
贝加尔针茅草原土壤原位矿化过程中碳氮转化耦合特征
2020-06-10 03:26刘红梅张海芳赵建宁杨殿林
干旱地区农业研究 2020年2期
刘红梅,张海芳,秦 洁,赵建宁,王 慧,杨殿林
(农业农村部环境保护科研监测所,农业农村部产地环境污染防控重点实验室,天津市农业环境与农产品安全重点实验室,天津300191)
草原生态系统的碳循环和氮循环是草原生态系统最重要的物质循环过程,调节和维持着生态系统生产力与稳定性。草原生态系统是陆地生态系统重要的土壤碳库和氮库,研究其碳氮转化过程对全球变化的响应非常重要。土壤碳氮含量及其动态平衡直接关系到土壤中养分元素的释放与供应,影响着土壤肥力和草原生产力。氮沉降增加作为全球变化的重要现象之一[1],其对草原生态系统碳氮循环的影响和反馈存在诸多不确定性。研究土壤碳氮转化特征对氮沉降增加的响应是揭示这些不确定性,提高草原土壤碳积累,减少氮素损失的科学基础。在氮沉降增加的背景下,对微生物驱动的碳、氮元素过程耦合机制的研究,将有利于认识土壤微生物在养分循环中的作用,对减少氮沉降对草原生态系统的危害具有重要意义。
在大气氮沉降增加的背景下,土壤碳氮含量、碳氮转化速率与温室气体排放密切相关。高氮沉降造成陆地生态系统土壤中活性氮含量的增加,从而增加土壤的硝化作用和反硝化作用,增加N2O排放[2-3]。有研究表明,氮素添加会促进土壤呼吸速率,降低土壤碳储量[4]。土壤有机碳(Soil Organic Carbon,SOC)含量较低时,SOC矿化受碳素限制,而当SOC含量和C/N比值较高时,SOC矿化受氮素限制[5]。Hyvönen 等[6]对北欧的 15 个森林长期氮添加定位试验表明,增加氮素输入降低了SOC矿化,有利于SOC积累。张璐等[7]对内蒙古羊草草原不同氮添加处理土壤的室内培养试验研究表明,氮添加显著改变土壤累积氮矿化量,低氮添加累积氮矿化量最高,中等氮添加净矿化作用最高。罗亲普等[8]对内蒙古温带典型草原原位培养试验表明,高氮处理促进净硝化速率、氨化速率和矿化速率。吕玉等[9]研究认为,在一定施氮范围内,随着施氮量增加,土壤硝化势和土壤硝化速率增加,而高氮处理下降低。土壤碳氮的转化受土壤碳氮比值的控制[10],在氮素充足的条件下,较高的植物残体投入和较低的碳氮比导致土壤微生物更倾向于利用新的碳源底物,减少原有有机质的矿化[11];而当氮素受限制条件下,高的植物残体投入和较高的碳氮比会增加土壤微生物对氮素的需求,从而促进土壤原有机质的分解[12]。土壤碳矿化和氮矿化的关系大都研究认为是正相关关系,微生物分解有机质释放CO2的同时释放无机氮。但Song等[13]对青藏高原高山草甸研究表明,在土壤碳、氮元素都为缺乏情况下,土壤碳矿化和氮矿化为负相关关系,认为只有有机物释放的无机氮满足微生物需要之后才会释放到土壤中。
贝加尔针茅草原是我国温带草甸草原的代表性类型之一,在我国畜牧业生产中占有重要地位。被高强度用于放牧的贝加尔针茅天然草原,牧草生长所需要的氮素由土壤微生物矿化有机氮来满足[14]。在氮沉降增加条件下,贝加尔针茅草原土壤碳氮转化速率发生怎样的变化,以及这些变化对土壤碳氮积累产生怎样的影响尚缺乏明确的认识。为此,本研究以贝加尔针茅草原为研究对象,通过模拟氮沉降增加的氮添加长期试验,运用PVC顶盖埋管法进行原位矿化培养试验,研究SOC、可溶性有机碳(dissolved organic carbon,DOC)和微生物生物量碳(microbial biomass carbon,MBC)与土壤有机氮(soil organic nitrogen,SON)、无机氮以及净氨化速率、净硝化速率、净矿化速率的变化特征以及碳氮耦合关系。以期揭示贝加尔针茅草原土壤碳氮转化对未来全球变化可能的响应,为制定温带草甸草原应对气候变化的策略提供科学依据。
1 材料与方法
1.1 研究区域概况
研究区位于内蒙古自治区鄂温克自治旗伊敏苏木境内,地理位置为北纬 48°27'~48°35',东经119°35'~119°41'。半干旱大陆性季风气候,处于温带草甸草原区,主要植被类型为贝加尔针茅(Stipa baicalensis)和羊草(Leymus chinensis)。土壤类型为暗栗钙土。试验开始时土壤基础理化性质为:土壤pH值7.07,总有机碳 27.92 g·kg-1,全氮 1.85 g·kg-1,全磷 0.45 g·kg-1。
1.2 样地设置与原位培养试验
样地设置开始于2010年6月,氮素添加处理强度和频度参考国际上同类研究的处理方法[15]。氮素添加处理设8个水平依次为:对照N0,低氮添加(15、30、50 kg·hm-2·a-1)分别记为 N15、N30 和N50,高氮添加(100、150、200、300 kg·hm-2·a-1)分别记为 N100、N150、N200和 N300。氮素为NH4NO3,每年分2次施入,分别于6月中旬和7月中旬施入。水溶后均匀喷施到小区内,对照小区喷洒相同量的水。共8个处理,4次重复,小区面积8 m×8 m,小区间设2 m隔离带,重复间设5 m隔离带。
采用PVC顶盖埋管原位培养法,于2015年8月中旬,用力将PVC矿化管(长12 cm,内径5 cm)砸入土壤,直到管上端与地面相平,将PVC管取出,剥离底部2 cm土壤,顶部用透气不透水的塑料薄膜封口,下端用脱脂棉和纱布封口后放回原处培养。在每个处理小区各埋入矿化管18根。同时在每个处理小区用土钻取0~10 cm土壤样品3钻,混匀装入1个自封袋,用冰盒带回实验室,测定土壤硝态氮(nitrage nitrogen,)、铵 态 氮 (ammonia nitrogen,)含量,作为氮转化培养的初始值。在各个处理小区内于2016年6月(培养300 d)、7月(培养330 d)、8月(培养 360 d)、9月(培养 390 d)中旬分别取出3根矿化管,去除管中的根系,用冰盒带回实验室,测定SOC、DOC、MBC、土壤微生物生物量氮(Microbial Biomass Nitrogen,MBN)、N和含量,计算碳氮矿化速率。
1.3 测定方法
1.4 数据统计分析
采用Excel 2010进行作图分析,对土壤碳氮相关指标作线性回归分析。SPSS 16.0统计软件对8个不同氮添加水平同一培养时间和同一氮添加水平不同培养时间土壤理化指标分别进行单因素方差分析(one-way ANOVA)。碳氮组分与碳氮转化速率进行pearson相关性分析。
净氨化速率(mg·g-1·d-1)=(培养后-培养前)/天数
净硝化速率(mg·g-1·d-1)=(培养后-培养前/天数
净矿化速率(mg·g-1·d-1)=[培养后(+)-培养前/天数
SOC、SON、MBC和 MBN的转化速率与上述计算相同,即培养后对应含量减去培养前含量,再除以培养天数。
2 结果与分析
2.1 土壤原位矿化过程中碳素的变化特征
2.1.1 SOC、土壤DOC和土壤MBC含量变化 各个氮处理在矿化期间SOC、DOC和MBC含量变化见表1。N0,N15,N30和N50在整个矿化期间变化不显著。N300处理在培养初期SOC含量最高,显著高于其他矿化培养时间。在培养第300 d时,各个氮添加处理SOC含量无显著差异。培养第330 d时,高氮添加(N100、N150、N200和 N300)SOC含量均显著高于低氮添加(N15,N30和N50)和对照N0。各个氮处理在整个矿化期间,土壤DOC含量总体表现为先升高后降低的趋势,在培养360 d时最高。培养0 d,7个氮添加处理土壤DOC均显著高于对照。培养300 d时,低氮添加(N15、N30和N50)高于高氮添加(N100、N150、N200和 N300)和对照。培养 360 d时,高氮添加(N100、N150、N200和N300)土壤DOC含量均显著高于低氮添加(N15、N30和N50)和对照,N100处理土壤DOC含量最高。在整个矿化培养期内,7个氮添加处理土壤MBC平均含量均显著低于对照。在培养360 d时,N200、N150、N100处理土壤 MBC含量显著高于N15、N30、N50和N300,而与对照相比无显著差异。在培养390 d时,高氮添加(N150、N200和N300)土壤MBC含量显著低于低氮添加(N15、N30和N50)和对照。
2.1.2 SOC和土壤MBC转化速率变化 矿化期间的SOC转化速率见图1。在矿化培养期内,N15与N30处理的SOC转化速率无显著差异。N30处理在整个矿化期间,SOC转化速率均为正值。N0和N50处理在培养360 d与N100、N150和N200处理在培养330 d的SOC转化速率为正值。高氮添加(N100、N150、N200和 N300)在培养第 330 d时,SOC转化速率均高于其他培养时间。在培养第360 d和 390 d时,高氮添加(N100、N150、N200和N300)的SOC转化速率显著低于低氮添加(N15、N30和N50)和对照。培养期间,N15和N30的有机碳转化速率高于对照N0,N150、N200和N300处理的有机碳的转化速率低于对照N0。

表1 土壤原位矿化过程中SOC、DOC和MBC含量变化Table 1 Changes of SOC,DOC and MBC contents in soil during in-situ mineralization

图1 SOC转化速率Fig.1 Dynamics of SOCconversion rate
矿化期间的土壤MBC转化速率见图2。N0、N50、N100、N150、N200 和 N300在培养第 360 d 时,土壤MBC转化速率均显著高于其他培养时间。在培养 360 d时,高氮添加(N100、N150、N200和N300)土壤MBC转化速率显著高于低氮添加(N15、N30和 N50)和对照。培养 390 d时,高氮添加(N150、N200和N300)低于或显著低于低氮添加和对照。培养300 d和培养330d时,N300处理土壤MBC转化速率均显著低于其他氮添加处理。
2.2 土壤原位矿化过程中氮素的动态变化特征
2.2.2 土壤无机氮转化速率变化 培养期内土壤净氨化速率变化见图3。在培养期间,土壤净氨化速率均为负值。培养期内,高氮添加(N100、N150、N200和N300)处理在培养360 d时土壤净氨化速率最高,N0、N15和N30时处理在培养300 d时最高。N50处理的净氨化速率在培养390 d时最高,但与培养330 d和360 d土壤的净氨化速率无显著差异。在培养300 d、330 d和390 d时,N15处理的土壤净氨化速率显著高于其他氮添加处理和对照。在培养360 d时,N150处理土壤净氨化速率显著高于其他氮添加处理和对照。培养期间,N30、N50和N100处理的土壤净氮化速率均显著低于对照N0,分别降低了16.88%、169.60%和150.67%。

图2 土壤MBC转化速率Fig.2 Dynamics of soil MBCconversion rate
表2 土壤原位矿化过程中含量变化/(mg·kg-1)Table 2 Changes of contents in soil during in-situ mineralization

表2 土壤原位矿化过程中含量变化/(mg·kg-1)Table 2 Changes of contents in soil during in-situ mineralization
培养时间Incubation time/d铵态氮NH+4-N N0 N15 N30 N50 N100 N150 N200 N300 0 27.42±1.28Ac 21.11±0.43Ad 30.97±2.24Ac 66.26±2.06Aa 61.16±3.88Ab 27.60±0.85Ac 29.25±1.05Ac 30.32±0.80Ac 300 11.40±0.69Bc 10.57±1.03Bc 11.35±0.98Bc 10.94±0.86Bc 9.06B±0.71Cd 11.80±0.75Cbc 13.22±1.11Bb 16.89±0.75Ba 330 3.85±0.20Cf 8.28±0.69Cc 5.00±0.43Ce 12.44±0.27Ba 6.54±0.11CDd 7.76±0.37Dc 5.91±0.75Dde 10.22±1.16Db 360 3.43±0.18Cd 2.93±0.28Dd 3.35±0.23Cd 6.65±1.32Cc 10.52±0.57Bb 13.5±1.43Ba 9.31±0.64Cb 14.19±0.67Ca 390 3.26±0.50Ccd 3.8±0.12Dbc 3.13±0.36Cd 3.50±0.15Dbcd 3.91±0.53Db 2.98±0.22Ed 3.56±0.17Ebcd 6.41±0.08Ea培养时间Incubation time/d硝态氮NO-3-N N0 N15 N30 N50 N100 N150 N200 N300 0 1.82±0.04Ee 1.92±0.23Ce 2.93±0.24Ee 2.82±0.07Ee 9.90±0.13Ed 37.67±0.48Dc 48.09±1.46Cb 66.21±0.98Ca 300 13.22±0.99Cg 17.48±0.56Be 22.48±3.06Ce 30.73±1.58Cd 36.89±0.89Cc 49.30±1.91Cb 50.72±0.63Cb 85.69±5.08Ba 330 16.34±0.61Bg 22.56±0.72Af 46.11±1.42Ae 52.29±4.59Bd 51.15±0.40Bd 77.90±1.00Ab 65.27±1.67Bc 114.14±3.61Aa 360 21.61±0.77Ag 23.11±0.19Ag 32.02±0.70Bf 61.85±0.42Ad 85.37±4.89Ab 56.44±0.86Be 80.20±1.92Ac 89.56±0.89Ba 390 6.93±0.53Df 16.70±1.39Be 18.00±0.73De 23.61±0.89Dcd 21.00±2.54Dd 25.81±1.61Ec 37.72±1.22Db 65.06±2.96Ca
随培养时间的延长,土壤净硝化速率表现为先升高后降低趋势(图4)。培养390 d时,8个氮处理土壤净硝化速率均为最低。培养390 d时,N150、N200和N300处理土壤净硝化速率为负值,其他时间为正值。在培养300 d、330 d和390 d时,N50处理土壤净硝化速率高于或显著高于其他氮添加处理和对照。在培养360 d时,N100处理土壤净硝化速率显著高于其他氮添加处理和对照。培养期间,N15、N30、N50和N100的土壤净硝化速率均显著高于对照N0,分别增加了40.80%、110.31%、206.83%和202.04%。

图3 培养期间土壤净氨化速率变化Fig.3 Dynamics of net ammonization rate

图4 培养期间土壤净硝化速率变化Fig.4 Dynamics of net nitrification rate

图5 培养期间土壤净矿化速率变化Fig.5 Dynamics of net mineralization rate
培养期间土壤净矿化速率见图 5。N0、N30、N50、N100、N150、N200 和 N300 处理在培养 360 d时净矿化速率最高,N15处理在培养300 d时土壤净矿化速率最高。培养390 d时,各处理土壤净矿化速率均为负值,且N15处理显著高于其他处理。培养360 d时,高氮添加(N100、N150、N200和N300)净矿化速率显著高于低氮添加(N15、N30和N50)和对照。N300处理在培养300 d和330 d时,土壤净矿化速率高于或显著高于其他氮处理。培养期间,N15和N30处理的净矿化速率高于对照N0,分别提高了 150%、50%;N50、N100、N150和 N200处理的净矿化速率低于对照N0,分别降低了254.52%、161.50%、33.90%和79.85%。
2.2.3 土壤有机氮含量变化 SON、土壤DON和土壤MBN含量变化见表3。N15、N30、N50和N100处理SON含量随培养时间延长呈升高趋势,培养390 d时最高。N0、N150、N200和N300的 SON含量随培养时间延长,呈现先升高后降低趋势。N0和N200在培养360 d时SON含量最高,N300在培养330 d时SON含量最高。在培养0 d和300 d时,7个氮添加处理SON含量与对照相比无显著差异。在培养330 d时,高氮添加(N150、N200和 N300)SON含量显著高于低氮添加(N15、N30和N50)和对照。培养390 d时,7个氮添加处理SON含量均高于对照。8个氮添加处理的土壤DON含量均在培养第300 d时最低。培养330 d、360 d和390 d时,高氮添加(N100、N150、N200和 N300)土壤DON含量显著高于低氮添加(N15、N30和N50)和对照。培养 300 d时,N150、N200和 N300处理的土壤DON 含量显著高于 N0、N15、N30、N50和 N100。培养0 d时,高氮添加(N100、N150、N200和 N300)土壤DON含量显著高于 N0、N15和 N50。N0、N15、N30和N50在培养初期土壤MBN含量最低,N100在培养330 d时土壤MBN含量最低,N150、N200和N300在培养390 d时土壤MBN含量最低。培养0 d时,高氮添加(N100、N150、N200和 N300)土壤 MBN含量显著高于低氮添加(N15、N30和N50)和对照。
2.2.4 SON含量及其转化速率变化 SON转化速率见图6。培养期内,各处理SON转化速率均为正值。N15、N30和N50处理 SON转化速率在培养390 d时最高。N100、N150、N200和 N300的 SON转化速率随培养时间延长呈先升高后降低趋势,N100、N150和 N300处理在培养 330 d时最高,N200在培养360 d时最高。培养330 d和培养360 d时,高氮添加(N100、N150、N200和 N300)SON 转化速率高于或显著高于低氮添加(N15、N30和N50)和对照。培养300 d时,N300处理的SON转化速率显著高于其他氮处理。培养390 d时,7个氮添加处理的SON转化速率均高于或显著高于对照。
土壤MBN转化速率见图7。培养300 d和培养390 d时,N200和N300处理土壤MBN转化速率为负值;其他处理在培养期内均为正值。N0和N15处理在培养390 d时土壤MBN转化速率最高,N30、N50和 N150处理在培养300 d时最高,N100和N300处理在培养360 d时最高。培养300 d和330 d时,N200和N300处理土壤MBN转化速率显著低于其他处理。培养390 d时,N150、N200和N300处理的土壤MBN转化速率显著低于其他氮处理。
2.3 土壤碳氮转化的耦合关系
在整个原位矿化培养期间,SOC与 Total N,MBC与MBN,土壤DOC与土壤DON,SOC转化速率与净氨化速率、净硝化速率和MBN转化速率的相关性存在明显不同。这说明不同形式的碳发生变化,与之相对应的不同形式氮的反应不同。设定土壤Total N、MBN、DON、净氨化速率、净硝化速率和MBN 转化速率 6 个自变量分别为x1,x2,x3,x4,x5,x6,对应的应变量 SOC、MBC、DOC、SOC 转化速率,分别设定为y1,y2,y3,y4。土壤碳氮转化耦合结果如表4所示,SOC与Total N比值呈极显著正相关,土壤MBC与土壤MBN呈极显著负相关,土壤DOC与土壤 DON呈极显著正相关,SOC转化速率与MBN转化速率呈极显著正相关。

表3 土壤SON、DON和MBN含量变化Table 3 Changes of SON,DON and MBN contents in soil during in-situ mineralization

图6 培养期间SON转化速率变化Fig.6 Dynamics of SON conversion rate

图7 培养期间土壤MBN转化速率变化Fig.7 Dynamics of MBN conversion rate
将8个氮添加处理所有时间段的土壤相关化学指标进行相关分析,结果见表5。土壤MBC与土壤Total N、DOC、和SON呈极显著负相关(P<0.01),与呈显著负相关(P<0.05),与土壤C/N比值呈极显著正相关(P<0.01)。土壤MBN与土壤 Total N、SOC、DOC、和SON呈极显著正相关(P<0.01),与呈显著正相关(P<0.05),与土壤C/N比值呈显著负相关(P<0.05),与MBC呈极显著负相关(P<0.01)。土壤MBC/MBN与土壤C/N和MBC呈极显著正相关(P<0.01),与Total N、SOC、DOC、SON和MBN呈极显著负相关(P<0.01)。土壤DON与土壤Total N、SOC、DOC、、SON和MBN呈极显著正相关(P<0.01),与土壤 C/N比值呈显著负相关(P<0.05),与MBC呈极显著负相关(P<0.01)。土壤MBN转化速率与土壤Total N和DOC呈显著负相关(P<0.05),与土壤SOC和MBN呈极显著负相关(P<0.01),与土壤MBC呈极显著正相关(P<0.01)。土壤MBC转化速率与含量呈显著负相关(P<0.05)。土壤净氨化速率与土壤呈极显著负相关(P<0.01)。土壤净硝化速率与呈极显著正相关(P<0.01)。土壤净矿化速率与土壤呈极显著负相关(P<0.01)。土壤有机碳转化速率与土壤DOC、和MBN呈极显著负相关(P<0.01),与土壤SOC呈显著负相关(P<0.05),与MBC呈极显著正相关(P<0.01)。

表4 土壤碳氮转化的耦合关系Table 4 Coupling relationship between soil carbon and nitrogen conversion
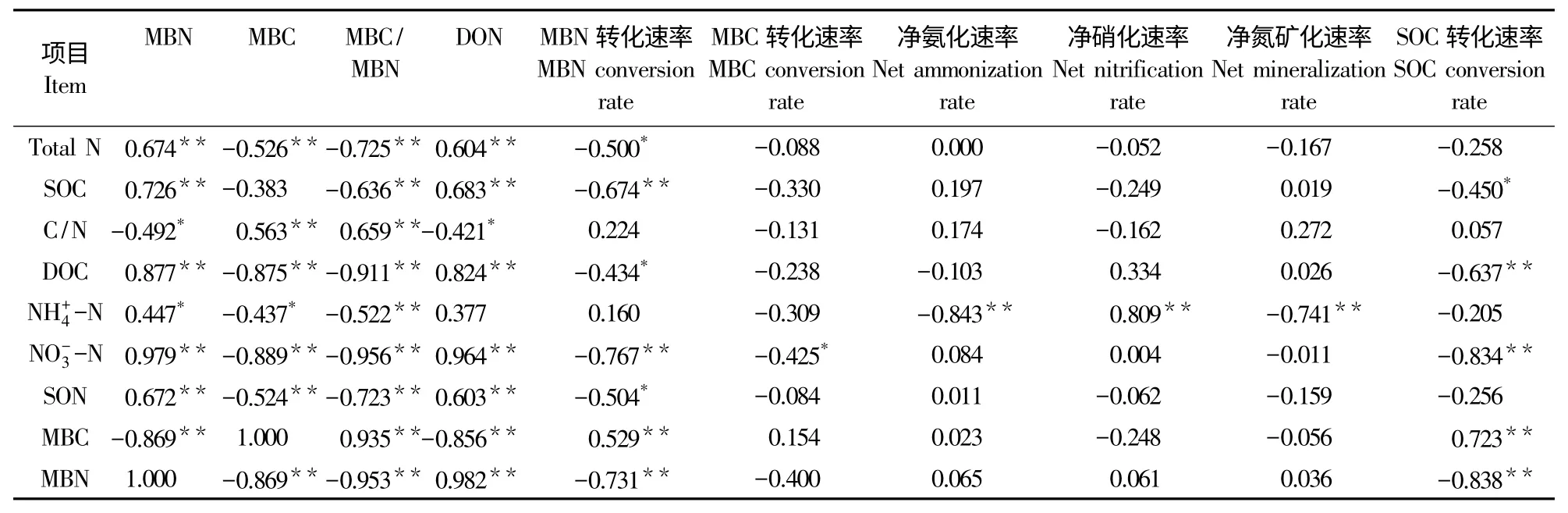
表5 土壤碳氮转化速率与土壤化学指标的相关关系Table 5 Correlation between soil carbon and nitrogen conversion rate and soil chemical index
3 讨论
3.1 氮添加水平对土壤碳氮转化特征的影响
氮沉降对草原生态系统土壤生态过程的影响已成为近年来生态学研究的热点[14,18-19]。适量氮沉降可增加土壤氮供应,对生态系统影响表现为正效应,但过量氮沉降会导致土壤中的碳氮磷化学计量特征改变[20],对土壤养分循环影响表现为负效应。SON的矿化是决定土壤供氮能力的重要生态过程,氮素添加在调控土壤氮转化方面有着重要的作用。本研究表明,氮添加显著增加了土壤的累积氮矿化量,并对土壤硝化、氨化及矿化速率具有显著的影响。在整个培养期,氮添加处理土壤NO-3-N含量显著高于对照,土壤净硝化量最大值出现在7月份(培养330 d)或8月份(培养360 d),而这两个时期正值贝加尔针茅草原雨水充沛的月份,因此可能会导致淋溶损失。进入9月份(培养390 d),所有处理的净硝化速率达到最低值。相比无氮添加对照,氮添加处理在培养大部分时间段增加了土壤净硝化速率,促进了土壤的硝化作用。培养300、330、360 d和 390 d 时,各氮添加处理的土壤含量均高于各处理的含量,且土壤净硝化速率明显高于净氨化速率,表明贝加尔针茅草原土壤氮矿化主要以硝化作用为主,是形成植被生物量的主要的有效氮素。这与邹亚丽等[19]对黄土高原典型草原和王学霞等[18]对青藏高原退化高寒草地的研究结果类似。本试验所添加氮素为硝酸铵,一方面输入,增加了土壤中;另一方面+输入,增加了硝化作用的底物,导致硝化作用加强。
本研究中,氮添加总体上抑制了土壤净氨化速率。土壤净氨化量在整个原位矿化过程中均为负值。负值表示原位培养后的含量低于培养前的初始值,发生了土壤的固持或转化为由于土壤含量变化同时受到硝化作用和氨化作用的共同影响,因此土壤净氨化速率不能全面反映土壤氨氧化作用的强弱。土壤氮矿化作用产物包括二者在土壤中可以相互转化与含量之和的变化速率用来表征土壤的净矿化速率[21]。在整个矿化期除N15处理矿化速率平均值为正值外,其他处理矿化速率平均值均为负值。草地土壤的净矿化速率出现负值,表明土壤无机氮向有机氮转化,消耗无机氮。在整个培养期间,各氮添加处理SON转化速率均为正值(图6),表明SON的固定大于矿化,含量表现为显著降低(表2)。这与培养期内,土壤净硝化作用和净矿化作用与生物固持的相对强弱有关[22]。在培养期内,N15和N30处理促进了氮矿化作用,N50、N100、N150和 N200处理抑制了氮矿化作用。这与张璐等[7]对内蒙古羊草草原的研究结果类似。净矿化速率并不与氮输入量增加呈显著正相关性,当氮添加量达到一定水平后,氮矿化速率会下降[23-24]。Turner等[25]研究表明,在氮饱和系统中,氮输入对氮矿化速率有抑制作用。可能原因是氮添加量增大降低了土壤胞外酶活性;也可能是高氮添加使土壤中可利用氮含量增加,从而降低净氮矿化速率;也可能是高氮添加,引起土壤酸化,降低了土壤微生物活性。项目组前期研究表明,高氮添加降低土壤pH、微生物活性和微生物功能多样性指数[26],降低0~10 cm土层土壤脲酶和过氧化氢酶活性[27]。土壤微生物数量和微生物活性降低会影响土壤净氮矿化速率。培养390 d时,高氮添加(N150、N200和N300)土壤净硝化、净氨化和净矿化速率均为负值,表明这个时期微生物对氮的固定大于矿化。
在整个原位矿化培养期,低氮添加(N15、N30和N50)与对照处理在培养前后SOC含量无显著性变化,而高氮添加(N100、N150、N200和 N300)在培养末期的SOC含量显著低于培养初期。培养300 d、360 d、390 d 时,N50、N100、N150、N200 和 N300处理SOC转化速率为负值,说明在这个时期SOC的矿化大于SOC的积累。总体上,N15、N30处理提高了 SOC 转化速率,N50、N100、N150、N200和 N300处理降低了SOC转化速率。Jussy等[28]研究表明,氮添加抑制酸性森林土壤碳矿化速率。李凯等[29]研究表明,中、高氮处理抑制石栎和苦槠幼苗的土壤呼吸速率。李新爱等[30]研究表明,长期单施化肥不利于稻田土壤有机质和Total N的积累。在整个培养期中,MBC转化速率都为正值,与SOC转化速率不一致。
3.2 土壤原位矿化过程中碳氮转化的耦合关系
碳和氮的耦合特征是草原生态系统过程研究关注的焦点之一[22]。氮沉降改变土壤草原土壤氮转化速率,必将影响土壤碳转化过程。在整个矿化培养期内,SOC含量与土壤Total N存在极显著正相关关系,相关系数为0.848**,且符合一元一次线性回归方程,这说明SOC与土壤Total N含量变化具有一致性。土壤DOC、DON含量与碳氮循环和植物生长密切联系。本研究中,土壤DOC与土壤DON呈显著正相关,相关系数为0.824**。SOC转化速率与MBN转化速率呈极显著正相关,相关系数为0.769**,这说明土壤碳氮转化过程相互影响。土壤微生物是驱动SOC转化的主导因子[31],本研究表明,土壤MBC与SOC转化速率呈极显著正相关。表明土壤MBC是影响SOC转化的重要驱动因子。土壤MBN与SOC转化速率呈极显著相关性,表明土壤MBN对草地SOC的转化有显著影响。土壤MBC、MBN与净氨化速率、净硝化速率和净矿化速率之间无显著的相关性。以往的研究也有类似的研究结果[32-33]。土壤MBC、MBN与净矿化速率无显著相关性,表明土壤MBC、MBN含量对草地土壤的氮矿化的影响有限[8]。矿化培养末期与培养前相比,各处理SON含量表现为增加,低氮添加SOC含量无显著性变化,而高氮添加SOC含量显著降低。在整个矿化培养期,土壤MBN与DOC呈极显著正相关,表明土壤DOC含量受土壤MBN含量的显著影响。
氮沉降对草原生态系统的影响是一个长期的过程,土壤质量的改变是长期积累的效应,是一个缓慢的过程。氮添加水平显著影响贝加尔针茅草原土壤碳氮转化特征、土壤微生物生物量[26]、土壤酶活性[27]和土壤微生物组成。在进行矿化培养过程中,本研究未同时进行土壤微生物活性的测定,而土壤微生物在土壤碳氮矿化过程中起着非常重要的作用。在今后研究土壤碳氮矿化研究中,需进一步结合土壤微生物活性测定。另外,本研究进行原位观测时间较短,只是初步反映了氮添加对土壤碳氮转化的影响。进一步研究土壤碳氮转化过程对氮添加的响应需要更密集的时间点和较长期的矿化培养。
4 结 论
1)氮添加促进了土壤净硝化作用,抑制了净氨化作用,N15、N30处理促进了土壤净氮矿化作用,N50、N100、N150和N200处理抑制了土壤净氮矿化作用。贝加尔针茅草原土壤氮矿化主要以硝化作用为主。N15、N30处理提高了SOC转化速率,N50、N100、N150、N200和 N300处理降低了 SOC转化速率。
2)土壤MBC与MBN呈极显著负相关;SOC转化速率与MBC呈极显著正相关,与MBN呈极显著负相关。SOC转化速率与MBN转化速率呈极显著正相关,土壤碳氮转化过程相互影响。
3)本研究在一定程度上表明,贝加尔针茅草原土壤碳氮转化速率受氮沉降水平的显著影响,连续高氮沉降对土壤碳氮转化过程产生负面影响,不利于土壤碳氮的积累。
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
建材发展导向(2022年6期)2022-04-18
天津医科大学学报(2021年1期)2021-12-05
昆明医科大学学报(2021年10期)2021-12-02
环境与发展(2019年12期)2019-02-12
读写算(2018年6期)2018-08-22
智富时代(2018年10期)2018-01-30
智富时代(2018年10期)2018-01-30
考试周刊(2016年63期)2016-08-15
太空探索(2016年6期)2016-07-10
