
4种园林树木叶片秋季变色期的呈色机理
2020-05-22 09:10:02王萌常格王琦李娟娟路斌路丙社
林业与生态科学 2020年1期
王萌,常格,王琦,李娟娟,路斌,路丙社
(河北农业大学 园林与旅游学院,河北 保定,071000)
植物叶片的颜色是由植物色素在细胞中所处位置以及其在细胞中所占比例、含量所决定的[1]。彩叶植物的叶色表现是遗传因素和外部环境共同作用的结果,通过改变植物叶片中各种色素的种类、含量以及分布形成了多彩的叶色[2]。秋色树种对美化环境、丰富园景以及调节春秋两季之间宏观景色的差距具有良好作用,是园林植物造景中不可缺少的重要观赏植物[3]。如今,三角枫[AcerbuergerianumMiq. var.ningpoense(Hance) Rehd.]、血皮槭[Acergriseum(Franch.) Pax]、丝棉木(EuonymusmaackiiRupr.)和北美豆梨(PyruscalleryanaDcne.)因其秋季叶色变红的观赏特性而逐渐被广泛应用,但是目前国内对于其研究局限于栽培繁殖方面,对于叶色方面的研究仍处于初步阶段。本试验通过对比4种秋季叶色变红的树种在秋冬变色期叶色参数和色素含量的变化情况,研究不同树种叶片叶色变化与叶色参数、色素以及可溶性糖含量的关系,旨在探讨秋色叶树种的呈色机理,为园林应用提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为河北农业大学西校区校园内的4种秋季叶色变红的园林树木,分别为:三角枫、血皮槭、丝棉木和北美豆梨。
试验于2018年10月18日至11月11日进行。选取树冠外围中上部的叶片,每隔6 d采取生长良好的功能叶5片用冰盒立即带回实验室进行指标的测定,共取样5次。
1.2 指标测定方法
1.2.1 叶色参数的测定 采用全自动色差计CR-400测定叶色参数,记录L*、a*、b*值,每个指标3次重复[4]。L*表示光泽明亮度,其值越大则明亮度越高;a*值表示样品红绿饱和度,正为红,负为绿,a*越大,红色越深,反之绿色越深;b*值表示样品黄蓝饱和度,正为黄,负为蓝,b*越大,黄色越深,反之,蓝色越深。
1.2.2 色素含量的测定 采用95%乙醇浸提法[5],测定叶绿素和类胡萝卜素含量,采用盐酸甲醇浸提法[6],测定花色素苷的含量,每个指标3次重复。
1.2.3 可溶性糖含量的测定 采用蒽酮比色法[5]。
1.3 数据处理
使用Excel 2010进行数据整理,SPSS 22.0进行数据统计分析。
2 结果与分析
2.1 叶片变色期叶片叶色参数的变化
叶片变色期叶色参数L*的变化,叶片变色期叶色参数a*的变化及叶片变色期叶色参数b*的变化分析见图1、图2和图3。
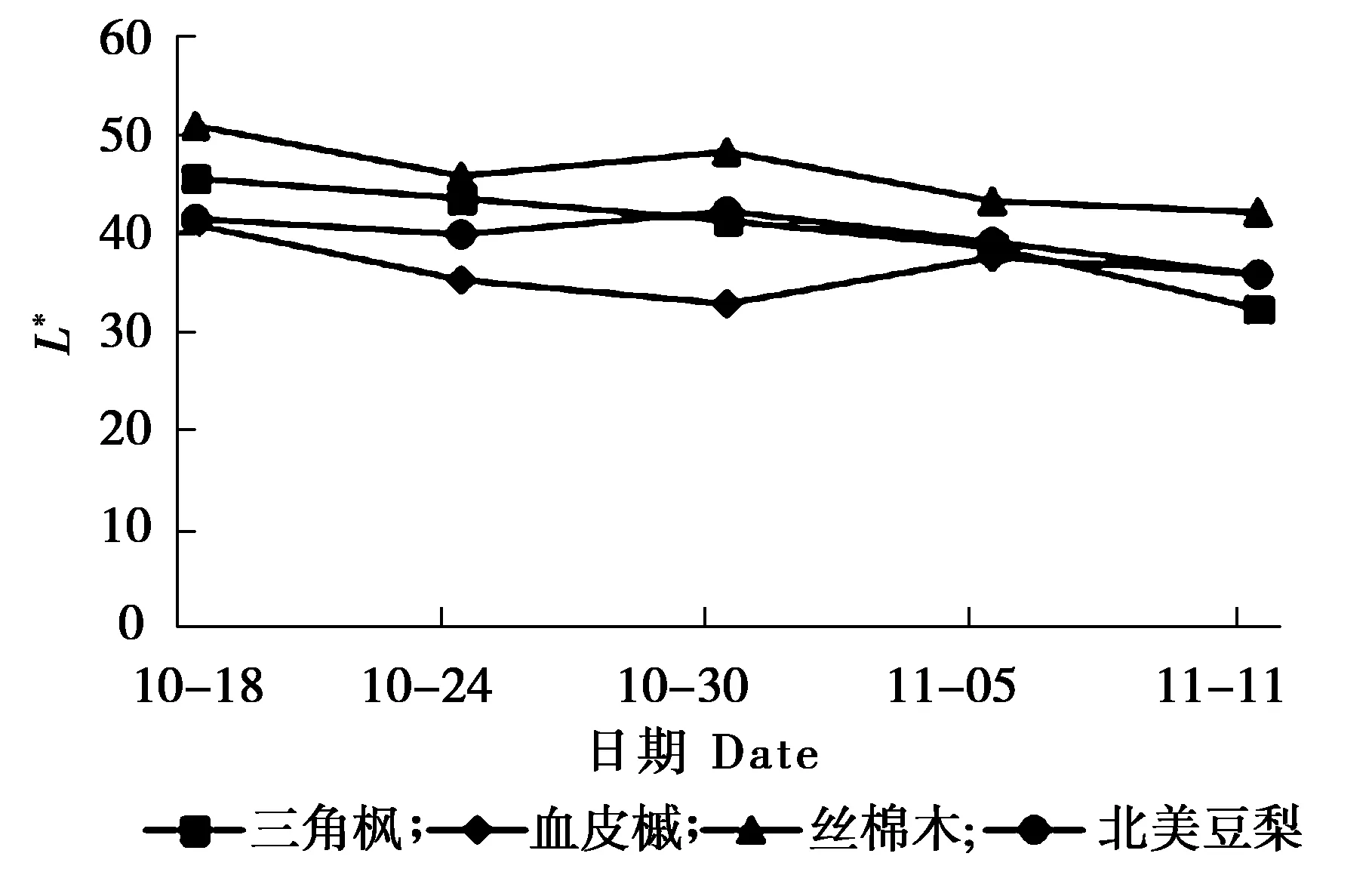
图1 叶片变色期叶色参数L*的变化
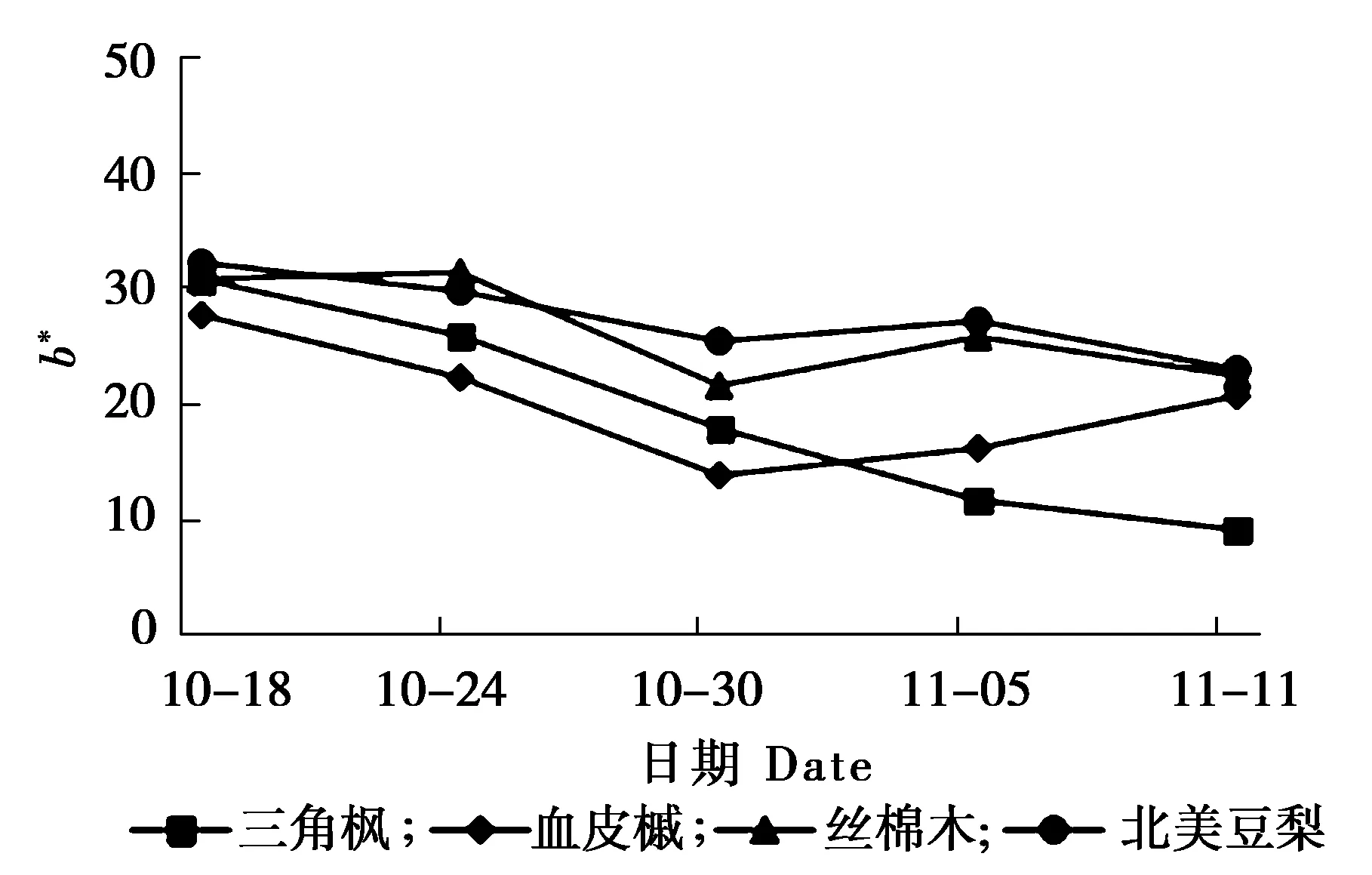
图3 叶片变色期叶色参数b*的变化
由图1可知,L*表示光泽明亮度,其值越大明亮度越高。在秋季整个叶色变化过程中4个树种的L*值均有一定程度的降低,但变化幅度较小,无明显差异。
由图2可知,a*表示红/绿的比值,正值偏红,负值偏绿。随着时间的变化,各树种叶片的a*值均呈不断上升的趋势,叶片也逐渐由绿变红。10月18日a*值均最小,三角枫、血皮槭、丝棉木、北美豆梨的a*值分别为-17.78、-16.92、-16.29、-14.83。10月30日,三角枫、血皮槭以及北美豆梨的a*值已达到正值,丝棉木仍为负值。丝棉木在10月18-30日期间,增长幅度较缓慢,10月30日-11月11日时,丝棉木的a*值较其他3种乔木增长幅度最大。11月11日,4种树木的a*值均达到最大值,血皮槭与丝棉木的值分别为41.30、39.08,三角枫与北美豆梨的最大值分别为24.36、32.46。a*值随时间的延长不断增大,表示叶片逐渐变红。
由图3可知,b*表示黄蓝程度,b*值越大,呈现黄色越深;反之,蓝色越深。三角枫在整个变色过程中b*值一直处于下降的趋势,血皮槭呈现先下降后上升,丝棉木和北美豆梨b*值无明显变化规律。
2.2 叶片变色期叶片色素含量的变化
叶片变色期色素含量的变化见图4。

图4 叶片变色期色素含量的变化
Figure 4 Changes of pigment content during leaf color transition
图4反映了在秋冬叶片转色期,三角枫、血皮槭、丝棉木、北美豆梨叶片中叶绿素a、叶绿素b、叶绿素(a+b)、类萝卜素含量的变化情况。在叶色变化过程中,各树种叶绿素a、叶绿素b、叶绿素总含量的变化趋势相似,整体呈现先快后慢的下降趋势。其中,各树种叶绿素a含量的降幅在10月30日前均较为明显,10月30日后趋于平缓。叶绿素b含量的降幅与叶绿素a相比较为缓慢。类胡萝卜素含量的变化趋势在不同树种间差异较大,而类胡萝卜素含量在整个变色期变化趋势较为平缓。
2.3 变色期叶片中花色素苷和可溶性糖含量变化
叶片变色期花色素苷和可溶性糖含量的变化见图5。
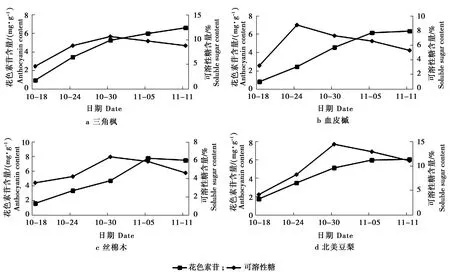
图5 叶片变色期花色素苷和可溶性糖含量的变化
Figure 5 Changes of anthocyanin and soluble sugar contents during leaf color transition
由图5可知,在秋冬叶片转色期,三角枫、血皮槭、丝棉木和北美豆梨各树种叶片中花色素苷和可溶性糖含量的变化。各树种之间叶片中花色素苷含量变化趋势一致,呈现较为明显的上升趋势,随着花色素含量的上升,叶片颜色经历绿色—红绿相间—红色这个变化过程,最终达到秋冬季最佳观赏效果。可溶性糖含量在不同树种间存在差异,呈单峰曲线的趋势,在10月30日均可达到峰值。在10月30日之前,可溶性糖含量增幅比花青素明显偏高,达到峰值后,花色素苷的含量增长幅度变大。
2.4 变色期各树种叶片花色素苷含量与叶绿素含量比值的变化
叶片变色期花色素苷和叶绿素含量比值的变化见图6。由图6可知,在秋冬叶片转色期,三角枫、血皮槭、丝棉木和北美豆梨各树种叶片中花色素苷含量与叶绿素含量比值的变化趋势基本相似。在试验期间,花色素苷与叶绿素含量之比呈不断上升的趋势,前期增长缓慢,在11月5日之后迅速升高,但各树种花色素苷与叶绿素含量之比上升幅度存在明显差异,增幅程度由高到低分别为丝棉木、血皮槭、三角枫、北美豆梨。
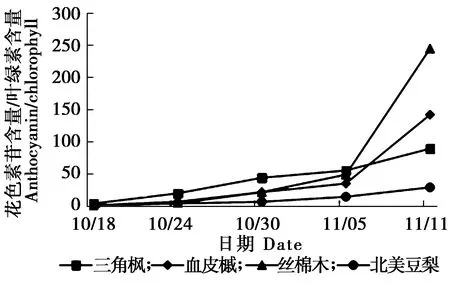
图6 叶片变色期花色素苷和叶绿素含量比值的变化
2.5 叶片变色期各树种叶片呈色相关指标的相关系数
不同树种叶片变色期呈色相关指标的相关系数见表1。
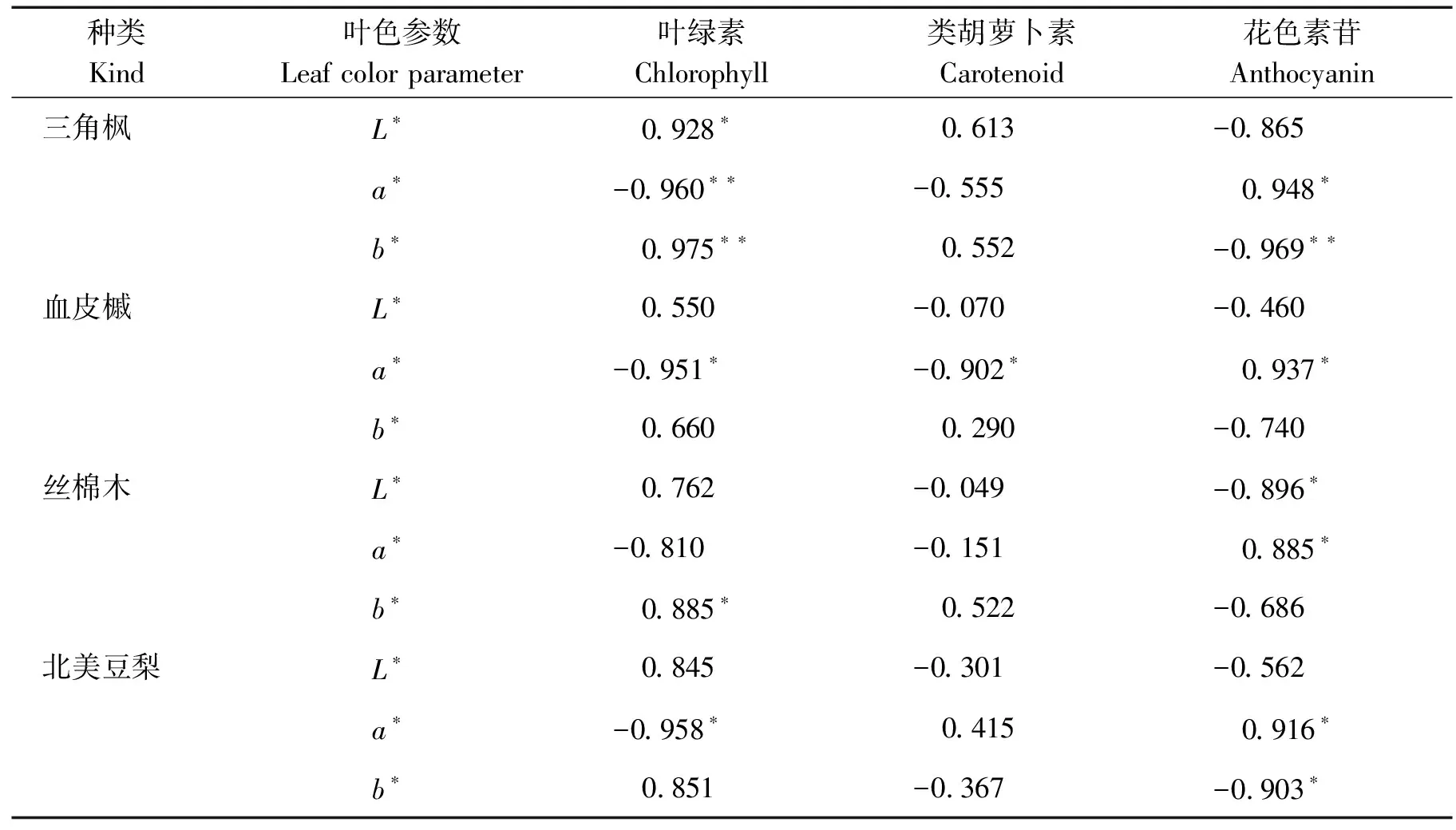
表1 不同树种叶片变色期呈色相关指标的相关系数
注:表中*表示显著相关,**表示极显著相关。
由表1可知,对叶片变色期各树种叶片色素含量与叶色参数进行相关性分析。各树种的a*值均与花色素苷含量呈正相关。除丝棉木外,三角枫、血皮槭以及北美豆梨叶片的a*与叶绿素含量呈负相关,且相关系数达到显著或极显著水平。叶色参数与类胡萝卜素含量均没有显著相关性。
3 结论与讨论
彩叶植物是近年来观赏植物的研究热点之一, 其叶片颜色的变化直接影响园林景观效果。色泽是评价彩叶树种观赏价值的重要指标,叶色参数可以对叶片颜色进行量化,可以准确、方便地描述叶片颜色变化。本试验利用色差仪对叶片参数进行测定,研究结果表明,随着时间变化,4个树种的L*值均有一定程度的降低,但变化幅度较小,无明显差异;各树种叶片的a*值均不断上升,叶片颜色由绿色逐渐变为红色;b*表示黄蓝程度,三角枫在整个变色过程中b*值一直处于下降的趋势,血皮槭呈现先下降后上升,丝棉木和北美豆梨则无明显变化规律,3种植物在变色期b*值变化规律不同。L*值、a*值和b*值共同决定叶片颜色,由于变色期b*值变化规律不同,所以4种植物在秋季变色期叶色变化过程也存在一定的差异。
叶绿素、类胡萝卜素和花青苷等色素的存在使得植物的叶片呈现出不同的色彩。秋季植物叶片变色是由叶片中各种色素含量和比例变化引起的[7]。秋季叶色变红的树种是由于叶绿素降解,花色素苷大量合成的结果。本试验中,随着时间变化,4个树种叶片中所含的叶绿素含量不断下降,花色素苷含量上升,花色素苷与叶绿素含量之比随着时间的变化大幅度增长,使4种植物叶片呈现不断变红的趋势,这与孙明霞等人研究结果存在一致性[8]。
可溶性糖是植物抵御低温的重要保护性物质,可以增强植物对低温的适应能力[9]。陈继卫等认为细胞中花色素的合成必须以足够含糖量为基础[10]。随着温度降低,可溶性糖含量增加,花色素苷含量也随之增加,可溶性糖积累有利于花色素苷的合成[11]。本试验中,各树种的可溶性糖含量均呈单峰曲线的趋势,在含量达到最大值后,花色素苷的含量增长幅度变大,说明在秋季变色期,叶片内可溶性糖的积累导致了花色素苷含量的上升。这与前人的研究一致[12]。
从叶片色素含量与叶色参数间的相关性看,各树种的a*值越大,花色素含量越高,叶绿素含量越低,说明秋季叶片变色期叶片叶绿素含量和花色素苷含量的多少决定着a*值,而类胡萝卜素含量的多少不是影响叶色参数的主要因素。此结论与朱书香等的研究结果相一致[13]。
综上可知,在秋季整个叶色变化过程中4个树种的a*值随时间的延长不断增大,叶片呈现不断变红的趋势,这可能是由于可溶性糖不断积累促进花色素苷合成,花色素苷与叶绿素含量之比不断上升造成的。
猜你喜欢
语文世界(小学版)(2021年5期)2021-06-04 16:30:11
山东林业科技(2019年2期)2019-06-03 10:10:58
小学生必读(低年级版)(2019年12期)2019-04-20 08:39:22
童话世界(2018年23期)2018-08-23 09:13:16
天然产物研究与开发(2018年4期)2018-05-07 06:47:55
山东农业科学(2018年3期)2018-03-29 10:27:08
现代园艺(2017年13期)2018-01-19 02:28:02
小学阅读指南·低年级版(2017年7期)2017-08-04 04:23:38
衡阳师范学院学报(2016年3期)2016-07-10 07:16:27
食品科学(2013年22期)2013-03-11 18:29:17
