.suchowensis)的4个无性系(见表1),来自于江苏省林业科学研究院国家柳树良种基地柳树种质资源圃。

表1 供试无性系
1.2 试验方法
采取水培法。将挑选的柳树苗置于定植篮中,每篮10株。营养液为改良Hogland。水培瓶200 mL,插穗3株/瓶,Pb2+以Pb(NO3)2的形式加入,处理为0(CK),20(处理1),40(处理2),80 mg/L(处理3)[12]。
1.3 测定方法
胁迫实验结束后收获植株,进行超氧化物歧化酶(SOD)和过氧化物酶(POD)的活性测定,以及丙二醛(MDA)和可溶性蛋白质含量的测定。所有生理指标均采用南京建成生物工程研究所的试剂盒进行测定。SOD酶活性采用羟胺法测定(试剂盒编号:A001-1-1),SOD酶活性采用比色法进行测定(试剂盒编号:A084-3),MDA含量采用TBA法进行测定(试剂盒编号:A003-1-1),可溶性蛋白含量采用考马斯亮蓝法进行测定(试剂盒编号:A045-2),柳树各无性系的耐铅能力选择加权隶属函数法进行计算。得到的数据利用Excel 2010和IBM SPSS Statistics 19进行分析。其中隶属函数的计算方法如下:
U(Xi)=(Xi-Xmin)/(Xmax-Xmin)
(1)
U(Xi)=1-(Xi-Xmin) /(Xmax-Xmin)
(2)
Xi=∑Xij/n
(3)
其中,(1)用于某指标与重金属抗性呈正相关,(2)用于呈负相关。Xi为第i个指标的测定值,Xmax为所有参试无性系中该指标测定值中的最大值,Xmin为最小值。
2 结果与分析
2.1 不同质量浓度铅胁迫对无性系生物量的影响
从图1可以看出,随着铅胁迫质量浓度的增加,柳树4个无性系的生物量(g)呈降低趋势。CK(5.41±0.83)>处理1(3.19±0.75)>处理2(2.77±0.42)>处理3(2.46±0.34)。在铅胁迫质量浓度为20 mg/L时,4个无性系的生物量均极显著低于对照组(P<0.01),分别是对照的42.96%,55.17%,71.23%,69.92%,表明已经对柳树的生长产生了明显的胁迫效应。在铅胁迫质量浓度为80 mg/L时,4个柳树无性系的生物量高于对照50%的只有P63和P1024,表明高质量浓度的铅胁迫处理抑制生长的能力增强。
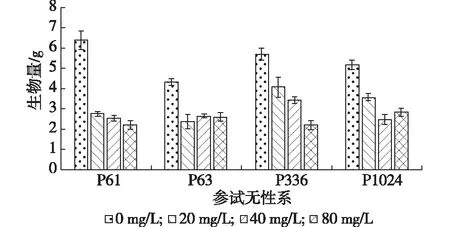
图1 不同铅胁迫质量浓度下参试无性系生物量的变化
2.2 不同质量浓度铅胁迫对无性系SOD酶活性的影响
从图2可以看出,随着铅胁迫质量浓度的增加,4个柳树无性系SOD酶活性(U/g)呈现增高的趋势。处理3(92.74±7.31)>处理2(87.95±7.57)>处理1(83.02±12.19)>CK(77.18±9.46)。在铅处理质量浓度为80 mg/L时,SOD活性最高,分别比对照增加了18.99%,12.99%,43.08%,11.75%。
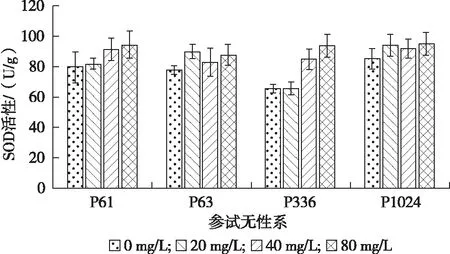
图2 不同铅胁迫质量浓度下参试无性系SOD酶活性的变化
2.3 不同质量浓度铅胁迫对无性系POD酶活性的影响
由图3可看出,随着铅胁迫质量浓度的增加,4个柳树无性系POD酶活性(U/g)均呈现先增加后减低的趋势。处理2(196.99±63.98)>处理1(177.66±49.34)>处理3(169.24±72.41)>CK(111.06±25.04)。P61和P63在铅质量浓度为40 mg/L时,达到最高值,P61比对照高110.25%,P63比对照高191.67%。P336和P1024均在铅质量浓度为20 mg/L时达到最高,P336比对照高18.75%,P1024比对照高261.76%。在铅质量浓度达到80 mg/L时,P63比对照高155.20%,P1024比对照高99.04%,而P61与对照无显著差异。
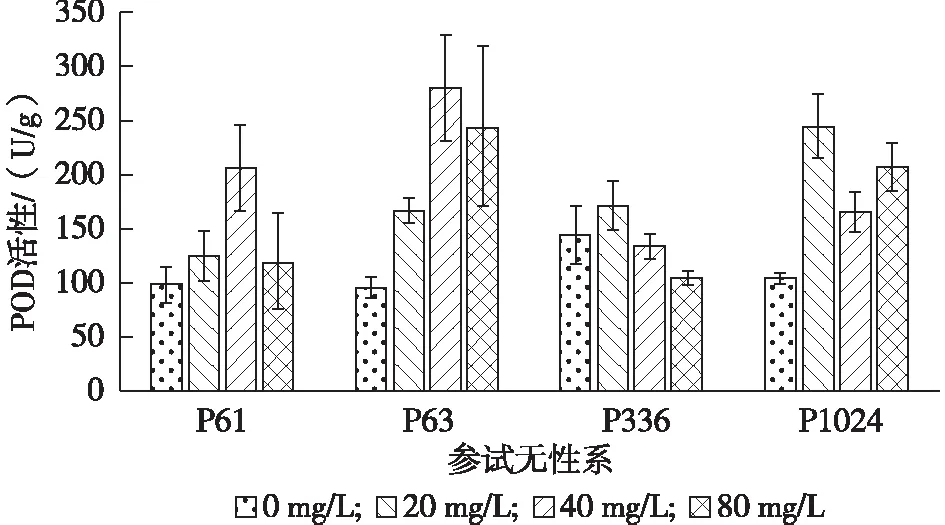
图3 不同铅胁迫质量浓度下参试无性系POD酶活性的变化
2.4 不同质量浓度铅胁迫对无性系MDA含量的影响
由图4可看出,随着铅胁迫质量浓度的增加,4个柳树无性系MDA含量(nmol/g)均呈整体升高趋势,且均显著高于对照。处理3(39.52±9.57)>处理2(24.58±5.29)>处理1(18.17±5.72)>CK(13.67±3.49)。在铅质量浓度达到80 mg/L时,P61,P63,P336,P1024分别比对照高206.52%,318.74%,174.38%,98.83%。
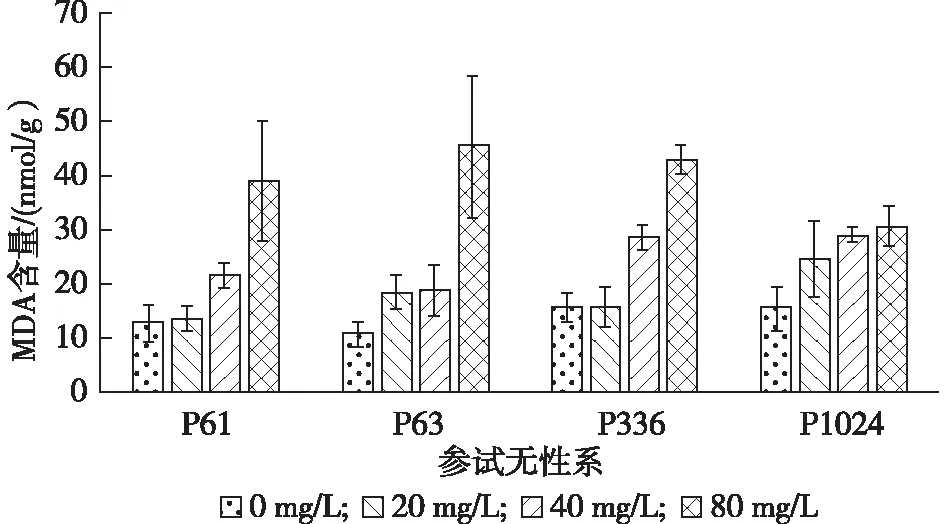
图4 不同铅胁迫质量浓度下参试无性系DA含量的变化
2.5 不同质量浓度铅胁迫对无性系可溶性蛋白含量的影响
由图5可看出,4个柳树无性系可溶性蛋白含量(mg/kg)随着铅胁迫质量浓度的增加均呈现下降的趋势。在质量铅浓度达到80 mg/L时,P61,P63,P336,P1024分别是对照的62.26%,58.61%,79.98%,61.88%,P63的变化最大,P336的变化最小。
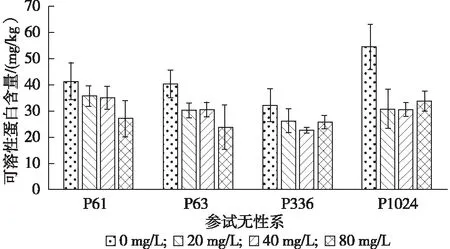
图5 不同铅胁迫质量浓度下参试无性系可溶性蛋白含量的变化
2.6 柳树各无性系耐铅能力比较
对植物的抗逆性状进行综合评价时,加权隶属函数法是常用也是较为认可的综合评价方法[13]。一般采用主成分分析法来筛选综合指标。再依据各综合指标的贡献率来确定各自的权重,进而利用加权隶属函数法算出每个参试无性系的抗逆性综合评价值(D值),据此对参试无性系的抗逆性进行排序。
对参试无性系的5个指标进行了主成分分析,共提取了2个主成分,第1主成分贡献率为47.89%,第2主成分贡献率为23.49%。其中主成分1=-0.77×生物量+0.17×SOD酶活性+0.59×POD酶活性+0.85×MDA含量-0.84×可溶性蛋白含量;主成分2=0.10×生物量+0.91×SOD酶活性-0.50×POD酶活性+0.31×MDA含量+0.05×可溶性蛋白含量;综合主成分=-0.34×生物量+0.29×SOD酶活性+0.17×POD酶活性+0.48×MDA含量-0.39×可溶性蛋白含量。
根据主成分分析确定的各指标贡献值进行加权隶属函数分析,根据式(1)(2)(3)得出各无性系耐铅性的综合评价值,结果(见表2)表明4个无性系耐铅能力的强弱顺序为P336>P63>P1024>P61。

表2 参试无性系可各指标的隶属函数值比较
3 讨论与结论
铅是植物的非必需元素,植物在受到铅胁迫时,其生长受到抑制,表现为植株矮小,生物量下降,甚至出现死亡。本研究中随着铅胁迫浓度的增加,柳树无性系的生物量呈降低趋势,这是因为铅胁迫抑制了柳树的生长。P61在铅质量浓度为20 mg/L时生物量下降程度最高,可能说明P61的生长受到铅胁迫的抑制较为明显。
植物在正常生长条件下,其体内活性氧的产生和清除处于动态平衡,而在逆境胁迫下,平衡被打破,体内积累大量的活性氧,从而使植物细胞受到氧化胁迫,超氧化物歧化酶(SOD)、过氧化物酶(POD)为抗氧化防御系统的重要组成成员,可清除活性氧[14-15]。本研究中随着铅胁迫质量浓度的增加,柳树无性系的SOD酶活性呈增高趋势,而POD酶活性呈先增高后下降的趋势,这可能是因为铅胁迫刺激导致体内活性氧的动态平衡遭到破坏,从而导致抗氧化酶活性发生改变[16-17],从而提高植物的抗逆能力[18]。P336的SOD酶活性的上升程度显著高于其他3个无性系,说明P336的SOD酶对铅胁迫的敏感程度较其他3个无性系高。而P1024在铅质量浓度为20 mg/L时极显著增高,达到最高值,这可能说明P1024的POD酶对铅胁迫较为敏感。
生物体内的自由基作用于脂质发生过氧化反应,产生丙二醛,可使蛋白质、核酸等交联聚合,产生细胞毒性。若丙二醛过多,会引发膜脂过氧化[19],因此MDA含量能间接反映植物的衰老生理和抗性生理。本研究中柳树在受到铅胁迫后,MDA含量显著上升,这说明柳树在受到铅胁迫以后导致了膜脂过氧化,植物受到损伤。P1024在铅质量浓度为20 mg/L时MDA含量即显著增加,可能是因为P1024的MDA对铅胁迫较为敏感。值得一提的是,P1024在铅胁迫3个质量浓度处理下的MDA的含量并无显著差异,这可能说明随着铅胁迫质量浓度的有限增加,P1024受损伤的程度并无增加,而其余3个无性系都有显著增加。
植物在逆境下会主动进行渗透调节,积累各种有机或无机物质来降低渗透势,从而抵抗逆境胁迫。但不同植物其溶质的累积程度也不同,主要是由于他们的基因表达不同,而基因表达的产物——蛋白质也会不同,因此可溶性蛋白也是植物抗性指标之一[20]。本研究中,随着铅胁迫质量浓度的增加,柳树无性系的可溶性蛋白含量呈下降趋势。P1024的下降程度显著高于其他3个无性系,且P1024在铅质量浓度为20 mg/L时可溶性蛋白即显著降低,而4个柳树无性系在铅胁迫3个质量浓度处理下的可溶性蛋白含量并无显著差异,这可能说明随着铅胁迫质量浓度的有限增加,铅进入细胞后对蛋白水解酶的活性以及蛋白合成相关酶的毒害和钝化作用并无显著增加。
综上所述,本研究中设计的铅质量浓度严重抑制了柳树的生长和生物量,并且破坏了体内抗氧化系统,造成了植物细胞膜损伤,渗透调节能力降低,对柳树的生理活动产生了严重的胁迫效应。
植物对重金属的耐受性与许多细胞活动都有关,如金属离子的螯合、运输、富集等[21-22],虽然利用生理生化指标可以一定程度上评价植物的耐铅性,相对缩短林木耐重金属新品种的选育过程,但单纯的用生理指标进行植物的耐铅性评价还不够,要想全面反映植物的耐铅性,还需综合考虑各项指标。
猜你喜欢
今日农业(2020年19期)2020-12-14 14:16:52
中成药(2018年8期)2018-08-29 01:28:34
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
少儿科学周刊·儿童版(2017年5期)2017-06-29 22:14:38
中国科技信息(2015年2期)2015-11-16 08:18:32
医学研究杂志(2015年9期)2015-07-01 17:28:24
中国当代医药(2015年9期)2015-03-01 02:02:13
小天使·一年级语数英综合(2014年3期)2014-03-15 01:11:21
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
食品科学(2013年24期)2013-03-11 18:30:25
