
帽儿山典型森林群落优势种空间分布格局及其关联性
2020-06-08 13:45王安彬
山东林业科技 2020年2期
王安彬
(北京林业大学林学院,北京100083)
空间分布格局是指种群个体在水平空间的配置状况或分布状态,它反映了种群个体在水平空间上彼此间的相互关系,是种群生物学特性、种内与种间关系以及环境条件综合作用的结果[1]。种群空间格局的聚集或分散的程度决定了一个物种如何利用资源,是种群空间属性的重要方面,也是种群的基本数量特征之一[2]。对于种群空间分布间格局及其与群落内其它主要种群的空间关联性的研究可以帮助我们认识群落内生物多样性的格局,有助于深化对群落结构的认识,对认识森林群落中树木种群的生物学特性及其相关的生态学过程有极其重要的意义,同时对掌握群落过程演化及预测其变化趋势亦具有重要意义,因此该领域的研究一直是生态学研究的重要方向之一[3-4]。
帽儿山地区森林茂密,生长着大面积的天然次生林和人工林,之前对该地区种群空间格局的研究较少,主要集中天然次生林中水曲柳种群[5]。本研究选取该地区3种典型森林群落即针阔混交林、红松人工林和水曲柳天然次生林,采用目前较为流行的3种空间分布格局分析方法即方差均值比、基于Ripley’sK函数的L函数和角尺度方法进行比较研究群落内乔灌层优势种的空间分布格局与空间关系,以期加深对帽儿山地区典型森林群落空间结构特征和一些主要树种演替规律的认识,为该区域天然次生林的恢复、保护和经营以及人工林的合理经营提供理论依据。
1 研究区概况
研究地点位于黑龙江省尚志市帽儿山地区,位于东经127°18′0″—127°41′6″,北纬45°2′20″—45°18′16″,平均海拔高度300 m,最高海拔为805 m,平均坡度在10~15°,属长白山系张广才岭西部小岭余脉,植被属长白植物区系,是东北东部山区较典型的天然次生林区,植被类型属温带针阔叶混交林,地带性顶极群落为阔叶红松林。本地区土壤多为典型暗棕色森林土,有白浆化暗棕壤、草甸暗棕壤两个亚类,土壤下层为岩石。主要树种有红松(Pinus koraiensis Sieb.et Zucc.)、白 桦(Betula platyphylla)、水曲柳(Fraxinus mandshurica Rupr.)、春榆(Ulmus japonica(Rehd.)Sarg.)、色木槭(Acer mono Maxim.)、胡桃楸(Juglans mandshurica Maxim.)等。
因离海洋较远,且有高山阻隔,大陆性气候较为明显,属温带季风气候。年均温度为2.8℃,1月平均温度-19.6℃,7月平均气温20.9℃,≥10℃积温2883.7℃,年平均湿度为70%,年均降水量为723.8 mm,降雨主要集中在7、8月份,年蒸发量为1093.9 mm,年日照时数2471.3 h,无霜120~140 d。
2 研究方法
2.1 野外调查
2019年7月,在对帽儿山地区森林资源全面调查的基础上,选择有代表性的阔叶红松林、红松人工林和水曲柳天然次生林,每个林分设置20 m×20 m 标准地,对样地中所有的乔木和灌木进行每木调查,其中树高大于1.5 m 记为乔木,小于等于1.5 m记为灌木。调查乔木的树种、胸径、树高、活枝高、冠幅和坐标等;同时调查样方内灌木的树种、高度和盖度等,并记录样地信息,包括坡度、坡向、海拔、郁闭度和起源等。样地基本信息见表1。
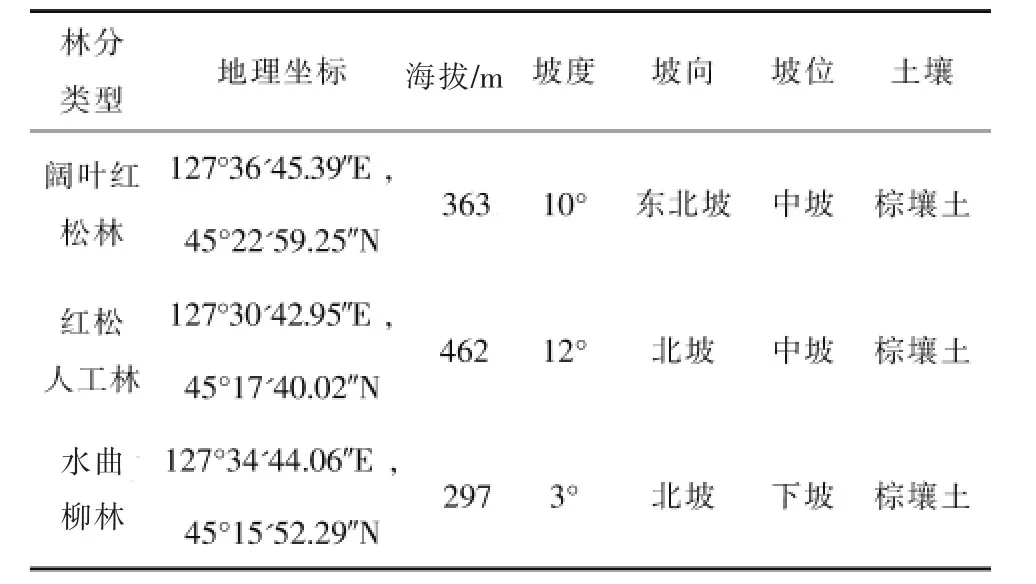
表1 样地基本概况Table1 Basic survey of sample plot
2.2 数据处理分析方法
2.2.1 重要值计算与优势种的确定

式(1)中,乔木显著度用胸高断面积计算,灌木和草本显著度用盖度计算
优势种是指对群落结构和群落环境的形成有明显控制作用的植物种,群落不同层次可以有各自的优势种。本研究中各级优势树种按重要值来确定,乔木层和灌木层重要值大于10%确定为该群落乔灌层的优势种[6]。
2.2.2 种群空间分布格局分析
本文采用方差均值比、Ripley's K函数的变形函数L函数和角尺度3种方法对比分析群落优势种的种群空间分布格局。
方差均值比是常用的空间格局指数之一,其计算公式如下:
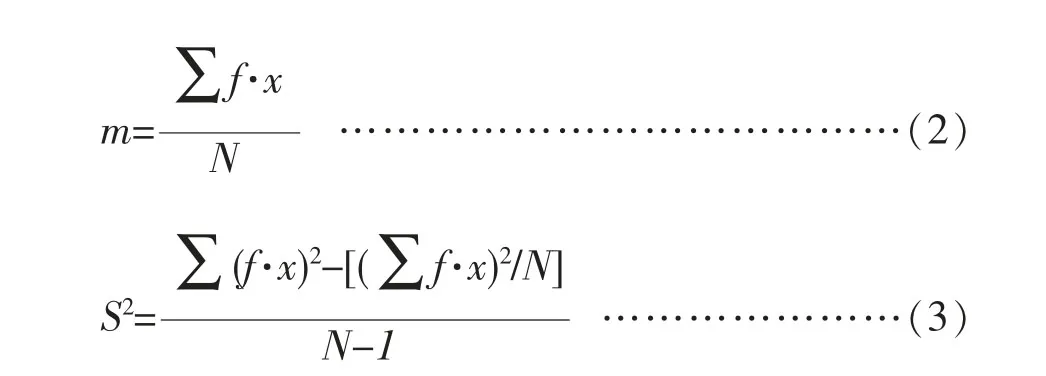
上式中,s:标准差;m:平均数;x:样方中个体数;f:出现频率;N:样本总数;s2/m 即为方差均值比,之后进行t 检验,公式如下:
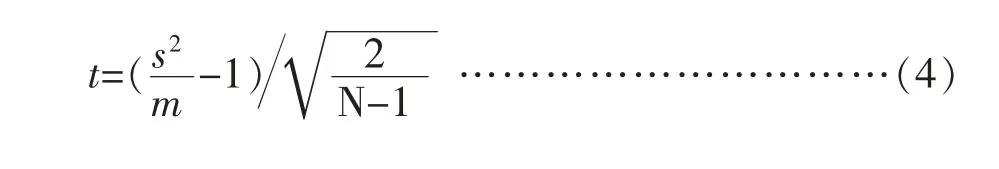

上式中A为样方面积;n为植物总数;Wij为边缘效应校正的权重值;It(i,j)为指标函数,r(ij)为点i,j间距离。
当L(r)落在期望值(即对角虚线)上时,则林木分布是随机分布;当L(r)落在期望值(即对角虚线)的上方时,则林木分布是聚集分布;当L(r)落在期望值(即对角虚线)的下方时,则林木分布是均匀分布。
为了确定计算结果偏离随机状态的显著性,通过1000次Monte Carlo 随机模拟,计算99.9% 置信区间,检验偏离随机格局的程度。以上所有计算在R软件上用spatstat 包完成。
角尺度Wi 是惠刚盈等1998年提出的[9]。从参照树出发,任意两个最近相邻木的夹角有两个,令小角为α,角尺度被定义为α 角小于标准角α0的个数占所考察的最近4株相邻木的比例。Wi=0表示4株最近相邻木在参照树周围分布是特别均匀的状态,Wi=0.5表示4株最近相邻木在参照树周围分布是特别随机的状态,Wi=1 则表示4株最近相邻木在参照树周围分布是特别不均匀的或聚集的状态。角尺度既可用分布的均值表达,更为精细的分析可以角尺度均值W的置信区间为准:随机分布时W 取值范围为[0.475,0.517];W>0.517时为聚集分布;W<0.475时为均匀分布。W 用公式表示为:
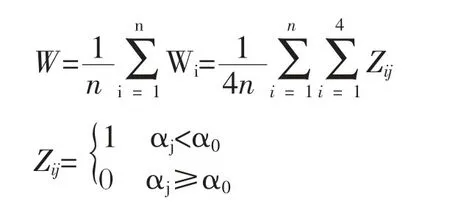
上式中n为林分内参照树的株数;i为任一参照树;j为参照树i的4株最近相邻木;Wi为角尺度即描述相邻木围绕参照树i的均匀性。
2.2.3 种间空间关联性分析
Ripley' s K(r)函数可以推广到类间格局分析,如不同树种之间的空间关联关系分析[10-11]。本研究采用双变量K12函数修正函数L12函数进行双变量点格局分析,从而分析种间空间关系。K12(r)计算公式如下:
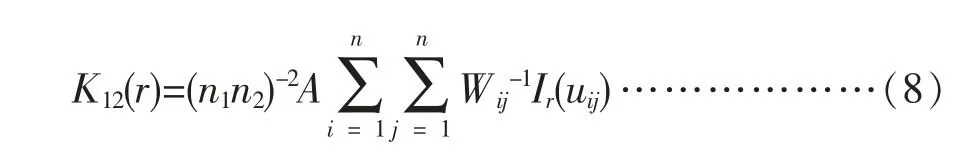
式(8)中n1、n2为格局1和格局2中的点数;当格局1中点i 与格局2中点j之间间距uij 小于r时,Ir (uij)=1;当格局1中点i 与格局2中点j之间间距uij 大于r时,Ir(uij)=0;A为研究区面积、wij为校正因子。
L12函数公式如下:

如果L12(r)=0,表示两类之间是相互独立的;如果L12(r)>0,表示两类之间是聚集的,即正关联;L12(r)<0,表示两类之间是分散的,即负关联。以上所有计算在R3.5.0软件上用spatstat 包完成。
3 结果与分析
3.1 帽儿山典型森林群落基本特征和优势种
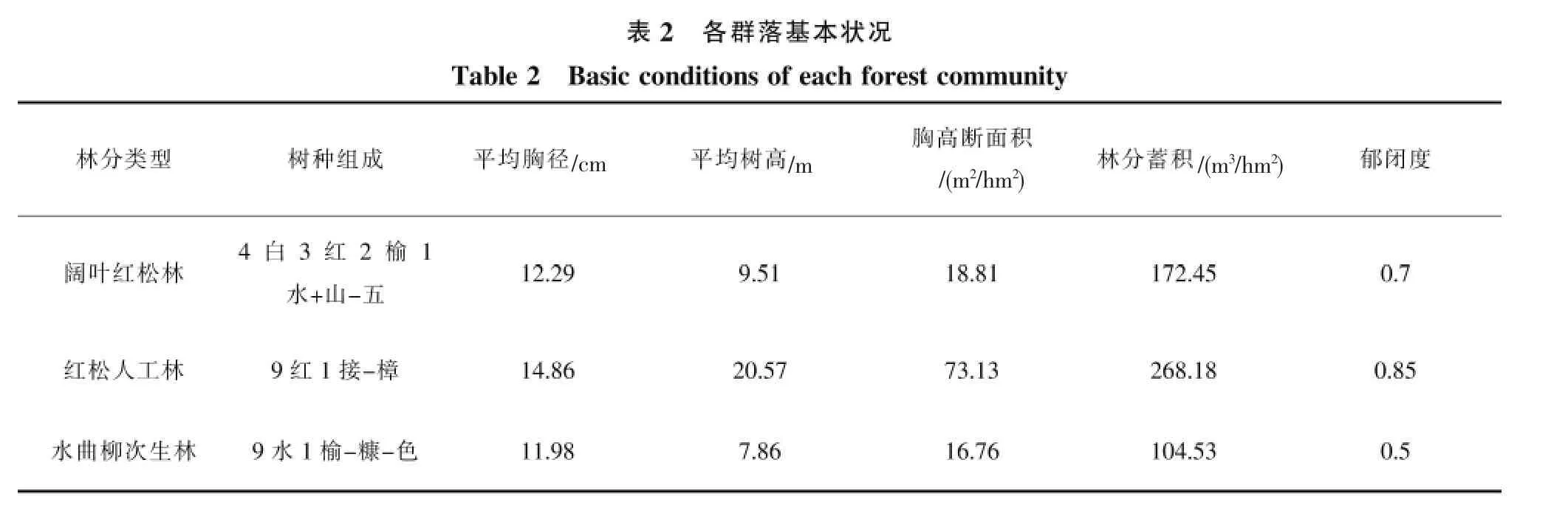
样地资料的统计表明(表2),3个群落树种组成与结构特征有明显差异,分别代表3种不同的群落类型。阔叶红松林针叶树种主要为红松,它和白桦、春榆、水曲柳等阔叶树种组成地带性顶级群落。根据重要值排序,该群落的乔木优势种有红松、白桦、春榆和水曲柳,灌木优势种主要有鼠李、稠李、暴马丁香、卫矛(表3)。红松人工林和水曲柳次生林树种组成比较单一,其中红松人工林的乔木优势种为红松和接骨木,灌木优势种为珍珠梅、忍冬和鼠李;水曲柳次生林的乔木优势种为水曲柳和春榆,灌木优势种为珍珠梅、春榆和鼠李。阔叶红松林和红松人工林的蓄积量、郁闭度均较高,而水曲柳天然次生林由于曾受到过择伐,林分内乔木稀疏,有林隙,郁闭度较低,林分蓄积也较小。
?
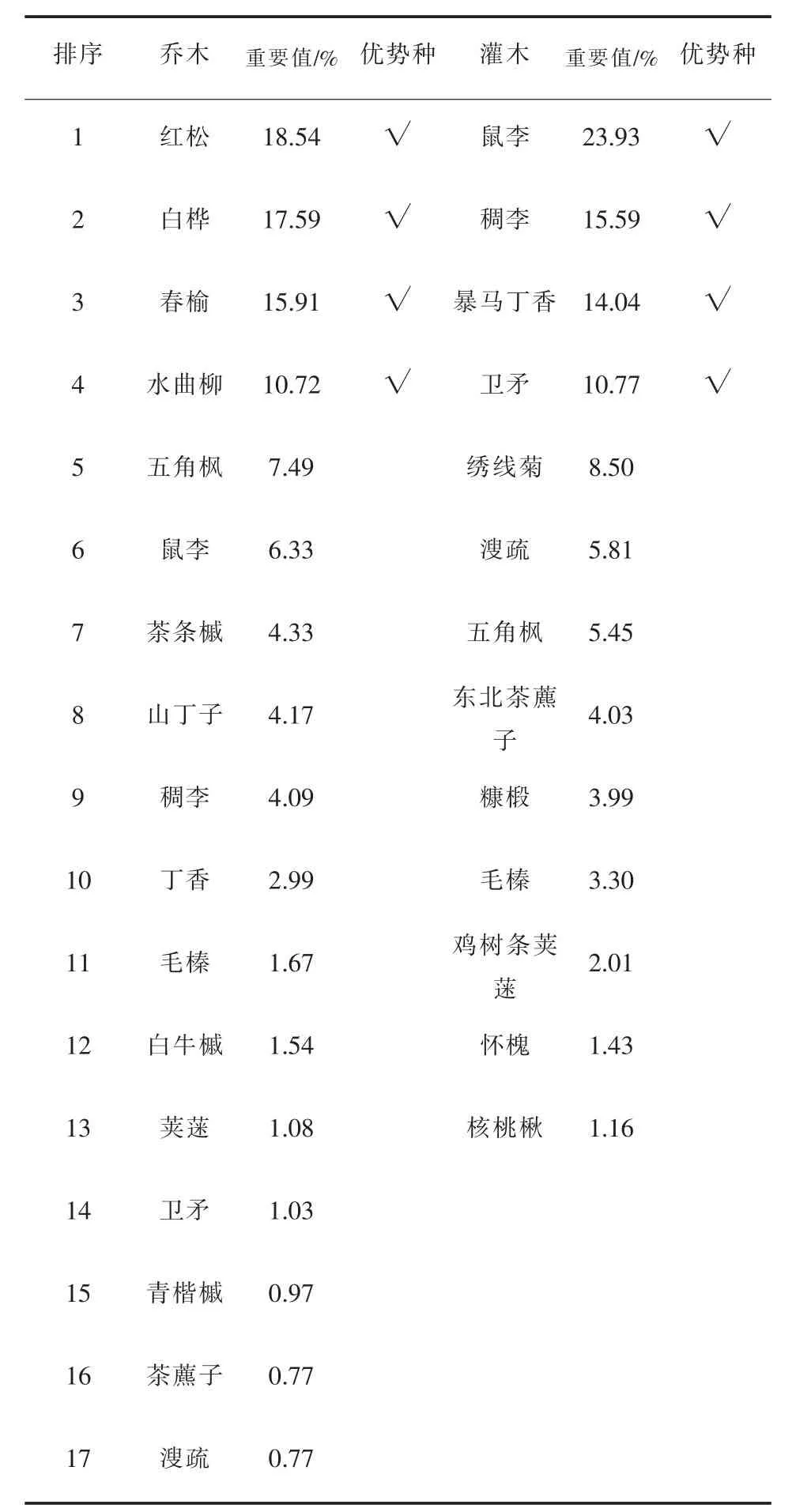
表3 阔叶红松林物种重要值Table3 Important values of species in broad-leaved red pine forests

表4 红松人工林物种重要值Table4 Important values of species in pine plantation

表5 水曲柳天然次生林物种重要值Table5 Important values of natural secondary forest species of Northeast China ash
3.2 群落优势种空间分布格局
3.2.1 方差均值比和角尺度分析
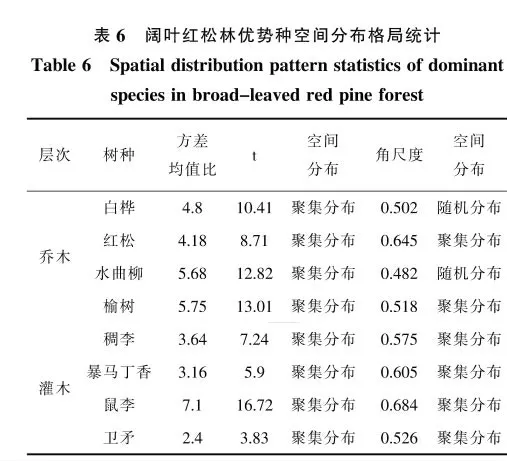
由表6、7、8的统计结果可知,根据方差均值比,阔叶红松林、红松人工林以及水曲柳天然次生林中所有乔木和灌木的优势种均呈聚集分布,而根据角尺度得到的种群空间分布格局则与此有差异,在针阔混交林中,白桦和水曲柳更趋向随机分布,其他优势种呈聚集分布。在红松人工林中,乔木层红松和灌木忍冬呈随机分布,而其他树种也呈聚集分布。在水曲柳天然次生林中两者的结果一致,所有乔木和灌木优势种均呈聚集分布。总体来看,二者大部分结果是一致的。
?
?

?
3.2.2 L函数空间点格局分析
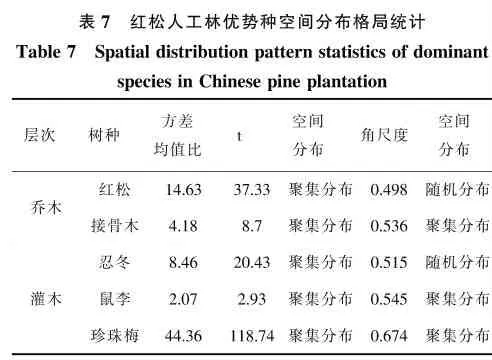
根据图1、图2点格局分析的结果来看,阔叶红松林中乔木优势种的空间分布格局主要为聚集分布或随机分布。其中红松在0-5 m尺度上呈不显著聚集分布;春榆在0-3.5 m 呈不显著聚集分布,3.5-5 m 趋向随机分布;白桦和水曲柳则在0-5 m尺度上均呈随机分布。乔木树种的空间分布格局与方差均值比的结果存在一定差异,但是与角尺度的结果比较相接近,结合树木的空间分布图来看,点格局分析的结果更符合实际状况。主要灌木树种的空间分布格局均为聚集分布,其中稠李聚集程度显著,与方差均值比和角尺度的结果一致。

图1 阔叶红松林乔木优势种空间点格局分析Figure1 Spatial point pattern analysis of dominant species in hardwood redwood forest
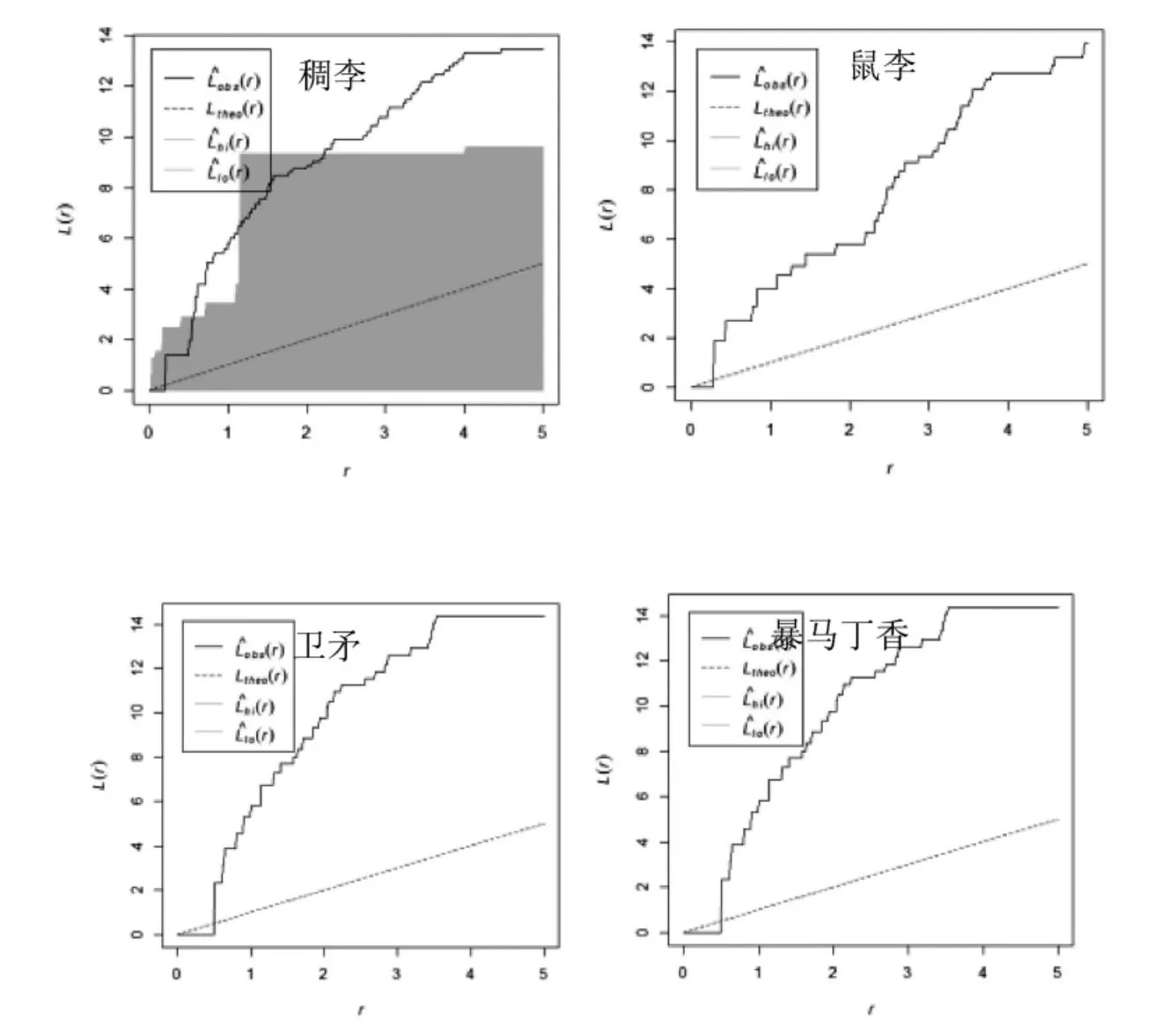
图2 阔叶红松林灌木优势种空间点格局分析Figure2 Spatial point pattern analysis of dominant shrub species in broad-leaved red pine forest
红松人工的乔木优势种红松在0-5 m的尺度上均呈随机分布;接骨木在0-4 m尺度上呈聚集分布,其中在0-1 m 聚集程度显著,在4-5 m 趋向随机分布(图3)。该林分的林下灌木主要为珍珠梅,其空间分布格局在0-5 m尺度呈显著聚集分布。忍冬在0-5 m尺度上呈随机分布,鼠李则在0-5 m 呈不显著聚集分布(图4)。根据图1,可以看出该林分中红松明显更趋向随机分布,而方差均值比显示为聚集分布,与实际情况显然不符。角尺度分析则呈随机分布,与L函数空间点格局分析结果相一致。
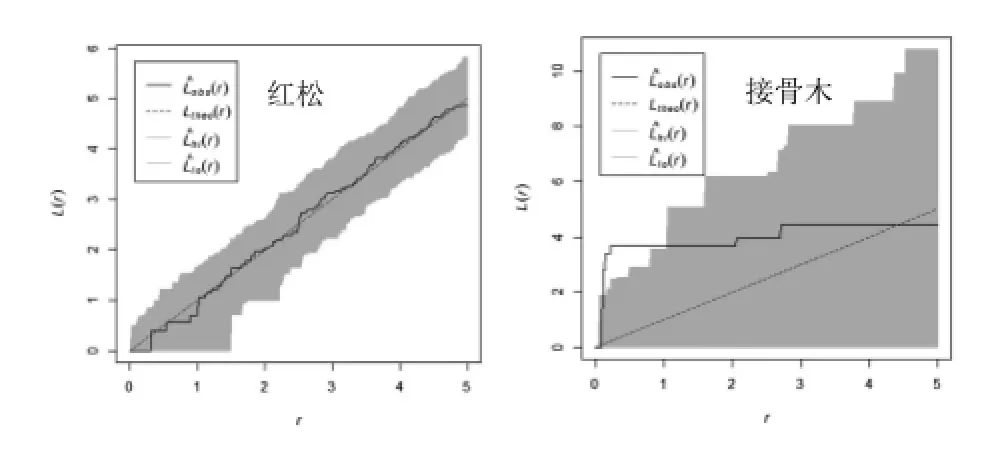
图3 红松人工林乔木优势种空间点格局分析Figure3 Spatial point pattern analysis of dominant tree species in pine plantation
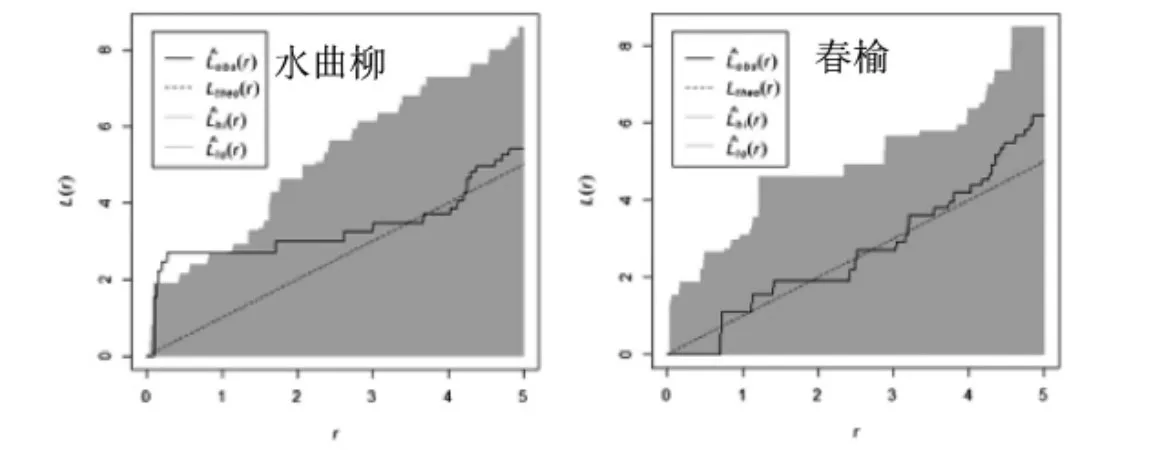
图5 水曲柳天然次生林乔木优势种空间点格局分析Figure5 Spatial point pattern analysis of dominant tree species in natural secondary forest of ash
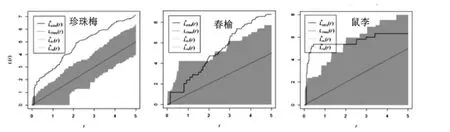
图6 水曲柳天然次生林灌木优势种空间点格局分析Figure6 Spatial point pattern analysis of dominant shrub species in natural secondary forest of ash
根据图5和图6,水曲柳天然次生林中乔木优势种呈随机分布或者不显著聚集分布,水曲柳在0-3 m尺度上呈不显著聚集分布,在3-5 m尺度上呈随机分布;春榆在0-4 m尺度上呈随机分布,在4-5 m尺度上趋向不显著聚集分布。该林分中各种灌木优势种均呈聚集分布,与方差均值比和角尺度的分析结果较一致。其中珍珠梅在0-5 m尺度呈显著聚集分布,春榆在0-3 m 呈不显著聚集分布,在3-5 m 聚集程度显著;卫矛均呈不显著聚集分布;鼠李在0-2 m 聚集分布变为程度显著,在2-5 m 聚集程度不显著。
3.3 群落乔木优势种种间空间关联性
对帽儿山地区三种典型群落的乔木优势种进行双变量点格局分析,结果表明,针阔混交林中针叶树种红松与其他几种阔叶树以及阔叶树种间在空间上呈正关联或空间独立。由图7可以看出,红松与春榆在0-3 m尺度呈正关联,在3-5 m尺度上二者无空间关联。而红松与白桦和水曲柳在0-2 m的尺度上无空间关联,相互独立,但是在2-5 m的尺度上呈正关联。阔叶树种间春榆与水曲柳和白桦均呈正关联,白桦与水曲柳之间则相互独立无空间关联。红松人工林中红松和接骨木以及水曲柳次生林中水曲柳与春榆在0-5 m 均呈正关联,其中在0-1 m 小尺度上呈显著正关联。这说明帽儿山地区这几种林分中在空间上种间竞争不是很激烈,经过长时间的演替进程中的适应,各优势种间以及相互适应,形成良好的空间关系,群落的结构比较稳定。
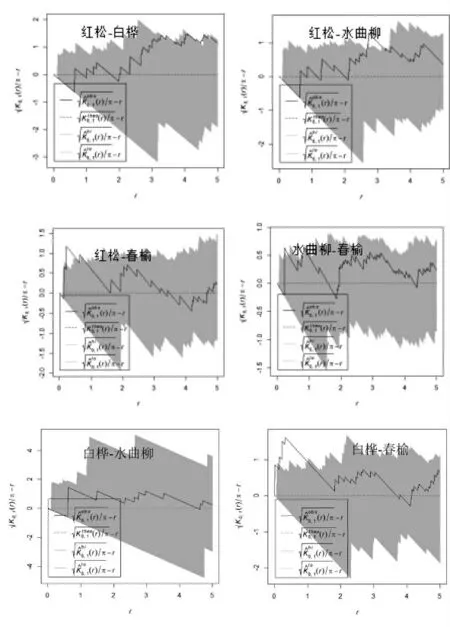
图7 阔叶红松林乔木优势种间空间双变量点格局分析Figure7 Bivariate point pattern analysis of dominant tree species in broad-leaved redwood forests
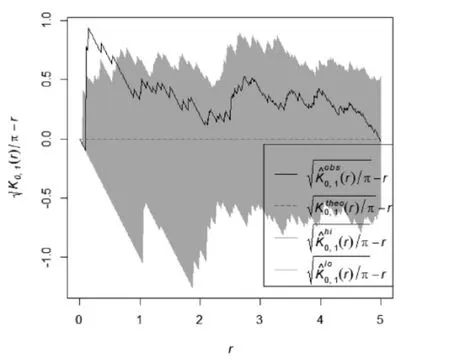
图8 红松人工林红松-接骨木空间双变量点格局分析Figure8 Spatial bivariate point pattern analysis of red pine and elderberry plantation
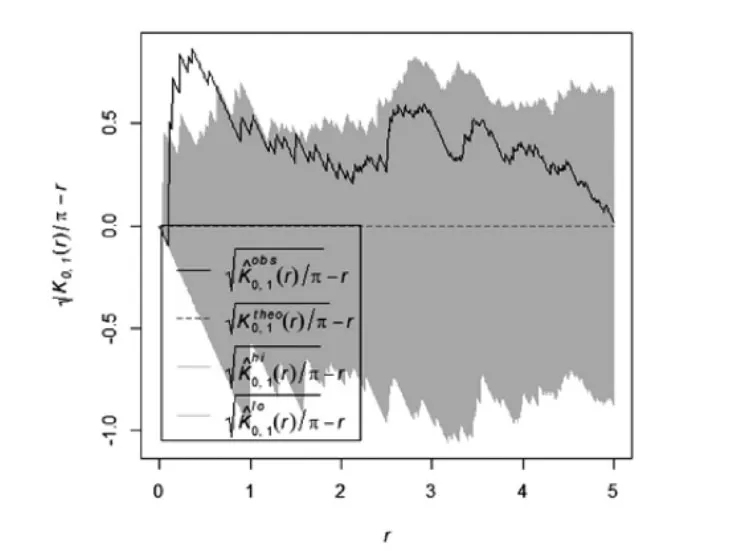
图9 水曲柳天然次生林水曲柳-春榆空间双变量点格局分析Figure9 Spatial bivariate point pattern analysis of mangrove and spring elm natural secondary forest
4 讨论与结论
通过对帽儿山地区阔叶红松林、红松人工林和水曲柳天然次生林的空间格局研究发现,整体来看帽儿山地区植物种群的空间分布格局主要呈聚集分布或随机分布。利用方差均值比计算得到优势种种群空间分布格局均呈聚集分布,角尺度的计算结果则与L函数空间点格局分析结果比较一致,对于阔叶红松林,乔木优势种更多呈随机分布或不显著聚集分布,灌木优势种则呈聚集分布;红松人工林中红松主要呈随机分布,灌木优势种趋向聚集分布。水曲柳天然次生林中,乔木优势种水曲柳和春榆主要呈随机分布,林下灌木优势种均呈聚集分布。从空间关联性上看,3个群落中各乔木优势种间在空间上主要呈正关联或无空间关联。其中阔叶红松林中红松与阔叶树在小尺度上无关联,大尺度上呈正关联,阔叶树种间主要呈正关联。而红松人工林与水曲柳天然次生林乔木优势种间均呈正关联,且在小尺度上关联程度显著。总体来看,红松人工林与水曲柳天然林两个纯林的群落的优势种空间分布格局与空间关联性都比较类似,与阔叶红松林有一定差异。从优势种的空间分布格局与空间关联性来看,三种群落均比较稳定,各物种间空间上的竞争较小,说明群落以及经过演替初期,达到演替的中期甚至晚期。
种群的空间分布格局并不是一成不变的,其受到自身生物学特性、环境条件、初始格局、种间关系等多种因素影响,同一物种的空间分布格局即使在同一地区的不同群落中也会有所差异[12],本研究中在红松人工林与红松阔叶林红松的种群空间格局差异很大,在人工林中,红松呈随机分布,而在针阔混交林中,红松呈聚集分布,主要原因可能是种间关系与年龄阶段的不同。本研究中帽儿山地区大部分优势种呈聚集分布,少量呈随机分布,过去的一些研究也表明大部分植物种群的空间格局为聚集分布,随着森林发育阶段的进行,聚集的程度可能会降低,最后可能演变成随机分布[13],我们的研究结果也与此相一致。而且吴文娟等人对帽儿山地区天然次生林中水曲柳的空间分局于空间关联性研究结果与本研究一致[5],水曲柳与其他树种均呈正相关。优势种群如果具有较高的聚集程度,种群间在空间上呈正相关都有助于维持林分的稳定性,对林分未来的演替趋势产生重要的影响[14],本研究中大部分群落空间结构都比较稳定,但是对于呈随机分布的红松人工林以及水曲柳林,可以采取适当的经营措施,比如在林隙、林下补植一些针阔叶树种,从而提高森林群落的稳定性与生物多样性。
本研究采用了3种典型空间格局分析方法,可以看出方差均值比得到的种群空间格局有些与实际不符,而利用Ripley's K函数的改进函数L函数进行的点格局分析的结果大部分与角尺度结果一致,但L函数充分利用了点与点之间的距离,可以分析任意尺度的林分空间分布格局,其结果更细致。当然Ripley's K函数进行点格局分析受空间尺度的影响很大,一般需要1 hm2以上的大样地来进行空间格局分析[15],本次的样地仅为20m×20m,分析的空间尺度过小,对结果有一定的影响,未来对帽儿山地区典型群落优势种更大尺度上的空间格局还有待深入研究。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
乡村科技(2021年1期)2021-12-03
农村实用技术(2021年6期)2021-12-01
新农民(2020年5期)2020-12-10
农民致富之友(2020年4期)2020-04-03
现代农业研究(2017年11期)2018-01-12
热带农业科学(2017年5期)2017-06-12
农业与技术(2017年9期)2017-05-28
