
利用野生甘蓝改良油菜Ogu CMS恢复材料的菌核病抗性
2020-06-03 09:41万华方丁一娟陈致富梅家琴钱伟
中国农业科学 2020年10期
万华方,丁一娟,陈致富,梅家琴,钱伟
(西南大学农学与生物科技学院/西南大学农业科学研究院,重庆 400716)
0 引言
【研究意义】甘蓝型油菜(Brassica napusL.,AACC,2n=38)是由白菜(Brassica rapaL., AA,2n=20)和甘蓝(Brassica oleraceaL.,CC,2n=18)杂交,并经自然加倍演化而来,是世界上重要的粮油作物和能源作物。中国是世界第二大油菜种植国,其长江流域油菜产业常年受到腐生真菌核盘菌(Sclerotinia sclerotiorum(Lib.) de Bary)的严重危害。由于现有油菜中缺乏菌核病的有效抗源,抗病甘蓝的发现为油菜抗病育种带来了新的思路。细胞质雄性不育(cytoplasmic male sterility,CMS),尤其是萝卜胞质不育系Ogu CMS的不育性十分稳定,是芸薹属作物的安全授粉控制系统[1],也是生产油菜杂交种最好利用的授粉控制系统之一。因此,以含有野生甘蓝优良抗病遗传成分的人工合成甘蓝型油菜为桥梁,培育抗菌核病的油菜Ogu CMS恢复系具有重要理论意义和实践意义。【前人研究进展】由于油菜基因库中匮乏高抗或免疫的资源[2-5],学者们已尝试从油菜的近缘物种中去发掘优良抗源[5-7]。作为油菜的亲本物种之一,甘蓝遗传多样性丰富[8-10],并且由于其长期受到野生环境的自然选择,存在优良的抗病资源。野生甘蓝B.incana具有良好的菌核病抗性,其茎秆抗病性可达到耐病油菜对照中油821的20倍[7],其抗病QTL(quantitative trait locus)分布在C1、C3、C4、C7和C9等多条连锁群[11],与现已报道的油菜抗菌核病QTL位置不同[12-14],其中,部分与甘蓝抗病QTL连锁的标记已成功应用于抗菌核病分子标记辅助选择[15-16]。为将此野生甘蓝的菌核病抗病位点导入油菜,DING等[17]以其为抗源,与白菜进行远缘杂交,发展了人工合成甘蓝型油菜,表现出较好的菌核病抗性。【本研究切入点】油菜中已成功育成了甘蓝型油菜Ogu胞质不育系[18-19]及Ogu CMS的恢复系[20-22],但受限于油菜抗源的匮乏,目前还没有抗菌核病的油菜 Ogu CMS恢复系报道。【拟解决的关键问题】本研究以人工合成甘蓝型油菜桥梁,将野生甘蓝的抗菌核病位点转入油菜,发展具有甘蓝抗病遗传成分且含恢复基因的材料,为发展抗菌核病油菜Ogu CMS恢复系提供种质资源。
1 材料与方法
1.1 试验时间和地点
田间试验于 2013—2016年在重庆市油菜工程技术研究中心试验基地和内蒙古大学试验基地(夏季加代繁殖)进行。
1.2 试验材料及选育策略
前期以高抗菌核病野生甘蓝B. incana(编号C01)与白菜(编号6Y733)杂交,经胚挽救、染色体加倍获得了抗菌核病的人工合成甘蓝型油菜AC1-4(相对于中双9号的枝秆和叶片,相对感病度S分别为0.75和0.57,双高品质)。以AC1-4为父本,对油菜Ogu CMS胞质不育杂种4Q292(源于Ogu CMS油菜与常规油菜的杂种F1,感菌核病,双低品质)人工授粉,通过对后代单株进行抗病位点连锁标记辅助选择(marker-assisted selection,MAS)、可育株抗性鉴定、结实率统计和品质分析,从中选择可育株,自交发展F2代;对F2代单株根据上述方式筛选携带野生甘蓝抗病QTL、且其他性状较优的个体自交;在自交后代中继续重复上述筛选,最后获得携带野生甘蓝抗病QTL、育性正常、产量和品质相对较优的 Ogu CMS恢复系材料。
1.3 分子标记辅助选择(MAS)
以野生甘蓝 C01的带型为参考,利用 4Q292和AC1-4对前期研究中鉴定到的野生甘蓝所有抗病QTL区间及侧翼最近SSR标记进行筛选,获得7个在双亲之间存在多态性的标记,涉及3个叶片抗性位点和2个茎秆抗位点(表 1)。利用表中标记对各世代单株进行分析。尽管其抗病QTL分布在C1、C3、C4、C7和C9等多条连锁群[11],但C1和C9联锁群上的2个QTL(C1-QTL和 C9-QTL)解释的表型变异度相对较高(13.6%—24.1%),为主效QTL,且QTL均具有至少2个连锁的多态性标记,因此,MAS过程中首要选择同时携带C1和C9抗病位点,其次考虑是否携带C3和C7上QTL连锁标记(对表型贡献率相对较低,且均为单标记选择)。MAS于苗期在每个世代开展,经MAS选中的单株进入后期的抗性鉴定、结实性调查和品质分析。
1.4 菌核病抗性鉴定
核盘菌菌种为 1980。参照 MEI等[23]的方法进行菌种活化、接种体(PDA菌丝块)制备及菌核病抗性鉴定。于油菜终花期后2周进行离体枝秆菌核病抗性鉴定:取一次有效分枝,截取长约30 cm一段,将两端用保鲜膜缠绕(以防止水分散失),然后转移到可控温、控湿的室内,将枝秆随机摆放于铺好湿毛巾的接种台上,并将准备好的PDA菌丝块带菌面紧贴于枝秆上,每枝秆接种2—3个菌丝块。接种完,在接种台上放上支架,以塑料膜密封,控制温度((22±1)℃)和湿度(90%—95%)。接种后第 4天测量菌斑的长度,以相对于耐病对照甘蓝型油菜中双9号的感病度(S)评价材料菌核病抗性,即S=L待测/L对照(L待测表示材料接种核盘菌的病斑长度(cm),L对照表示耐病对照中双9号的病斑长度(cm))。
1.5 结实率统计及籽粒品质分析
每个单株分别选取主花序上连续 50个自交角果和离主花序最近且未套袋一次分枝上 50个自由授粉角果,统计每角果籽粒数,分别代表自交和自由授粉结实率。用FOSS NIR system3700型近红外光谱仪检测自交种籽粒品质。光谱扫描条件:分辨率8 cm-1、扫描次数64次、光谱范围1 100—2 500 nm、温度20—25℃。
1.6 统计分析
采用SAS 8.0软件进行数据分析。
2 结果
以高抗菌核病的野生甘蓝C01为父本,与编号为6Y733的白菜杂交,10 d后取子房解剖后进行胚挽救及染色体加倍,获得人工合成甘蓝型油菜AC1-4,其苗期形态介于野生甘蓝(图 1-A)和白菜(图 1-B)之间,与甘蓝型油菜形态相似(图 1-C),分子标记也表明,AC1-4为B. incana和6Y733的真杂种(图2)。对杂种F1进行体细胞染色体观察发现,单倍体植株均含19条染色体(图1-F),经染色体加倍的AC1-4植株体细胞均含有38条染色体(图1-G)。花粉育性鉴定结果表明,单倍体植株的花粉育性(图 1-H)远低于人工合成甘蓝型油菜AC1-4的花粉育性(可达80%)(图1-I)。叶片和枝秆菌核病抗性鉴定结果(图1-J和图1-K)表明,人工合成甘蓝型油菜AC1-4的叶片抗性和枝秆抗性都显著高于对照甘蓝型油菜中双9号和白菜亲本。

图1 高抗菌核病人工合成甘蓝型油菜AC1-4的特征Fig. 1 Characteristics of AC1-4 (resynthesized Brassica napus) with high resistance against Sclerotinia sclerotiorum

图2 SSR引物SWU260检测亲本及F1Fig. 2 Verification of the relatives and F1 hybrid individual plants with SSR primer SWU260
人工合成甘蓝型油菜AC1-4与Ogu CMS甘蓝型油菜杂种4Q292(图1-D)杂交产生的F1代(图1-E)代共获得146个单株,其中69株可育,77株不育。经卡方检验,F1代育性符合1∶1分离,(χ21∶1= 0.17,P>0.05),符合预期分离比。对F1代146个单株进行分子标记辅助选择,其中128个单株遗传了源于抗病甘蓝B. incana的C1-QTL和C9-QTL,另外分布在C3和C7染色体的2个标记也在上述128个单株中表现出AC1-4与4Q292的杂合带型。剩余的18个单株中,有 11个单株丢失了 C1-QTL,2个单株丢失了C9-QTL,5个单株的C3或C7-QTL连锁标记未表现出抗病带。F1代植株的自交结实率与天然结实率显著相关(r= 0.52),但套袋自交的角果粒数变异较大,最多达24粒/角果,最少为5粒/角果,平均为9.07粒/角果。菌核病抗性鉴定表明,含有所有标记抗病带的49个F1可育单株的相对感病度S介于0.2—1.4,平均值为0.74,而不完全遗传上述7个标记抗病带的6株可育株相对感病度S介于0.9—1.5,平均值为1.23。结合结实率、分子标记结果以及收获种子状况(种子量及饱满度),选取编号为4C99和4C115的2个F1单株进入下一轮筛选,2份材料的相对感病度S分别为0.66和0.40,自交结实率分别为12.4和15.0粒/角果,自由授粉结实率分别为17.9和21.4粒/角果。
将4C99和4C115的自交种进行了夏繁种植,但由于环境条件的变化,导致大量材料F2单株未开花,最终4C99自交后代有59株开花,4C115有65株开花。对上述124个F2单株进行C9上枝秆主效抗病QTL连锁标记筛选,发现源于4C99的59个F2单株中有35个植株携带 C9-QTL,源于4C115的F2单株中有34个植株携带C9-QTL。由于夏繁地无抗性鉴定条件,因此,未对F2代植株进行抗性鉴定和筛选。随后对可育株上获得自交种的材料进行了籽粒品质分析,最终结合标记分析及品质分析数据筛选了8个F2,其芥酸含量介于7.50%—24.70%,硫苷含量介于15.88—78.63 μmol·g-1饼粕。利用该8个F2单株进行自交发展下一代,田间种植共获得的8个F2:3家系(以5X进行编号),家系大小介于46—184株。利用分子标记对8个家系进行分析(表2),F2:3代材料基因型分离明显,无论是C1还是C9上的QTL在各个家系中均存在分离,其中4个家系(5X78、5X80、5X82和5X84)中出现同时携带C1-和C9-QTL的概率(36.6%— 43.5%)显著高于其他4个家系(0—8.7%)。8个家系共计获得222株同时携带C1-和C9-QTL的植株。为了进一步缩小选择范围,利用所有抗病标记结果进行了各家系材料与抗病亲本的聚类分析,最终从5X78、5X80、5X81、5X82、5X83和 5X84中分别筛选到 4株、23株、2株、16株、1株和4株(共计50株)与抗性亲本遗传关系较近的可育株,而5X77和5X79 2个家系中没有发现与抗性亲本聚在一起的植株。对此50份材料进行枝秆菌核病抗性鉴定,同时以25株完全或部分缺失了抗病QTL的材料作为对照,结果显示,50份材料的相对感病度S介于0.34—0.89,平均值为0.60,而完全或部分缺失了抗病QTL的植株平均相对感病度S为0.98,显著高于前者(P<0.01)(图3)。对部分材料进行了籽粒品质检测(表3),发现所有材料均表现为双高品质。尽管如此,材料之间的硫苷和芥酸含量表现出分离状态,个别材料如5X82-2和 5X82-7的菜籽品质已较为接近单低水平,其中5X82-2 的硫苷含量为 45.38 μmol·g-1饼粕,5X82-7 的芥酸含量为6.5%。同时,这两个材料的相对感病度S分别为0.33和0.48(表3),表明其具有单低的品质,且菌核病抗性已得到改良。结合 MAS和枝秆抗性鉴定结果,上述材料中相对感病度S不超过0.5的单株自交种将进入下一轮筛选。
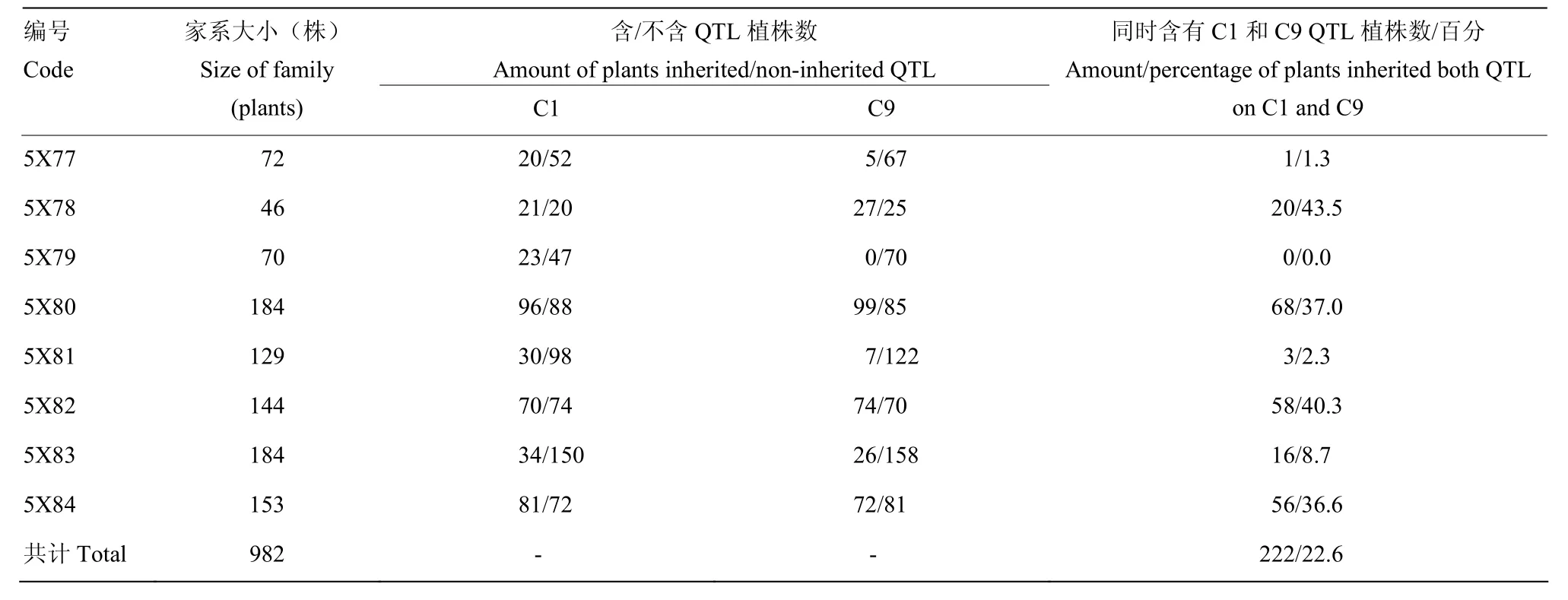
表2 各F2:3家系分子标记分析结果Table 2 Results from molecular marker analysis of F2:3 families

图3 各F2:3家系中选植株的菌核病抗性Fig. 3 The Sclerotinia resistance level of selected individuals in F2:3 families
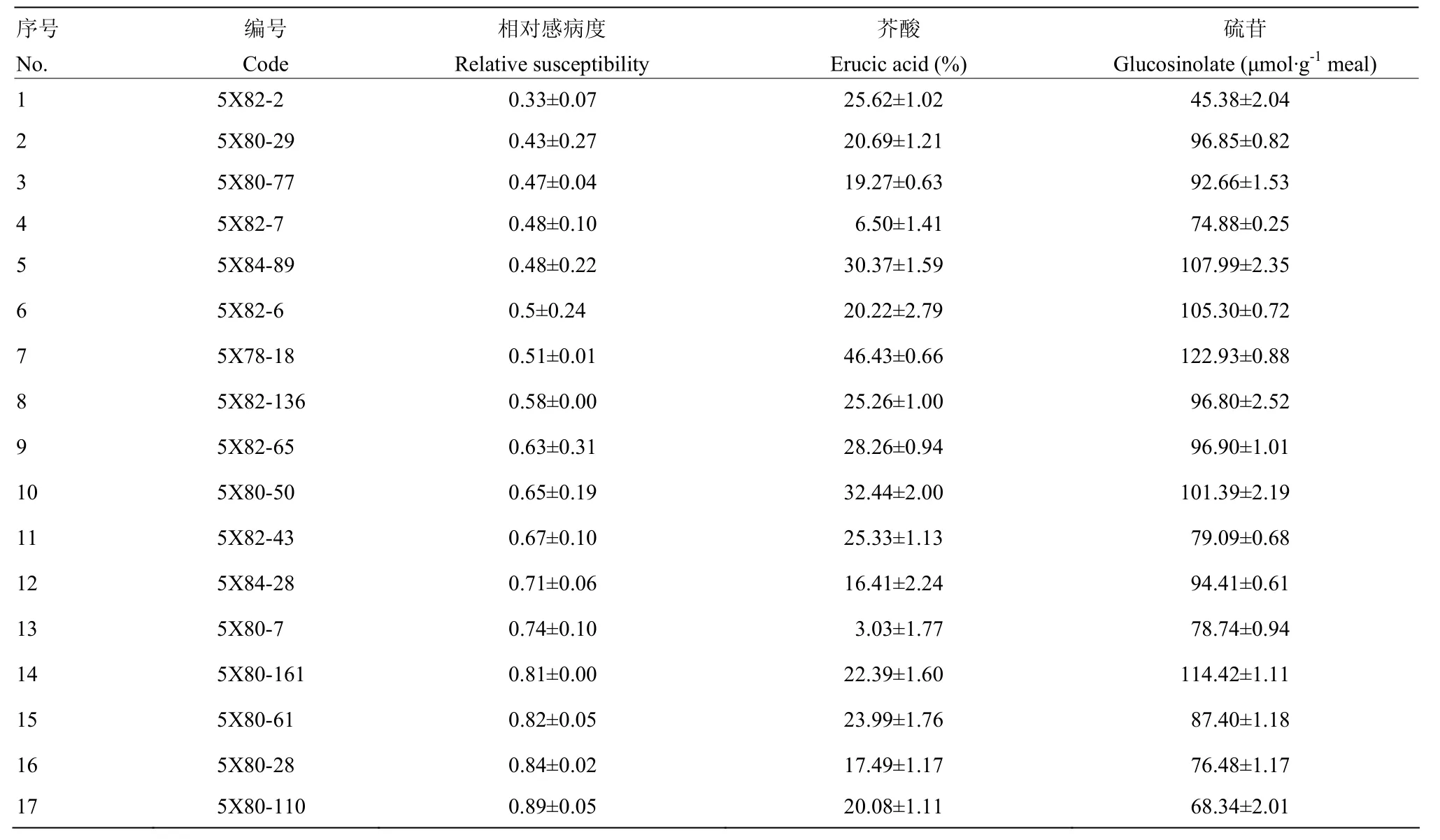
表3 F2:3家系中部分单株抗性和籽粒品质Table 3 Relative susceptibility and seed quality of some individuals in F2:3 generation
3 讨论
甘蓝型油菜的遗传背景因“双低”选育过程而趋于狭窄,但其二倍体祖先种白菜和甘蓝起源早、栽培历史悠久,形态、农艺性状具有丰富的变异[24]。利用甘蓝和白菜培育的人工合成甘蓝型油菜,已被广泛运用到甘蓝型油菜遗传育种,比如胞质雄性不育系“Bro CMS”创制[25]、大粒甘蓝型油菜创制[26],黄籽甘蓝型油菜品系创制[27-28]、早熟性状改良[29]、黄萎病抗性的种质创建[30]、根肿病抗性改良[31]以及Ogu育性恢复基因(Rfo)转育[32]等研究中。人工合成甘蓝型油菜与天然甘蓝型油菜遗传差异大,可有效拓宽天然甘蓝型油菜的遗传资源。然而,人工合成甘蓝型油菜的含油量较低,芥酸、硫苷含量较高,甚至出现一些源于祖先种的不良性状,常需要不断进行品质改良。本研究所使用的 AC1-4也表现出双高品质,以其为亲本发展的低世代中出现芥酸或硫苷含量较高的材料。后续研究中,将进一步发展高世代,进行双低品质材料的选择。
Ogura细胞质雄性不育败育彻底、转育容易,是芸薹属作物的安全授粉控制系统[1]。甘蓝型油菜 Ogu胞质不育系[18-19]及 Ogu CMS的恢复系[20-22]已成功创建,但目前油菜基因库中缺乏有效抗源,限制了抗菌核病的油菜Ogu CMS恢复系的研究。甘蓝型油菜的祖先种甘蓝是改良油菜菌核病抗性的优良资源[11,33]。在早期研究中,鉴定到野生甘蓝B. incana具有较好的菌核病抗性[7],并对其抗病位点进行了QTL定位[11]。利用该材料我们创制了人工合成甘蓝型油菜,该人工合成材料理论上应该遗传B. incana的整套C基因组染色体,因此,该人工合成系与Ogu CMS油菜的杂种F1所有植株理应继承B. incana的抗病QTL位点。然而,利用QTL区间或侧翼标记对F1代的标记分析后发现,有极少数单株存在个别QTL丢失的情况。这可能是由人工合成甘蓝型油菜的遗传不稳定造成的:人工合成甘蓝型油菜含有A、C两套共线性较高的基因组,减数分裂时可能产生A、C染色体之间的异源配对[34-35],形成多价体及单价体[36-37],从而导致染色体分离紊乱和非整倍体配子的产生[38-39],使某些遗传信息在后代可能丢失。
本研究利用人工合成系和Ogu CMS油菜寻找到7个与抗病QTL连锁的多态性标记,涉及4个染色体上5个抗病QTL(包括叶片抗性QTL和茎秆抗病QTL)。因此本研究首先采用C1和C9抗病QTL两侧均有多态性的标记对材料进行分析和选择,而其他3个QTL只开放出一个单标记,这些标记仅用于辅助选择和遗传分析,而用于MAS的准确性低。油菜及甘蓝的菌核病抗性均为数量性状,主要受到加性效应控制[11-14,40],因此,本研究在每个世代尽量筛选同时具有C1和C9 QTL、且尽量在其余3个标记结果中选择具有抗病甘蓝带型的个体。后期的抗性鉴定显示,标记选中的材料平均抗性显著高于 QTL出现丢失的材料,一定程度证明了标记选择的有效性。
结合 MAS和抗性选择,尽管在每个世代均能鉴定到同时携带多个抗病QTL且相对感病度S较低的单株,但是从对 F2:3家系的分析中可以看出,所有家系中的单个抗病位点均未达到纯合,且硫苷和芥酸含量也表现出分离。但是,将根据 1.2的选育策略对 F2:3中选择到的候选植株进行进一步的选择,且后期的选择将注重以家(株)系为单位进行分析,以确定目标位点及控制育性的核基因Rf是否纯合;同时需要增加家(株)系中个体数,从而出现更多的交换重组,从中获得既具有抗病位点、又具有低硫苷和低芥酸位点的重组个体;再者,可以考虑通过目前获得的抗病且芥酸或硫苷含量接近单低的个体相互杂交,聚合不同性状的优良位点,从而获得优质且抗性提高的恢复株系。
4 结论
以人工合成甘蓝型油菜为桥梁,野生甘蓝B.incana的菌核病抗性可被有效转移到甘蓝型油菜,从而改良油菜Ogu CMS恢复材料的菌核病抗性。
猜你喜欢
四川农业科技(2022年2期)2022-11-21
作物学报(2022年12期)2022-10-14
今日农业(2022年4期)2022-06-01
作物学报(2022年4期)2022-02-10
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
四川农业科技(2021年7期)2021-08-20
四川蚕业(2021年3期)2021-02-12
发明与创新·大科技(2019年5期)2019-07-31
现代农业科技(2009年17期)2009-03-08
