
不同植物当年生枝叶随时间变化生物量分配和水分含量变动策略
2020-04-29 12:56苗杰路兆军陈丽英刘娟孙晓慧李保进
安徽农业科学 2020年7期
苗杰 路兆军 陈丽英 刘娟 孙晓慧 李保进
摘要 通過连续3个月(6—8月)对9种植物当年生枝叶生物量和水分含量及其相对比例、总叶面积和比叶面积等指标进行比较,分析随时间变化植物当年生枝叶生物量分配和水分含量的变化情况。结果表明,不同植物当年生枝条和叶片形态变化差异显著(P<0.05),在枝叶之间生物量分配比和水分含量比表现出枝条越长,植物个体在枝条和叶片上生物量分配和水分含量差距越小的趋势,反之,则表现为枝条越短,相对比例差距越大的趋势;随时间变化,不同植物当年生枝条、叶片在生物量分配、水分含量及相对比例、总叶面积、比叶面积主要分为4种增长类型:6—8月均无显著差异;7月显著增加(P<0.05);8月显著增加(P<0.05);6—8月逐月显著增加(P<0.05)。可以认为,随时间变化不同植物枝条和叶片的生物量投入和水分保持以及枝条与叶片之间的物质分配权衡关系都在不断发生变化,同时表现出一定的规律性。
关键词 当年生枝叶;生物量分配;水分含量;变动策略
中图分类号 Q948文献标识码 A文章编号 0517-6611(2020)07-0129-04
doi:10.3969/j.issn.0517-6611.2020.07.036
The Strategy of Biomass Allocation and Water Content Variation in Currentyear Branch and Leaf of Different Plant Species over Time
MIAO Jie, LU Zhaojun, CHEN Liying et al
(Yantai Institute of Forestry Science, Yantai, Shandong 264013)
Abstract According to contrast biomass allocation, water content and their relative proportion, total leaf area and specific leaf area in currentyear branch and leaf of 9 plant species in three continuous months (June-August), we analyzed the variation of their biomass allocation and water content over time. The results showed that there was a significant difference change in the morphology of each plant branch and leaf(P<0.05). The longer branch length grew, the smaller biomass allocation ratio and water content ratio between branch and leaf they were, and vice versa. We summarized four growth types, which presented biomass allocation, water content and their relative proportion, total leaf area and specific leaf area in currentyear branch and leaf over time, there was no significant difference from June to August; there was a significant growth started from July(P<0.05); there was a significant growth started from August(P<0.05); each month had a significant growth(P<0.05). We concluded that biomass investment and water retention of branch and leaf in different plant species, and the tradeoff between branch and leaf all changed over time, which showed the regularity of themselves.
Key words Currentyear branch and leaf;Biomass allocation;Water content;Variation strategy
基金项目 山东省重点研发计划项目(2018GNC113005);烟台市重点研发计划(2015NC040)。
作者简介 苗杰(1986—),男,山西晋城人,工程师,硕士,从事野生种质资源收集与利用、森林培育、林木遗传育种研究。通信作者,高级工程师,硕士,从事森林培育、林木遗传育种研究。
收稿日期 2019-08-10;修回日期 2019-08-27
在个体发育过程中,植物能够通过控制地上部分枝叶的生长和分布来获取光能和空间等资源[1]。特定的生境中,植物占有更多的资源,在与其他植物共存的情况下具有较强的竞争优势。有研究表明,刺槐(Robinia pseudoacacia)能够通过控制自身小枝叶面积和叶片数量来适应坡向生境的差异,在保证光合效益最大化的情况下,可以避免叶片在不同坡向的空间上互相遮挡[2];五角枫(Acer oliverianum)成树将光合产物主要分配到同化和支持结构上,幼树则主要分配在光合面积的增加上[3];在郁闭度较高的林分中,灌木通过不断分叉和侧向生长来最大程度捕获光能[4]。
植物冠层是由枝条、叶片等构件共同组成。植物冠层中,枝条和叶片在不断重复产生、枯萎和凋落的过程,这些过程决定了树冠的光合作用[5]。植物在长期适应季节性变化的环境过程中形成了一定的生长节律——物候[6]。当遇到有利或者不利的环境时,植物通过生物量投入增加或者减少、部分枝叶产生或脱落的策略维持生存[7]。当年生枝叶生长发育是植物应对环境季节性变化调控自身枝叶构件的实际体现[8]。
目前对于植物调控枝叶生物量分配机制的研究大多基于“源-库-汇”理论和异速生长理论等[7-12],对于当年生枝叶在生物量和水分投入比例差异方面的研究较少。笔者以9种不同植物当年生枝条及其叶片作为研究对象,分析各植物不同时间段在生物量分配和水分含量上的变动情况。
1 材料与方法
1.1 研究区概况
研究材料取自烟台市林业科学研究所种质资源圃,该圃位于烟台市莱山镇曲村(37°21′7.19″N,121°24′53.93″E)。烟台市属温带季风气候,年均降水量651.9 mm,年平均气温11.8 ℃,年均空气相对湿度68%,年平均日照时数2 698.4 h,年均太阳辐射总量5 224.4 MJ/m2,平均全年无霜期为210 d。
1.2 取样
材料取自白檀(Symplocos paniculate)、垂丝卫矛(Euonymus oxyphyllus)、胶州卫矛(E.kiautschovicus)、苦糖果(Lonicera stanishii)、五角枫、一叶荻(Flueggea suffruticosa)、宜昌荚蒾(Viburnum erosum)、玉铃花(Styrax obassia)、紫珠(Callicarpa bodinieri)共9种植物。于2016年6—8月每月中旬取样,每次在各植物树冠上随机选取3个当年生枝条[8],将材料带回实验室进行测量。
1.3 方法
1.3.1 总叶面积。室内对枝条进行编号,将叶片(连带叶柄)与枝条分离。根据枝条叶片总体情况,取适量叶片,平展叶片置于坐标纸上,用铅笔描绘叶片轮廓并进行裁剪,称取1 cm2坐标纸10个,求平均值,然后用0.000 1 g精度分析天平稱取叶片总鲜重和裁剪后的叶片坐标纸重量,采用等比换算法取得总叶面积。
1.3.2 枝叶重量。用0.000 1 g精度分析天平称取枝条鲜重,量取枝条长度。将叶片和枝条置于80 ℃烘箱内连续烘干48 h待恒重,称取烘干后的叶片和枝条生物量,进而获得叶片和枝条的含水量和比叶面积(比叶面积=叶片面积/叶片生物量,SLA)。
1.4 数据分析
1.4.1 不同植物枝叶生长情况对比。
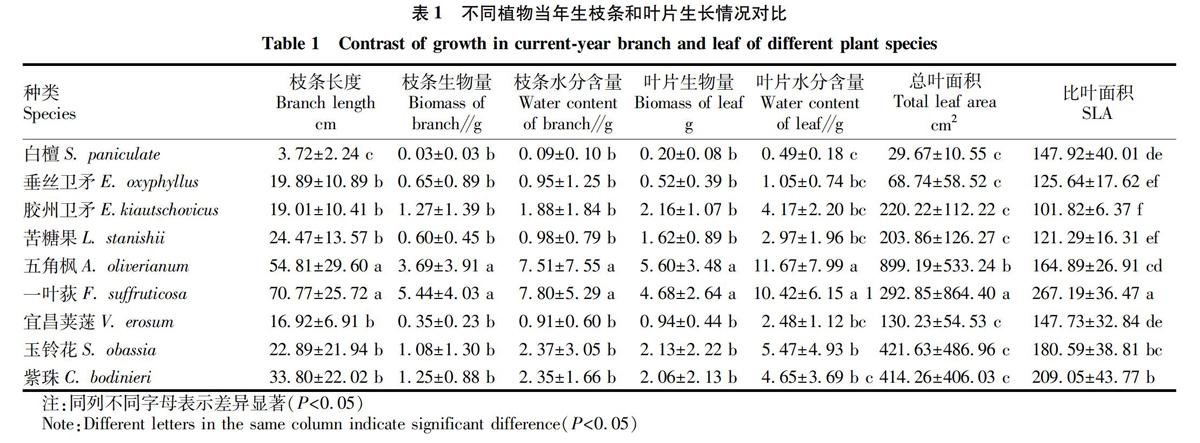
分别对9种植物的枝条长度(branch length)、枝条生物量(biomass of branch)、枝条含水量(water content of branch)、叶片生物量(biomass of leaf)、叶片含水量(water content of leaf)、总叶面积(total leaf area)、比叶面积(specific area of leaf,SLA)进行多重比较(Duncans test)。
1.4.2 不同植物枝叶生物量分配比、水分含量比对比。
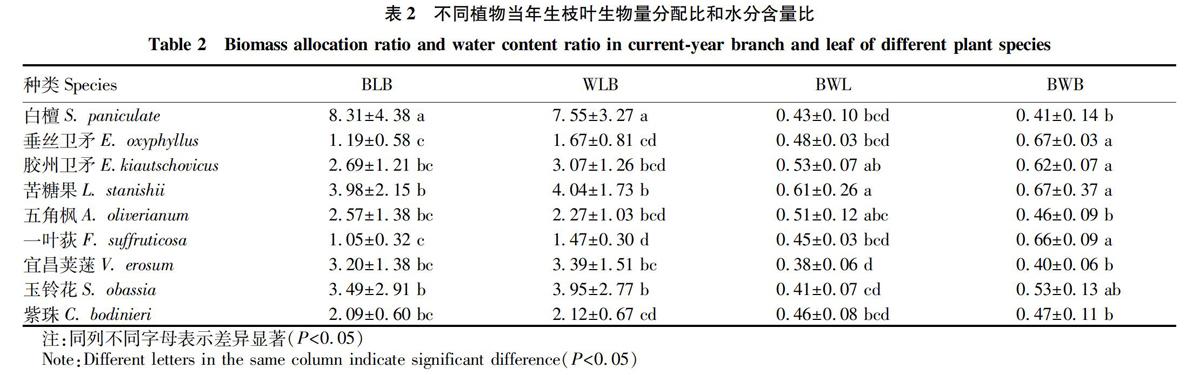
分别对9种植物枝条与叶片生物量分配比、水分含量比,叶片、枝条生物量分配与水分含量比进行多重比较。
叶枝生物量分配比(biomass allocation ratio of leaf and branch,BLB)=叶片生物量/枝条生物量;
叶枝水分含量比(water content ratio of leaf and branch,WLB)=叶片水分含量/枝条水分含量;
叶片生物量分配与水分含量比(biomass allocation and water content ratio of leaf,BWL)=葉片生物量/叶片水分含量;
枝条生物量分配与水分含量比(biomass allocation and water content ratio of branch,BWB)=枝条生物量/枝条水分含量。
1.4.3 不同植物叶片和枝条的生物量和水分含量随时间变化及其相对比例。
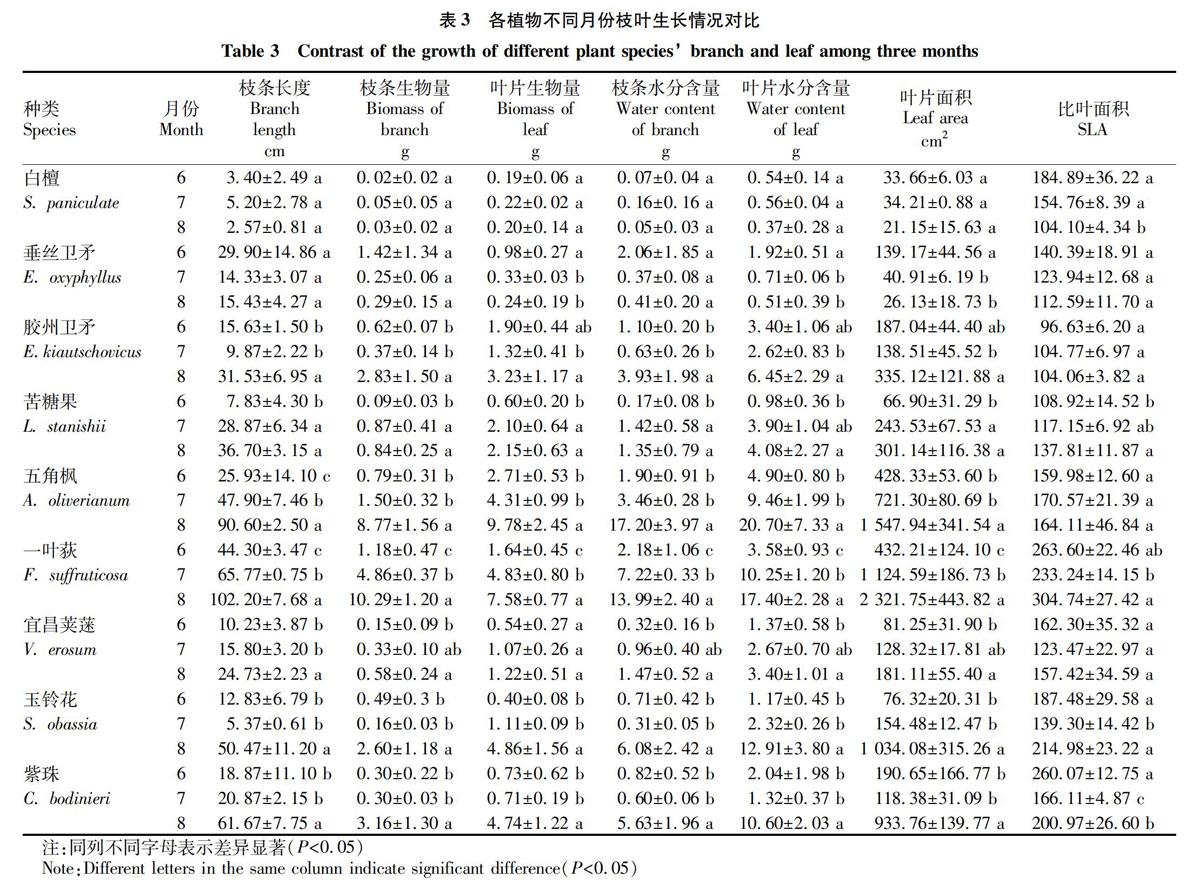
分别对9种植物6—8这3个月中枝条和叶片生物量分配和水分含量及其相对比例进行多重比较。
数据计算、多重比较以及绘图均采用R软件(3.3.1)。
2 结果与分析
2.1 不同植物当年生枝叶生长情况对比
一叶荻和五角枫在枝长、枝条生物量、枝条水分含量、叶片生物量和叶片水分含量上均显著大于其他植物(P<0.05,表1),白檀与其他植物相比在枝长、枝条生物量、枝条水分含量、叶片生物量和叶片水分含量上均小;在总叶面积上,一叶荻>五角枫>其他植物(P<0.05);在比叶面积上,一叶荻>紫珠>其他植物(P<0.05)。
2.2 不同植物枝叶生物量分配比、水分含量比对比
由表2可知,在叶片与枝条生物量分配比和水分含量比方面,均表现为白檀>玉铃花、苦糖果>垂丝卫矛、胶州卫矛、五角枫、一叶荻、紫珠(P<0.05);在叶片生物量分配与水分含量比中,表现为苦糖果>胶州卫矛、五角枫>白檀、垂丝卫矛、一叶荻、玉铃花、紫珠>宜昌荚蒾(P<0.05);在枝条生物量分配与水分含量比中,表现为垂丝卫矛、胶州卫矛、苦糖果、一叶荻>玉铃花>白檀、五角枫、宜昌荚蒾、紫珠(P<0.05)。
2.3 不同植物叶片和枝条的生物量和水分含量随时间变化及其相对比例
由表3可知,从枝条和叶片的生物量和水分对比上来看,大部分植物表现为6月和7月显著小于8月(P<0.05),其中苦糖果和一叶荻表现为逐月增加的趋势(P<0.05)。从叶面积和比叶面积来看,大部分植物叶面积无显著差异。
3 讨论
3.1 不同植物当年生枝叶生物量分配、水分含量和叶片形态差异
研究可以看出,在枝条生物量投入上,一叶荻、五角枫显著大于其他植物(P<0.05),其叶片生物量投入也显著大于其他植物(P<0.05)。枝叶的生长发育和分布对于植物体单位时间内的光合产物总量和光能截取面积具有很大的影响。许月等[4]研究表明植物体在枝条上的生物量投资用来支撑叶片、花和果等器官,有利于植物体对周围空间资源的利用,从而增强自身的光合效率和竞争力,促进自身的生存和发展。这是因为较大的枝条总叶面积必然单叶面积较大或叶片数量较多,容易导致植株个体内部发生互相遮挡,降低叶片对光能的捕获[13]。通过增加枝条生物量的投入,促使枝条增粗和延长生长,不仅能够扩展植株个体叶片的空间分布,而且能够起到对自身及叶片重量的物理支撑作用。可以推断,在相同的生长环境中,一叶荻和五角枫光能截获能力较其他植物强,且具有较强的生长竞争优势。
水分含量是植物体生理过程的重要关键因素之一,如光合作用、蒸腾作用、净初级生产和物质运输等生理生化过程[14],同时也是表征植物代谢速率的指标之一[15]。从枝条和叶片的水分含量来看,一叶荻和五角枫均显著大于其他植物(P<0.05)。 这是因为一叶荻与五角枫在空间占有(枝条长度)和光合面积(总叶面积)上均显著大于其他植物(P<0.05),生物量的生产和运输相对较大,需要植物体内较多的水分来参与光合作用和物质运输等生理生化过程[16-17]。
比叶面积是反映植物体对周围光环境响应策略的一个指标。比叶面积大,植物光合效率高,单位叶面积的投入较少,反之,比叶面积小,植物光合效率低,单位叶面积投入较大[18-20]。从研究中可以看出,不同植物间比叶面积差异显著(P<0.05)。这是因为不同叶片中相同组织的比例不同,不同组织在相同叶片中的比例也不同[21-22],所以产生了不同植物对叶片生物量投入的差异。
3.2 不同植物枝条与叶片生物量分配和水分含量差异规律
从枝条与叶片生物量分配与水分含量比来看,白檀>苦糖果、玉铃花>胶州卫矛、五角枫>垂丝卫矛、一叶荻、紫珠,总体表现为枝条越长,植株个体在枝条和叶片的生物量分配和水分含量差距越小,反之,枝条越短,差距越大。在枝条生物量分配与水分含量比上,垂丝卫矛、胶州卫矛、苦糖果、一叶荻>玉铃花>白檀、五角枫、宜昌荚蒾、紫珠(P<0.05)。这是因为植物在枝条与叶片形态发育过程中,生物量分配比例必须满足机械力学和水分动力学的要求,才能维持枝叶机械支撑的平衡、水分运输和蒸腾的协调。这与Pickup等[23]较大叶片和枝条的植物生物量分配更高的研究结论一致,均符合Corner法则[24]。在叶片生物量分配与水分含量比上,苦糖果最大(P<0.05),宜昌荚蒾最小,其他植物差异较小,说明苦糖果相对于其他植物叶片含水量较少,生物量投入相对较多,可能主要用于叶片形态构建,因为较长的叶寿命需要更加稳固的叶片结构[25-26]。
3.3 当年生枝叶随时间变化生物量分配和水分含量变动策略
该研究表明,不同植物叶片和枝条在6—8月生物量分配、水分含量及相对比例、总叶面积、比叶面积主要分为4种增长类型:6—8月均无显著差异;7月显著增加(P<0.05);8月显著增加(P<0.05);6—8月逐月显著增加(P<0.05)。这是因为物候是植物长期适应生境季节性规律变化所形成的生长节律[27],由于不同植物生境相同,物候表现不同,相同植物生境不同,物候表现也不同[28],因此随时间变化,不同植物在枝条和叶片的生物量投入和水分含量上体现出较大的分配差异。这与郑亚琼等[6]研究结论相同。
綜上所述,植物在枝条和叶片的生物量投入和水分含量持有以及枝条与叶片之间的物质分配权衡关系[2]上都在随时间不断地发生变化,同时表现出一定的规律性。
参考文献
[1] 杜晶,赵成章,宋清华,等.祁连山北坡霸王枝-叶性状关系的个体大小差异[J].植物生态学报,2016,40(3):212-220.
[2] 陈静,赵成章,王继伟.兰州北山半干旱区刺槐叶大小-数量权衡与坡向间的关系[J].生态学杂志,2015,34(12):3300-3305.
[3] 姚婧,李颖,魏丽萍,等.东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化[J].生态学报,2013,33(13):3907-3915.
[4] 许月,杨晓东,谢一鸣,等.浙江天通木本植物小枝的“大小-数量”权衡[J].植物生态学报,2012,36(12):1268-1276.
[5] HIKOSAKA K.Leaf canopy as a dynamic system:ecophysiology and optimality in leaf turnover[J].Annals of botany,2005,95:521-533.
[6] 郑亚琼,冯梅,李志军.胡杨枝芽生长特征及其展叶物候特征[J].生态学报,2015,35(4):1198-1207.
[7] 刘颖慧,贾海坤,高琼.植物同化物分配及其模型研究综述[J].生态学报,2006,26(6):1981-1992.
[8] 李亚男,杨冬梅,孙书存,等.杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响:异速生长分析[J].植物生态学报,2008,32(5):1175-1183.
[9] 刘志国,蔡永立,李恺.亚热带常绿阔叶林植物叶-小枝的异速生长[J].植物生态学报,2008,32(2):363-369.
[10] 史作民,唐敬超,程瑞梅,等.植物叶片氮分配及其影响因子研究进展[J].生态学报,2015,35(18):5909-5919.
[11] 国红,雷相东,刁军.油松当年生小枝异速生长规律研究[J].林业科学研究,2013,26(6):698-703.
[12] 李嵩,郑新军,唐立松,等.基于异速生长理论的准噶尔盆地荒漠灌丛形态研究[J].植物生态学报,2011,35(5):471-479.
[13] 祝介东,孟婷婷,倪健,等.不同气候带间成熟林植物叶性状间异速生长关系随功能型的变异[J].植物生态学报,2011,35(7):687-698.
[14] DE JONG S M,ADDINK E A,DOELMAN J C.Detecting leafwater content in Mediterranean trees using highresolution spectrometry[J].International journal of applied earth observation and geoinformation,2014,27:128-136.
[15] 程栋梁,钟全林,林茂兹,等.植物代谢速率与个体生物量关系研究进展[J].生态学报,2011,31(8):2312-2320.
[16] 龚容,高琼.叶片结构的水力学特性对植物生理功能影响的研究进展[J].植物生态学报,2015,39(3):300-308.
[17] 曹生奎,冯起,司建华,等.植物叶片水分利用效率研究综述[J].生态学报,2009,29(7):3882-3892.
[18] 刘明秀,梁国鲁.植物比叶质量研究进展[J].植物生态学报,2016,40(8):847-860.
[19] NIINEMETS ,PORTSMUTH A,TENA D,et al.Do we underestimate the importance of leaf size in plant economics? Disproportionate scaling of support costs within the spectrum of leaf physiognomy[J].Annals of botany,2007,100:283-303.
[20] HIKOSAKA K,SHIGENO A.The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity[J].Oecologia,2009,160:443-451.
[21] POORTER H,NIINEMETS ,POORTER L,et al.Causes and consequences of variation in leaf mass per area(LMA):A metaanalysis[J].New phytologist,2009,182:565-588.
[22] VILLAR R,RUIZROBLETO J,UBERA J L,et al.Exploring variation in leaf mass per area (LMA)from leaf to cell:An anatomical analysis of 26 woody species[J].American journal of botany,2013,100:1969-1980.
[23] PICKUP M,WESTOBY M,BASDEN A.Dry mass costs of deploying leaf area in relation to leaf size[J].Functional ecology,2005,19:88-97.
[24] CORNER E J H.The durian theory or the origin of the modern tree[J].Annals of botany,1949,13:367-414.
[25] WRIGHT I J,WESTOBY M,REICH P B.Convergence towards higher leaf mass per area in dry and nutrientpoor habitats has different consequences for leaf lifespan[J].Journal of ecology,2002,90:534-543.
[26] WESTOBY M,FALSTER D,MOLES A,et al.Plant ecological strategies:some leading dimensions of variation between species[J].Annual review of ecology and systematics,2002,33:125-159.
[27] 陸佩玲,于强,贺庆棠.植物物候对气候变化的响应[J].生态学报,2006,26(3):923-929.
[28] 常兆丰,韩福贵,仲生年.甘肃民勤荒漠区18种乔木物候与气温变化的关系[J].植物生态学报,2009,33(2):311-319.
