基于细胞亚群调控提升生物合成效率的研究进展
2020-05-13 14:55:04曹燕亭刘延峰李江华刘龙堵国成
生物技术通报 2020年4期
曹燕亭 刘延峰 李江华 刘龙 堵国成
(1. 江南大学 糖化学与生物技术教育部重点实验室 生物工程学院,无锡 214122;2. 江南大学 工业生物技术教育部重点实验室 生物工程学院,无锡 214122)
近年来,合成生物学飞速发展,设计与构建微生物细胞工厂生产高附加值产品成为化学品绿色生物制造的重要手段[1-4]。目前,研究人员已开发出许多合成生物学工具和策略来提高微生物细胞工厂生产的产量、转化率和生产强度,如酶活性和表达水平的优化、竞争途径的阻断、中心代谢改造和基于合成遗传回路的动态调控等[4-6]。随着生物合成的转化率逐步接近理论的最大值,进一步提升生物合成效率变得更加困难,因此亟需开发新的工具和方法[7]。
随着显微镜和流式细胞术等单细胞检测技术的发展和应用,研究发现同基因型的纯培养微生物中单细胞间的生长和代谢表现出异质性,且存在不同的细胞亚群,即非生产、低产或高产细胞亚群同时存在[8-13]。高产细胞亚群由于引入的异源途径会与细胞内源过程竞争底物、核糖体、聚合酶等资源,其中间产物、终产物、基因表达的蛋白等也可能对细胞有毒性而形成生产负担,从而表现出降低的生长适应能力[14-17]。与此相反,低产细胞亚群和非生产亚群消耗营养物质却不能高效合成目标产物,它们可能将更多的营养物质用于细胞生长,因而比高效合成目标产物的高产细胞亚群更加具有生长优势。在长时间或者大规模发酵过程中,更加具有生长优势的低产细胞亚群和非生产亚群会逐渐取代高产细胞亚群,从而降低产物合成的综合效率[18-19]。因此,细胞亚群调控有望成为提升生物合成效率的新途径。
目前已发表综述主要针对细胞亚群产生的机制、细胞亚群对生物合成的潜在影响、以及基于生物传感器的细胞生长与产物合成偶联的细胞亚群调控系统的介绍和展望[7,20-23],缺少关于细胞亚群调控系统适用范围受限和低产细胞亚群逃逸的讨论和解决方法的提出。综上,本文归纳总结了细胞亚群调控系统设计和构建的常用方法,重点讨论了目前细胞亚群调控系统存在的问题及其解决策略。
1 细胞亚群调控系统设计和构建的常用方法
随着合成生物学的发展,研究人员已经利用合成遗传回路开发了细胞亚群调控系统,这些系统结合“奖励-惩罚”规则,利用正向响应产物浓度的生物传感器调控抗生素抗性基因、氨基酸合成途径关键基因和必需基因表达,将产物合成与细胞生长关联起来,赋予高产细胞亚群生长优势,提高纯培养物中高产细胞亚群所占的比例,从而改善了大肠杆菌中游离脂肪酸、酪氨酸和甲羟戊酸的合成[24-25](图1)。
Xiao 等[24]结合流式细胞仪分析了大肠杆菌中游离脂肪酸合成的异质性,发现在同基因型的生产游离脂肪酸的大肠杆菌群体中只有15%的细胞能高效合成游离脂肪酸,且这个亚群的游离脂肪酸总产量超过了整个群体总产量的50%。基于以上分析,研究者们开发了一个利用基于转录调控因子的生物传感器的群体质量控制系统(PopQC)调控细胞亚群,并将游离脂肪酸的产量提高了3 倍。群体质量控制系统包括一个基于转录调控因子FadR 的生物传感器,该生物传感器可以正向响应游离脂肪酸浓度:在低产细胞或者非生产细胞内,FadR 可以结合在启动子PAR上,抑制tetA基因表达;而在高产细胞内,高浓度目标产物的存在可以解除FadR 对启动子PAR的阻遏,下游tetA基因连续表达。群体质量控制系统通过这种方式偶联合成和细胞生长,从而赋予高产细胞亚群生长优势,提升生物合成效率。同样的策略应用于酪氨酸合成,致使酪氨酸产量提高了 2.6 倍[24]。
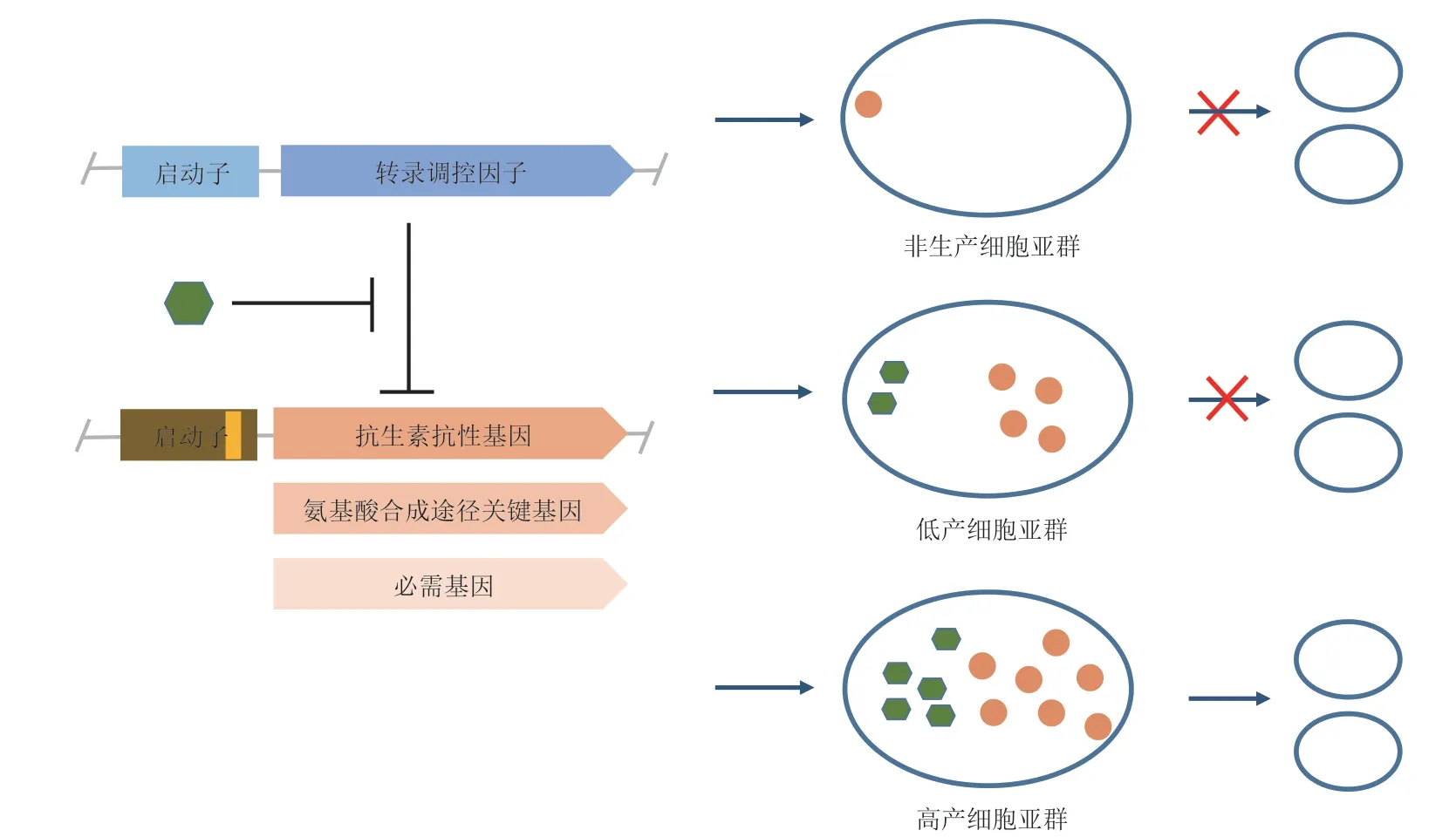
图1 细胞亚群调控系统设计与构建的常用方法
由于抗生素的使用会提高生产成本,增加抗生素污染的机会等,因此一种可避免抗生素使用的细胞亚群调控系统被开发[24]。该系统将生产菌株基因组上的leuABCD基因拷贝敲除后,在质粒上用基于FadR 的生物传感器调控亮氨酸合成途径基因(leuABCD)的表达,导致低产细胞因亮氨酸合成不足而生长受到抑制,高产细胞因为可以合成更多的亮氨酸而更加具有生长优势。应用该细胞亚群调控系统后,游离脂肪酸产量提升了5 倍。虽然该细胞亚群调控系统能提升生物合成效率,且避免抗生素的使用,但是该系统在有效工作前需要进行适当的改造以使菌株成为亮氨酸缺陷型,且培养基中亮氨酸的存在也会导致亚群调控系统失效。
2018 年,Rugbjerga 等[25]利用胞内必需过程,开发了更加具有优势的细胞亚群调控系统。该系统利用生物传感器调控必需基因表达,避免了抗生素的使用,又解决了培养基和宿主受限的问题。在工业发酵后期,改造的细胞工厂合成能力常常降低或者完全丧失,一个重要原因是非生产细胞亚群存在[24,26-27]。因此,该系统利用正向响应甲羟戊酸的生物传感器调控必需基因的表达,从而来惩罚发酵过程中的非生产亚群。因为控制细胞生长可能会给细胞带来一些适应成本[28-29],这可能会降低生物合成效率,所以该系统在选择必需过程时避开了转录、翻译和葡萄糖代谢等必需过程,并替换了调控必需基因的启动子和核糖体结合位点(RBS),以优化细胞亚群调控系统。最终,改造的细胞工厂可以维持高产甲羟戊酸状态直至细胞传代95 次,而没有进行亚群调控的菌株在传代50 次以后甲羟戊酸合成能力逐步丧失。
2 目前已开发的细胞亚群调控系统存在的 问题
目前已开发的细胞亚群调控系统都是基于响应目标产物的生物传感器构建的,因此只能应用于有特异性响应目标产物的生物传感器存在的生物合成途径。随着合成生物学和生物信息学的发展,虽然已经发现了许多能够响应不同种类的小分子代谢物的蛋白质或者RNA 元件(如转录调控因子、核酸适配体等),响应的小分子代谢物涵盖氨基酸、糖磷酸酯、脂类和糖类等,且在此基础之上,已成功开发了许多不同的生物传感器[30-33],但是在实际应用中,仍然存在缺少天然响应或者特异性响应目标代谢产物的生物传感器的问题[34-35],这严重限制了细胞亚群调控系统的可扩展性。除此之外,目前的细胞亚群调控系统都是利用生物传感器直接感知目标产物浓度的变化,利用合成遗传回路富集高产细胞亚群,应用于不同目标产物的生物合成时需要构建不同的生物传感器,增加了细胞亚群调控系统应用的复 杂性。
自发突变和负担是限制很多改造的细胞工厂应用的主要因素[17]。目前已开发的细胞亚群调控系统利用基于正向响应产物的转录调控因子的生物传感器控制细胞生长,这可能会给细胞带来一些生长负担,增加细胞的适应成本[28-29],从而降低生物合成效率。在基于生物传感器调控抗生素抗性基因和必需基因表达的细胞亚群调控系统中,研究者们采用抗生素浓度和添加时间优化、调控必需基因表达的启动子和RBS 替换优化细胞亚群调控系统。优化后的细胞亚群调控系统可以使低产或者非生产细胞亚群的生长受到抑制,而高产或者生产细胞亚群能维持与野生型细胞几乎一致的生长[24-25],一方面减少了因off 状态的泄露导致的低产或者非生产细胞亚群逃逸;另一方面避免引入细胞亚群调控形成负担而降低生物合成效率。但是上述研究未充分考虑到可遗传突变对细胞亚群调控系统的影响。由于同源重组、非同源重组、转座元件、质粒不稳定和胞内外环境压力等因素存在,不论是经过改造的工程菌还是未经改造的野生型菌株都以10-2-10-10/bp/代的突变速率经历着持续进化[17]。而在细胞亚群调控系统中,生物传感器部分转录调控因子因自身突变或转录调控因子的结合位点突变而不能结合到其调控的启动子上,或者调控转录调控因子表达的启动子因突变而不能有效转录(图2)等均可能引起细胞亚群调控系统中调控生长的基因持续表达,造成非生产细胞亚群或者低产细胞亚群逃逸。逃逸的低产细胞亚群在发酵过程中逐步取代因高产失去一定生长优势的高产细胞亚群,从而使细胞亚群调控系统失效。因此,细胞亚群调控系统很有可能因可遗传突变引起低产或者非生产细胞亚群逃逸而失效。
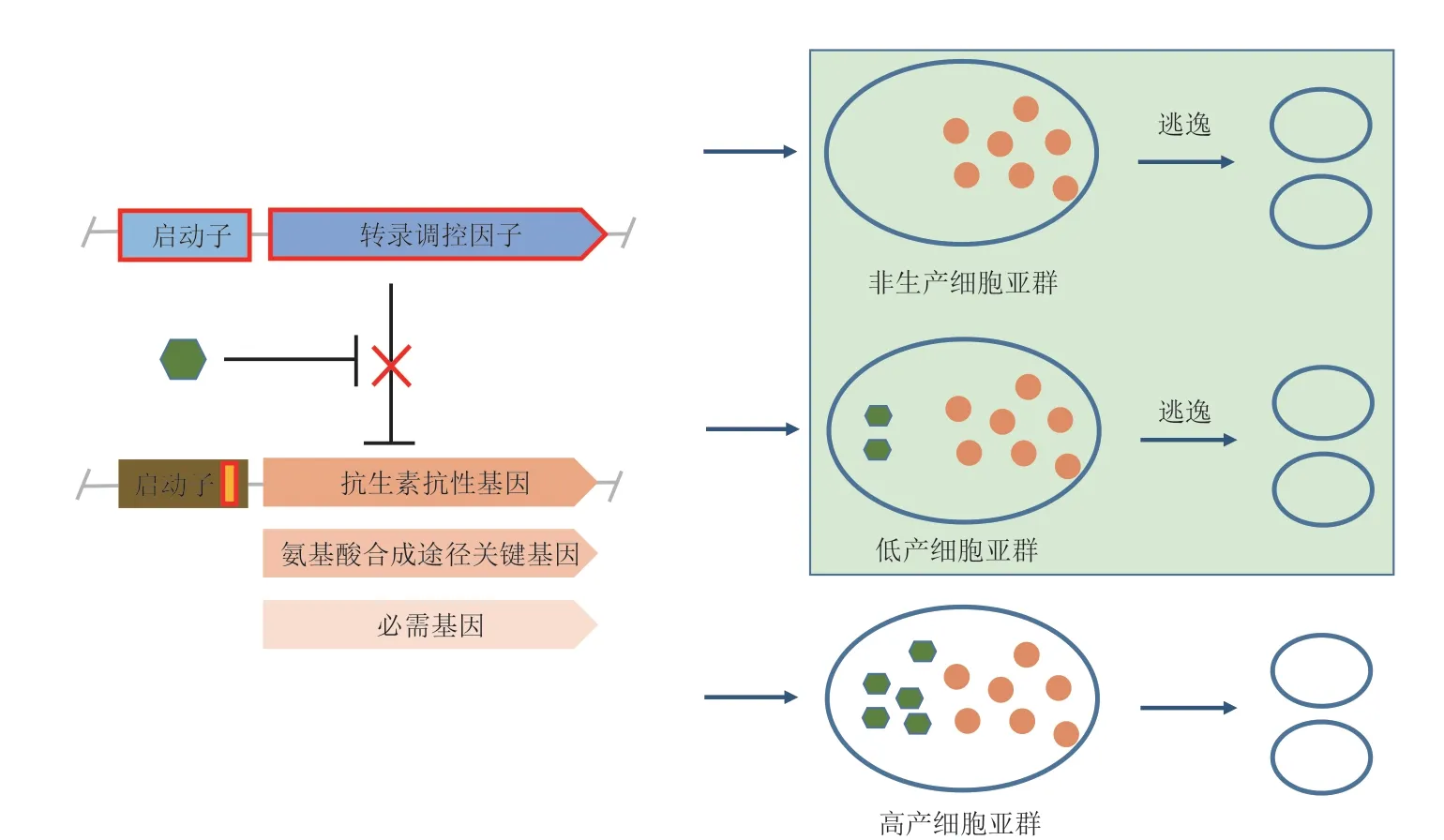
图2 可遗传突变引起细胞亚群调控系统失效
3 目前已开发的细胞亚群调控系统存在问题的解决策略
目前已开发的细胞亚群调控系统依赖于生物传感器,其使用受到可用的生物传感器限制,因此扩展生物传感器的特异性可以扩展细胞亚群调控系统到更多的生物合成途径。结合化学信息学分析、基于转录组的生物信息学分析等,目前已成功鉴定响应多种人们高度感兴趣的目标代谢产物的蛋白质元件,如内酰胺[36]、苯丙氨酸[37]和正丁醇[38]等,以及它们调控的启动子,且在此基础上开发了不同的生物传感器。除了基于蛋白质元件之外的生物传感器,目前也有一些基于RNA 元件的生物传感器,这类传感器因不需要翻译成蛋白质,给细胞造成的负担小而备受关注。经典的SELEX[30]、最近开发的RNA Capture-SELEX[39]、结合下一代测序技术的染色质免疫沉淀法(ChIP-Seq)以及一种原核生物和真核生物均适用的基因组范围内鉴定天然RNA 适配体的方法(PARCEL)[40]等都可用于筛选鉴定新的响应目标代谢产物的RNA 元件,进一步扩展生物传感器的特异性。在实际应用中,天然响应目标产物的RNA 或者蛋白质元件不一定存在。因此,改造已存在的生物传感器的特异性不失为一种有效方法。目前已有综述讨论了如何利用随机突变、结构域组装、计算机辅助设计等技术改变已存在的生物传感器的特异性,扩展其应用范围[34,41],所以这里将不再赘述。除了开发响应小分子代谢物的生物传感器,目前已有感知胞内负担[42-43]或者代谢通量变化[44]等而非特定目标代谢物的生物传感器,基于此设计合理的遗传回路,也许可以富集低负担或者低耗细胞亚群,从而提升产物合成的综合效率;且这类合成遗传回路在应用于不同目标产物的生物合成时,可能不需要构建不同的生物传感器,从而简化细胞亚群调控系统的扩展应用。但基于这类传感器的细胞亚群调控系统的效果还需要实验验证。
在细胞亚群调控系统中,由于自发突变存在,细胞亚群调控系统很有可能因可遗传突变引起低产或者非生产细胞亚群逃逸而失效。目前已有综述总结了通用的降低宿主自发突变率,增加改造的细胞工厂的遗传稳定性的策略,如敲除用于转座的插入序列、敲除或者替换低保真性的DNA 聚合酶编码基因和敲除与同源重组相关的基因,避免使用短的重复序列和高度相似的序列等[17]。因此,本文重点关注如何重构和优化细胞亚群调控系统减少低产细胞亚群逃逸。可能有效减少低产细胞亚群逃逸的策略包括增加响应靶信号的蛋白质或者RNA 元件的拷贝数[45]、串联增加这些响应元件的结合位点数和构建单输入多输出遗传回路[46]等。当增加生物传感器部分能响应靶信号元件的拷贝数或者这些响应元件的结合位点数时,其中一个拷贝或者一个结合位点突变后还有其它的可发挥作用;用生物传感器以单顺反子形式调控两个或者多个不同的控制生长的基因的表达,形成单输入多输出遗传回路,这时只有调控生长的基因前的启动子都发生突变,低产细胞亚群或者非生产细胞亚群才会逃逸。
4 总结与展望
单细胞检测技术的发展帮助揭示了细胞亚群的存在,低产细胞亚群和非生产细胞亚群会降低产物合成的综合效率。细胞亚群调控系统结合“奖励-惩罚”规则,利用正向响应产物浓度的生物传感器将产物合成与细胞生长关联起来,赋予高产细胞亚群生长优势,有效提升了生物合成效率。
基于响应不同目标代谢物的新型生物传感器构建的细胞亚群调控应用范围拓展,基于感知胞内负担或者代谢通量变化等富集低负担或者低耗细胞亚群调控系统设计与构建将会成为细胞亚群调控的新趋势。另外,降低宿主的可遗传突变率、重构和优化亚群调控系统以减少低产细胞亚群或者非生产细胞亚群逃逸,有利于细胞亚群调控系统高效提升生物合成效率。综上,作为细胞代谢调控新工具的细胞亚群调控系统将受到越来越多的关注,设计构建更优的细胞亚群调控系统将成为新的发展趋势。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:56
现代临床医学(2023年1期)2023-03-24 08:30:06
中国医药科学(2022年5期)2022-05-05 23:58:07
今日农业(2021年16期)2021-11-26 06:05:34
天然产物研究与开发(2019年10期)2019-11-05 10:12:44
现代园艺(2017年19期)2018-01-19 02:49:58
山东林业科技(2016年5期)2016-07-05 00:43:08
中国卫生标准管理(2015年3期)2016-01-14 03:41:45
安徽医药(2014年4期)2014-03-20 13:13:40
石油化工应用(2014年6期)2014-03-11 17:39:42