
Pathological characterization of occult hepatitis B virus infection in hepatitis C virus-associated or non-alcoholic steatohepatitis-related hepatocellular carcinoma
2020-05-19 06:22HatemElalfyTarekBesheerDinaElhammadyAhmedElMeseryShakerWagihShaltoutMohamedAbdElMaksoudAhmedAminAhmedNasrBekhitMahmoudAbdElAzizMahmoudElBendary
World Journal of Meta-Analysis 2020年2期
Hatem Elalfy, Tarek Besheer, Dina Elhammady, Ahmed El Mesery, Shaker Wagih Shaltout,Mohamed Abd El-Maksoud, Ahmed I Amin, Ahmed Nasr Bekhit, Mahmoud Abd El Aziz, Mahmoud El-Bendary
Hatem Elalfy, Tarek Besheer, Dina Elhammady, Ahmed El Mesery, Mohamed Abd El-Maksoud,Mahmoud Abd El Aziz, Mahmoud El-Bendary, Endemic Medicine Department, Faculty of Medicine, Mansoura University, Mansoura 35516, Egypt
Shaker Wagih Shaltout, Tropical Medicine Department, Faculty of Medicine, Port Said University, Port Said 42511, Egypt
Ahmed I Amin, Internal Medicine Department, Faculty of Medicine, Port Said University, Port Said 42511, Egypt
Ahmed Nasr Bekhit, Tropical Medicine Department, Zagazig General Hospital, Zagazig 44511,Egypt
Abstract
Key words: Occult hepatitis B virus; Hepatitis C virus; Non-alcoholic fatty liver disease;Hepatocellular carcinoma; Clinical outcome; Pathophysiology of occult hepatitis B virus
NATURAL HISTORY OF HEPATITIS B VIRUS, HEPATITIS C VIRUS AND NON-ALCOHOLIC FATTY LIVER DISEASE
Despite an increased understanding of the immunopathogenesis and virology of hepatitis B, the directional course of chronic hepatitis B (CHB) viral infection remains uncertain.Nevertheless, it has been established that the natural history of this disease is primarily dependent on the age at time of exposure to the infecting virus[1].
Chronicity is the hallmark of hepatitis B virus (HBV) infection acquired in infancy,with over 90% of cases developing CHB, while this same percentage of patients undergo resolution of the disease if the infection occurs in adulthood.Therefore, with consideration of the levels of hepatitis B e-antigen (HBeAg) and HBV DNA, in addition to alanine aminotransferase (ALT) values and extent of liver inflammation,the clinical practice guidelines for the management of HBV infection established by the EASL in 2017 have classified the natural history of chronic HBV infection into five phases[2](Figure 1).
Phase I includes patients with HBeAg-positive chronic HBV infection.Previously coined as “immune tolerant”, these patients are characterized as having detectable serum HBeAg associated with high HBV DNA levels, while ALT levels remain in the normal range (ULN, approximately 40 IU/L)[3].Histologically, little or no necroinflammatory changes or fibrosis are detected in these cases.However, the presence of integration affiliated with the increased HBV DNA levels and clonal hepatocyte expansion suggest that the hepatocarcinogenic process may be initiated early in the course of this infection[3,4].Phase 2 comprises HBeAg-positive CHB patients, formerly termed “immune reactive HBeAg positive”.In addition to detectable serum HBeAg and elevated HBV DNA levels, these patients are distinguished by increased ALT levels associated with moderate to severe hepatic necroinflammation with increased evolution to the development of fibrosis.Patients in phase 3, who have antecedently been called “inactive carriers”, are now known to have HBeAg-negative chronic HBV infection.Described by a lack of detectable serum HBeAg with absent or low (< 2000 IU/mL) levels of HBV DNA and normal ALT values, these patients have minimal liver necroinflammation and fibrosis.
Phase 4 of this categorization constitutes cases of HBeAg-negative CHB infection.These patients demonstrate absence of serum HBeAg in association with moderate to high levels of HBV DNA (> 2000 IU/mL).However, ALT levels in these subjects are elevated, the manner being either persistent or fluctuating, and hepatic necroinflammatory activity and fibrosis are evident.The final phase of the EASL classification, phase 5, is the hepatitis B surface antigen (HBsAg)-negative phase,which includes patients with negative serum HBsAg, regardless of the appearance of antibodies to HBsAg (anti-HBs), and the presence of detectable antibodies to hepatitis B core antigen (HBcAg) (anti-HBc).Known as “occult HBV infection”, patients in this phase most commonly exhibit undetectable HBV DNA in serum and ALT levels within the normal range.However, histological examination often demonstrates the presence of hepatic HBV DNA in the form of covalently closed circular DNA(cccDNA).As a consequence, reactivation of HBV infection may occur in patients undergoing immunosuppressive therapy[3].
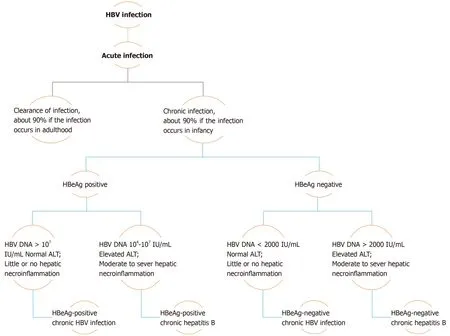
Figure 1 Natural history and assessment of patients with chronic hepatitis B virus infection.
Progression of hepatitis B infection to the stage of cirrhosis and the development of hepatocellular carcinoma (HCC) is of variable incidence and largely affected by immune response of the infected patient to the virus.Untreated CHB patients have a 5-year cumulative incidence of developing cirrhosis of approximately 8%-20%.Those who do progress to cirrhosis have a 5-year cumulative risk of advancing to hepatic decompensation of 20% and an annual risk of HCC of about 2%-5%[5].
As occult HBV infection can only be diagnosed during assessment and evaluation of other liver diseases, such as HCV infection or non-alcoholic fatty liver disease(NAFLD), the natural history of these diseases should be taken into consideration.HCV infection may either manifest mildly, as in acute cases, or present with more serious symptoms in either acute or chronic cases.Chronicity develops in about 80%of patients infected with HCV, and is predicted by several factors such as male sex,age over 25 years at initial infection, mild asymptomatic acute infection, intake of immunosuppressive therapy, and co-infection with human immunodeficiency virus(HIV).The intensity of chronic hepatitis is variable between chronically infected patients but, in general, 5%-15% of patients with chronic HCV infection advance to develop liver cirrhosis over a period of 20 years.Of these cirrhotic patients, 4%-9%will progress to liver failure, with a 2%-5% annual risk of developing HCC[6](Figure 2).
Progression of liver disease identified as simple steatosis to more advanced steatohepatitis with transformation to fibrosis and development of cirrhosis forms the basis for what is collectively defined as NAFLD.While progressive liver disease is a highly unlikely consequence of simple steatosis, the development of non-alcoholic steatohepatitis (NASH) and associated fibrosis have both been shown to be associated with poor prognosis in these patients.This was demonstrated in a study of patients with paired biopsies and showed that 49% of subjects demonstrating features of NASH at baseline proceeded to develop advanced fibrosis in contrast to only 17% of those presenting with simple steatosis[7].However, no difference in mortality was detected among patients with NAFLD in a large population study utilizing blood tests and ultrasonography for assessment of this condition[8].However, compared with the general population, the overall mortality rate was found to be higher in NAFLD patients assessed primarily by radiological evaluation with ultrasound[9].Nevertheless, upon scrutinizing the results of an overlapping cohort with 26.4 years as the mean period of follow-up, only patients diagnosed with late-stage fibrosis had an increased risk of all-cause mortality when compared to the reference population[10].The risk of developing HCC is increased in patients afflicted with cirrhosis in the setting of NAFLD, with about 1%-2% of primary hepatic cancer appearing in patients also suffering from NAFLD[11].
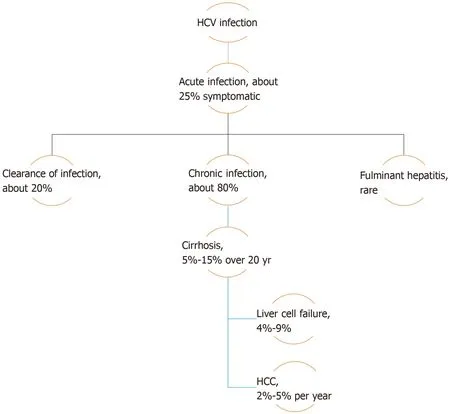
Figure 2 Natural history of hepatitis C virus infection.
INTEGRATION OF HBV GENOME INTO HEPATOCYTES
HBV is a member of theHepadnaviridaefamily characterized by its incorporation of a partially double-stranded relaxed circular DNA of about 3.2-kb that comprises four open reading frames (ORF), each of which encodes for different virion proteins[12].TheSORF encodes the large, middle, and small viral surface envelope glycoproteins collectively known as HBsAg, these being categorized into pre-S1, pre-S2 and S regions based on both structure and function.TheCORF encodes one of two proteins depending on the site of translation initiation, either the HBcAg or viral nucleocapsid,initiated from the core region of the viral genome, or the HBeAg initiated from the pre-core region.Viral polymerase, distinguished by reverse transcriptase activity, is encoded by thePORF, while the small regulatory HBV X protein (HBxAg) is encoded by theXORF[13,14].
Upon entry of an infectious virion into the host hepatocyte, possibly by way of the pre-S protein[15], the process of replication begins with the uncoating and release of the relaxed circular DNA (RC-DNA) followed by its transport to the nucleus of the cell where it is re-organized into the stable form of the viral genome known as the cccDNA.This cccDNA is used as a template for transcription of a group of RNAs utilizing RNA polymerase II enzyme, including pre-genomic RNA (pgRNA), pre-core RNA and sub-genomic HBV RNAs.PgRNA functions as a template for completion of HBV DNA synthesis and serves as the messenger RNA for polymerase and core protein[16], while sub-genomic HBV RNA acts as mRNA for translation of the different sized surface proteins of HBsAg as well as HBxAg protein.At the cytoplasm,produced nucleocapsids are gathered into a glycoprotein envelope in the endoplasmic reticulum forming mature virions to be secreted extracellularly[17].
Development of occult HBV infection, as well as HCC, may possibly be explained by the phenomenon of hepatitis B viral integration into the host genome[18], a process that comprises the redistribution of HBV DNA sequences into host chromosomal DNA.This integration mechanism is often defective[19], and is associated with decreased virion production accompanied by undetectable HBV DNA and HBsAg in the serum of infected patients[7].Integration is commonly seen in chronically infected HBV patients, particularly those with HBsAg-positive HCC[20], although it has also occasionally been seen in HBsAg-negative HCC cases.However, HBV viral integration has not been detected in patients co-infected with HCV[21-23].
Defects that have been associated with HBV DNA integration include loss of HBV core gene with consequent loss of core protein, leading to viral assembly at substandard levels with excessive aggregation of viral DNA in hepatocytes.This may possibly offer clarification for findings of detectable HBV DNA in the liver but not in serum of infected patients[18].Excessive translation of the large protein of HBsAg causes impaired release of other forms of the surface protein, leading to their aggregation in the form of granules in the hepatocyte cytoplasm[24].In addition,HBsAg expression is also affected by HBV DNA disruption and rearrangement when the downstream region of theSORF is replaced with the pre-S1 promoter, leading to decreased S gene transcription associated with impaired secretion of S protein[18].
REACTIVATION OF OCCULT HBV INFECTION
Patients with occult HBV infection (OBI) show HBV DNA existing in two forms,either as episomal free cccDNA or integrated into DNA of the host hepatocyte.The state of OBI may be induced by a number of situations including the subsequent resolution of an acute HBV infection, the “a” determinant arising from mutation in the HBsAg gene, co-infection with HCV or HIV, or epigenetic changes in the cell.OBI is defined by the absence of detectable HBsAg in serum regardless of the status of the antibodies against HBcAg (anti-HBc)[25].
Therefore, cases of immune suppression are associated with cessation of the decreased viral replication and suppressed gene expression that commonly accompany OBI, resulting in reactivation of viral replication consequent to the deterioration of control by the immune system.This provides crucial, albeit indirect,evidence of the important function of immunological control in induction of the state of occult HBV infection.However, recent studies have reported that reactivation of OBI also occurs with histone deacetylase inhibitor use, thus providing confirmation of the role of epigenetic mechanisms in maintaining HBV activity in check with modification of the structure and dynamics of viral cccDNA mini-microsome, these also being projected as probable causes of reactivation of HBV[26].
Conditions associated with a high risk of reactivation of OBI include hematological malignancies, in addition to transplantation with hematopoietic stem cells and immunosuppressive therapies consisting of anti-CD20 monoclonal antibody(Rituximab), drugs included in the CHOP regimen as well as fludarabine.However,only a small number of patients undergoing these therapeutic interventions experience severe clinical manifestations following alteration of the HBV serological profile[27].
On the other hand, reactivation of OBI appears to occur infrequently in sufferers of rheumatologic diseases subjected to high doses of corticosteroids or biological agents for prolonged periods of time.In addition, liver cancer patients going through transarterial chemoembolization (TACE) and those with inflammatory bowel diseases treated with biologics rarely experience OBI reactivation, although patients undergoing chemotherapy for solid tumors have increasing shown HBV reactivation.As a result of these inconsistencies, consensus regarding this topic remains inconclusive, with the entire scope of risk remaining obscure[27].However, the possibility of reactivation of HBV in patients with OBI undergoing direct acting antiviral therapy for the treatment of HCV has been raised in recent reports, although there appears to be negligible risk in these instances and no clinical or virological consequences[27].
MOLECULAR MECHANISMS UNDERLYING OCCULT HBV INFECTION
HBV genome mutations and deletions
A double looped structure comprising amino acids 124 to 147 is known as the “a”determinant of HBsAg.On account of its abundant cysteine residues, it is known to take part in disulfide bond formation and maintenance of this region[28].Mutations in the “a” determinant propose a serious health burden in that they are undetectable by certain commercial HBsAg assays as well as having the capability to infect both vaccinated and unvaccinated persons[29].The first mutation in the “a” determinant of HBsAg to be reported was the sG145R mutation, which was described in a patient who was both actively and passively immunized[29], after which other mutations were later disclosed both inside and outside the “a” determinant[30,31].In addition, this mutation most likely causes reinfection with HBV in patients following liver transplantation in spite of prophylaxis with hepatitis B immunoglobulin (HBIG),probably due to HBIG induced suppression of the secretion of HBsAg into serum without affecting replication of the HBV DNA[32].
Similar mutations characterized by being undetectable by some commercial HBsAg assays as well as infecting vaccinated subjects, are mutations with the added health complication of inducing lamivudine resistance in patients undergoing this treatment[29].These include the Q563S mutation in HBV polymerase, surface gene mutation sS207R, the V539I mutation in the “C” domain of the polymerase, sS143L substitution in the “a” determinant of HBsAg, and M204I and L180M/M204I mutations of the polymerase gene[33,34].
Furthermore, deficient HBsAg expression associated with low replication is also seen with the G-to-A mutation of the surface gene at position 458.Due to the close proximity of this position to the 5’ splice site of the S gene mRNA, this mutation impedes S gene mRNA splicing through a co-/posttranscriptional mechanism with resultant defective export of S gene mRNA or dysregulated RNA folding[35].Genotype D strains of HBV are the only ones that have evolved to accommodate the accumulation of substitutions thus advocating positive selection, as well as having an acceptor site at nucleotide 202.Splicing at this site as well as at the nucleotide 2986 donor site produced intracellular viral particles lacking the surface protein, this resulting in the accumulation of mutations subsequent to alleviation of coding restrictions[36].
Pre-S region deletion mutations are associated with eradication of HLA-restricted B-cell and T-cell epitopes, and are another form of mutation also accompanied by inadequate expression of the surface proteins of HBV[37,38].Deletions and mutations in the pre-S gene result in modified expression of HBsAg as well as reduced HBeAg and HBV DNA levels in hepatocyte cell lines[39].In addition, pre-S1 and pre-S2 region mutations were found to be associated with low secretion of HBsAg in cell culture systems[40].
CO-INFECTION OF HBV WITH OTHER VIRUSES
Contradictory data exists with regards to the clinical outcome of occult HBV coinfection with HCV.A significant association was noted between occult HBV infection and cirrhosis in patients infected with HCV in a study by Cacciolaet al[41], in spite of the enhanced response to interferon therapy reported in these co-infected patients.A higher likelihood of developing HCC in patients co-infected with HCV and occult HBV has also been reported[42], although no correlation was found between the presence of infection with occult HBV and the severity of liver disease related to HCV[43].Co-infection with HBV and HCV is associated with reduced levels of HBV replication accompanied by diminished expression of hepatic HBsAg[44,45], but patients infected with HBV alone demonstrated significantly higher rates of spontaneous HBsAg clearance[46].
Inhibition of HBV replication and HBV protein production by HCV occurs through a variety of underlying mechanisms, including co-localization of HBV and HCV genomes within the nucleus in which the inhibitory effect of HCV on HBV replication was demonstrated using a double fluorescent in situ hybridization technique[47],although this result could not be replicated in Huh-7 cells[48].In addition, replication of HBV and expression of HBsAg can be inhibited by HCV core protein[49].HBV core and polymerase proteins interact with the package signal (Σ) situated at the 5’ end of HBV pgRNA to start the process of encapsidation[50].Direct interaction of HCV core protein with HBx protein causes obstruction of the core and polymerase binding to this package signal within the hepatocyte, thus preventing encapsidation of HBV with consequent inhibition of HBV gene expression[29].Furthermore, HCV NS2 protein downregulates secretion of HBsAg and HBeAg accompanied by suppression of different cellular and viral promoters[51].
Another viral infection in which occult HBV infection is commonly seen is HIV, in which HBV DNA is also detected intermittently.As a result, recurrent sampling has been proposed for HBV DNA detection in HIV-positive patients, although this recommendation remains controversial[52].
APOLIPOPROTEIN B MRNA-EDITING ENZYME CATALYTIC POLYPEPTIDE
Apolipoprotein B mRNA-Editing Enzyme Catalytic Polypeptide (APOBEC) has been shown to play a role in cytidine deamination and is known to inhibit and edit replication of HIV[53].Similarly, expression of APOBEC3G has been associated with a 50-fold reduction in HBV DNA[54].Inhibition of HBV replication by APOBECs occurs in either a deamination-dependent or a deamination-independent manner.
APOBECs have been shown to edit up to 35% of the HBV genome present in the liver[55], including regions encoding for the surface proteins, the polymerase, and the HBx protein[56].While little data exists demonstrating the non-cytolytic clearance of HBV by APOBEC proteins, this has steadily been improving over the past few years with the appearance of more studies[29].
On the other hand, deamination-independant inhibition can be mediated by enhanced nuclease susceptibility of the pre-genomic RNA associated with HBV core protein[57].In addition, targeting of single-stranded or hybrid HBV DNA is another mechanism of inhibiting early stage HBV DNA[58].Secretion of both HBsAg and HBeAg can also be inhibited by APOBEC3B, although the mechanism remains unknown[59].
HOST IMMUNE RESPONSES AND OCCULT HBV INFECTION
Occult hepatitis B virus infection may be caused by impedance of the immune response to HBV.Modulation of both replication of HBV and synthesis of HBV proteins are the outcome of a number of mechanisms related to the host immune response, including apoptosis, T-cell responses of both cytolytic and non-cytolytic nature, and polymorphisms of vitamin D receptor (VDR)[29].Compared to patients with CHB, those with OBI demonstrated lower expression levels of the apoptotic factor Fas, which plays a role in the apoptosis of infected hepatocytes and removal of old hepatocytes[60].Another potential mechanism for occult HBV is the non-cytolytic immune responses seen to be associated with indistinquishable HBsAg levels[61].Similarly, VDR gene polymorphisms were associated with occult HBV infections and were characterized by variable levels of HBV DNA with HBeAg loss[62,63].
EPIGENETIC CHANGES
Occult HBV infection may be the result of modulated HBV replication and virion production due to control of transcriptional activity by methylation[29].Cytosine methylation of CpG dinucleotides within CpG islands inside the gene promoters has been linked to silencing of genes[64].Furthermore, methylated HBV cccDNA has been detected in liver tissues of infected humans, this being associated with a 90%diminution of HBsAg secretion in hepatocyte cell lines[38].In addition, HBeAgnegative individuals demonstrated methylated cccDNA in a higher ratio to total cccDNA when compared to HBeAg-positive subjects[65].
In addition, enhanced replication of HBV in cell culture is associated with hyperacetylation of cccDNA-bound histones, a finding similarly demonstrated with histone deacetylase inhibitors resulting in acetylated histones bound to cccDNA leading to elevated HBV replication with high levels of HBV transcripts.Conversely,hypoacetylation of cccDNA-bound histones is linked to diminished viral loads associated with recruitment of histone deacetylase[66].Remodelling of the HBV minichromosome with regulation of HBV replication is the culmination of histone phosphorylation and methylation[67].
GENOME INTEGRATION
Integration of HBV DNA sequences into the host genome is an established mechanism by which chronic HBV patients develop HCC[68].During the integration process,interruption and rearrangement of genes into chromosomal DNA may result in either loss of serum HBsAg, reduced virus production, or the appearance of undetectable serum HBV DNA.Therefore, particularly in patients suffering from chronic HBV infection for several years, the fundamental mechanism underlying occult HBV is integration of HBV DNA[29].
Loss of HBV core gene during HBV DNA integration results in HBV core protein loss which is associated with minimal levels of viral assembly with aggregation of unencapsidated HBV DNA in the hepatocyte, this offering an explanation as to why HBV-related HCC is associated with undetectable serum HBV DNA although it may be easily detected in liver tissure[18].
IMMUNE COMPLEXES IN OCCULT HBV INFECTION
Immune complexes containing HBsAg have been demonstrated in the serum of patients with acute HBV infection, those with chronic HBV infection, as well as HBsAg carriers who are asymptomatic[69].HBsAg may be become entrapped with anti-HBc, thus hindering detection of HBsAg by conventional serological assays.In addition, these HBsAg-containing immune complexes have been found in HBsAgnegative occult HBV infected patients with HCC[70].In fact, about 40% of blood donors identified as HBsAg-negative but having positive anti-HBc were found to have detectable HBV DNA, most of these being shown as having HBV-containing immune complexes.These circulating immune complexes demonstrated an absence of nucleic acid changes that may cause changes in the major epitopes of HBsAg, thus providing further confirmation of the role of these immune complexes in OBI[71].
CLINICAL OUTCOME OF OBI (OCCULT HBV INFECTION)AND ITS RELATION TO HCC
Although the clinical characteristics of HCC developing in the setting of OBI has not been extensively studied, several attempts have been made to explore this point.A recent study by El-Maksoudet al[72], performed on 50 Egyptian patients with HCC who had undergone either resection or transplantation, found that over 30% of chronic hepatitis C patients with HCC had OBI.Although no significant association could be ascertained between OBI and clinical tumor characteristics such as size and number, BCLC staging or vascular invasion, findings showed that these patients were younger with histology demonstrating a more advanced grade of 3 or 4.In addition,most of these patients had serological positivity for HBcAg, a possible indicator for prediction of occult infections[72].These findings have been supported by a number of studies[73-75].Therefore, the complexity yet uniqueness of the state of OBI grant it an exceptional status that is worthy of consideration when investigating any liver disease.
CONCLUSION
As OBI does not demonstrate the diagnostic laboratory criteria of conventional HBV infection, it is often missed in patients with HCC developing in the settings of HCV infection or NAFLD.Addressing the natural history of these conditions, and the molecular mechanisms by which hepatitis B virus becomes occult then reactivated in circumstances of immunosuppression emphasizes the unique characteristics of this infection.Studies conducted on occult HBV in patients with HCC on top of HBV have collectively concluded that although no significant association could be determined between OBI and tumor characteristics, the presence of OBI in up to 30% of patients,these being mostly younger patients who also tested positive for HBcAg, does in fact warrant its consideration during the investigative workup of other liver diseases.
 World Journal of Meta-Analysis2020年2期
World Journal of Meta-Analysis2020年2期
- World Journal of Meta-Analysis的其它文章
- European Specialty Examination in Gastroenterology and Hepatology examination — improving education in gastroenterology and hepatology
- Probiotics in inflammatory bowel disease:Does it work?
- Hypoxia and oxidative stress:The role of the anaerobic gut, the hepatic arterial buffer response and other defence mechanisms of the liver
- Helicobacter pylori and gastric cardia cancer:What do we know about their relationship?
- Treatment strategies and preventive methods for drug-resistant Helicobacter pylori infection
- Utility of gastrointestinal ultrasound in functional gastrointestinal disorders:A narrative review