
南海湖非冰封期浮游植物群落生态特征及其与环境因子关系*
2020-04-27 01:52杨文焕李卫平卜楠龙张明钰
环境污染与防治 2020年4期
杨文焕 申 涵 李卫平# 卜楠龙 张明钰
(1.内蒙古科技大学能源与环境学院,内蒙古 包头 014010;2.包头南海湿地管理处,内蒙古 包头 014040)
湖泊具有相对封闭且水流缓慢等特点,当氮、磷等营养物质大量输入湖泊时,就容易造成水体富营养化,城市小型湖泊更易受到外界输入的影响[1]。
浮游植物作为水体中的初级生产者,其种类、密度及多样性对水环境变化反应敏感,常作为评估水体富营养化的重要指标[2-5]。国内外大量学者利用浮游植物来监测和评价水质情况[6-8]。
南海湖(40°30′8″N~40°33′32″N,109°59′2″E~110°2′26″E)位于内蒙古包头市东河区南侧,是黄河改道南移后形成的河迹湖,湖面面积约333 hm2。特殊的地理位置、气候特征和水文地质条件使其成为高纬度寒旱区独特的河迹湖生态系统[9]。近年来,由于受黄河补水量减少、人口增多、城市污染等影响,南海湖水体质量下降,各种浮游植物大量生长,富营养化已成为南海湖主要的环境问题,生态系统受到威胁,特别是非冰封期。于玲红等[10]研究了南海湖的细菌群落及其多样性;杨文焕等[11]研究了南海湖营养盐和叶绿素a的时空分布特征;于玲红等[12]和高静湉等[13]对南海湖的生态系统功能和水质状况进行了分析和评价;姜庆宏等[14-15]对南海湖浮游植物不同时期的群落结构进行了初步研究。然而,南海湖浮游植物群落生态特征及其与环境因子关系的研究目前还鲜有报道。
本研究对南海湖水体环境因子和浮游植物种类、密度等进行了调查,并分析探讨了浮游植物群落生态特征及其与环境因子的关系,以期为南海湖水环境保护及综合治理提供科学依据。
1 材料与方法
1.1 采样点布置及采样时间
参考《地表水和污水监测技术规范》(HJ/T 91—2002),并根据南海湖的地形地貌及其周边环境,共设置12个采样点(见图1)。其中,1号采样点位于进水口处,湖泊出水口已终止出水,湖泊整体靠人工抽取黄河水进行补水,水分散失仅靠蒸发和下渗,5号采样点有一排污口,12号采样点位于芦苇区。采样时间为2017、2018年的非冰封期(5—10月),平行测定两年,每月的数据取两年平均值。
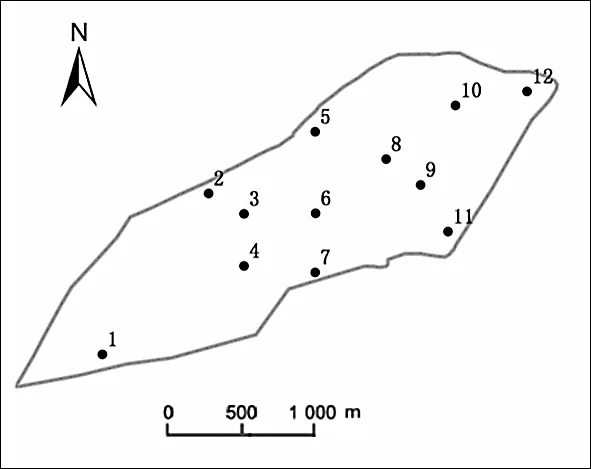
图1 南海湖采样点布置Fig.1 Sampling sites distribution in Nanhai Lake
1.2 样品采集及测定
使用聚乙烯瓶在每个采样点采集1 L水样,加入鲁哥试剂固定,带回实验室静置沉淀48 h,利用虹吸法吸去上清液,将样品浓缩至50 mL,摇匀后吸取0.1 mL在OLYMPUS BX51型显微镜下鉴定浮游植物种类并计数[16-17]。同时在采样现场利用法国PONSEL DDEON双通道多参数便携式水质监测仪测定温度、DO、pH,并用聚乙烯瓶采集1 L水样,带回实验室分别用碱性过硫酸钾消解—紫外分光光度法、过硫酸钾消解—钼酸铵分光光度法、纳氏试剂分光光度法、重铬酸钾氧化滴定法、丙酮萃取分光光度法测定TN、TP、氨氮、COD、叶绿素a。
1.3 数据处理方法
用优势度指数来判断浮游植物群落优势种,计算方法参考文献[18]。
用Margalef丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数对浮游植物群落多样性进行分析,计算方法参考文献[19]。
用生态位宽度和生态位特定重叠指数分析浮游植物群落生态位,计算方法参考文献[20]。
2 结果与分析
2.1 环境因子特征
表1为南海湖非冰封期各环境因子不同月份的变化情况。由表1可知,温度在9.03 ~25.64 ℃之间变化,各月份间温度均存在极显著差异(P<0.01);9月与其他月份pH差异极显著(P<0.01),其他各月份间pH差异不显著,平均值为9.23;DO在5、10月极显著高于其他月份(P<0.01);TN为2.25~3.54 mg/L,平均值为2.88 mg/L,TN随月份增大总体呈现平稳上升的趋势,10月与其他月份差异显著(P<0.05);氨氮为0.66~1.42 mg/L,平均值为1.22 mg/L,5月与其他月份存在极显著差异(P<0.01);TP为0.08~0.47 mg/L,平均值为0.31 mg/L,各月份间TP均存在极显著差异(P<0.01);COD为72.22~134.20 mg/L,平均值为106.12 mg/L,随月份增大呈现逐渐增加的趋势,5、8月与其他月份差异显著(P<0.05);叶绿素a变化幅度较大,随月份增大总体呈现先增加后减少的趋势,变化范围为30.57~61.27 mg/m3,平均值为44.59 mg/m3,7月与其他月份差异显著(P<0.05)。
2.2 浮游植物种类

表1 南海湖非冰封期的环境因子特征
本次调查共发现绿藻门(Chlorophyta)、硅藻门(Bacillariophyta)、蓝藻门(Cyanophyta)、裸藻门(Euglenophyta)、金藻门(Cryphtophyta)、隐藻门(Chrysophyta)和黄藻门(Xanthophyta)浮游植物共计80属157种(含变种)。其中,绿藻门种数最多,共62种,然后依次是硅藻门(40种)、蓝藻门(32种)、裸藻门(10种)、金藻门(6种)、黄藻门(4种)、隐藻门(3种)。从图2可见,位于芦苇区的12号采样点附近浮游植物种数最多,整体呈现东高西低、南高北低的趋势。分析图3可知,浮游植物种数7月最多,10月最少。

图2 浮游植物种数的空间分布Fig.2 Spatial distribution of phytoplankton species

图3 浮游植物种数的时间分布Fig.3 Temporal distribution of phytoplankton species
2.3 浮游植物密度
由图4可知,南海湖非冰封期浮游植物密度为23.35×106~126.00×106个/L,平均值为66.78×106个/L,在时间和空间上均呈现明显的差异,最高值出现在7月的6号采样点,最低值出现在10月的5号采样点。从时间上看,各采样点浮游植物密度呈现出单峰型,5—7月升高,而后下降。从空间上看,各月均呈现出湖心高、四周低的趋势,排污口、芦苇区附近浮游植物密度相对较低。

图4 浮游植物密度时空分布Fig.4 Temporal and spatial distribution of phytoplankton density
由图5可知,5月绿藻门相对密度最大,10月硅藻门相对密度最大,而蓝藻门在7、8月相对密度最大。这是因为绿藻门和硅藻门很多是普生种和冷水种,而蓝藻门大都喜高温,通常在20 ℃以上的水温下才能大量的生长繁殖。

图5 浮游植物各门相对密度Fig.5 The phylas relative density of phytoplankton
2.4 浮游植物群落优势种
根据优势度指数得出南海湖非冰封期浮游植物优势种共13种,其中蓝藻门6种、绿藻门4种、硅藻门2种、裸藻门1种。从表2可以看出,南海湖优势种主要在蓝藻门和绿藻门,检出率较高的优势种有微小平裂藻、水华束丝藻和螺旋弓形藻,其中微小平裂藻连续6个月均为优势种,水华束丝藻和螺旋弓形藻有5个月为优势种,微小平裂藻和螺旋弓形藻指示的水质状态为α-中污带或β-中污带[21],说明南海湖水质总体呈现中污染状态。
2.5 浮游植物群落多样性分析
Margalef丰富度指数评价标准为0~1重污染、>1~3中污染、>3轻污染或无污染[22]。从图6(a)可见,南海湖非冰封期浮游植物Margalef丰富度指数为1.48~3.20,平均值为2.39,总体为中污染。Shannon-Wiener多样性指数评价标准为0~1重污染、>1~3中污染、>3 轻污染或无污染[23]。从图6(b)可见,南海湖非冰封期浮游植物Shannon-Wiener多样性指数为0.82~3.40,平均值为1.81,总体为中污染。Pielou均匀度指数评价标准为0~0.3重污染、>0.3~0.5中污染、>0.5~0.8轻污染[24]。从图6(c)可见,南海湖非冰封期浮游植物Pielou均匀度指数为0.22~0.84,平均值为0.46,总体也为中污染。综上所述,南海湖水质总体处于中污染状态,与优势种的指示结果一致。

表2 浮游植物优势种
注:1)-表示不能明确指示水质状态。

图6 浮游植物群落多样性分析Fig.6 Variety analysis of phytoplankton
2.6 浮游植物群落生态位分析
生态位理论在现代生态学中起着核心作用[25]。生态位宽度不仅能表现物种所占用的资源量,还能反映物种的群落分布规律[26]。生态位特定重叠指数反应的是若干资源类别中每一物种所消耗的资源量,其值正/负表明物种属于发展/衰退状态,其绝对值大小说明该物种发展或衰退的程度,其值若为0,则表明该物种为中性种[27]。
由表3可以看出,在所有优势种中,绿藻门的螺旋弓形藻和蓝藻门的水华束丝藻的生态位宽度较宽,说明这两种浮游植物分布广泛而均匀,数量较多,资源利用充分。

表3 浮游植物群落生态位分析
整体来看,绿藻门拥有较大的发展空间,球衣藻的发展空间最大。绿藻门的四尾栅藻、蓝藻门的为首螺旋藻和阿氏项圈藻、硅藻门和裸藻门的浮游植物呈衰退状态,其中多形裸藻衰退程度最大。
2.7 浮游植物与环境因子的关系
采用CANOCO 4.5软件对浮游植物种群与环境因子进行梯度分析,浮游植物主要考虑优势种以及检出率大于12.5%且至少在一个样点中相对密度大于1%的常见种,环境因子不考虑与浮游植物关系比较明确的叶绿素a。首先对物种和环境因子做去除趋势对应分析(DCA),最大梯度长度小于3,因此进行基于线性模型的冗余分析。
排序轴1和排序轴2的特征值分别为0.245、0.212,其物种与环境因子的相关系数分别为0.913、0.928,说明结果能较好地反映南海湖浮游植物分布及其与环境因子的关系。分析结果表明,TN、TP、pH和温度是影响南海湖非冰封期浮游植物分布的主要环境因子。四尾栅藻、螺旋弓形藻、镰形纤维藻等与TN显著相关(P>0.05),这与李兴等[28]研究得出的乌梁素海绿藻门与TN密切相关的研究结果相似;水华束丝藻、不定微囊藻、阿氏项圈藻与TP、温度具有很大的相关性,李建茹等[29]也认为蓝藻门主要与TP有密切关系;近缘针杆藻、克洛脆杆藻与pH呈显著相关(P>0.05)。
3 讨 论
影响浮游植物群落结构变化的机理是比较复杂的。虽然每种环境因子都可能成为浮游植物生长的潜在限制性因素,但是影响浮游植物的主要环境因素通常是营养盐(特别是氮磷)和温度[30]。一般认为,水体中TN超过0.2 mg/L、TP超过0.02 mg/L时,水华暴发的可能性将大大增加[31]。南海湖非冰封期TN平均值为2.88 mg/L,TP平均值为0.31 mg/L,有较大的潜在水华暴发风险。
优势种可以从其基本生态位中与处于劣势地位的物种进行竞争,甚至在极端情况下会使其灭绝。物种的存在与否主要取决于它们的生存和扩散策略,两个物种之间的竞争可能导致生态位特定重叠指数发生变化。当物种间竞争变得激烈,生态位特定重叠指数便会升高。
在实地采样过程中发现,南海湖非冰封期湖水呈绿色,透明度较低,并且7、8月湖水的颜色更深。南海湖水质总体处于中污染状态说明,浮游植物的大量生长已严重影响了南海湖的水质状况。
4 结 论
(1) 南海湖非冰封期(5—10月)调查发现浮游植物7门80属157种(含变种),以绿藻门、硅藻门、蓝藻门为主,主要优势种为微小平裂藻、水华束丝藻和螺旋弓形藻。浮游植物种数7月最多、10月最少,空间上呈现东高西低、南高北低的趋势。
(2) 浮游植物密度为23.35×106~126.00×106个/L,平均值为66.78×106个/L,时间上呈现单峰型,空间上湖心高、四周低。
(3) 南海湖非冰封期水质总体处于中污染状态。
(4) TN、TP、pH和温度是影响浮游植物分布的主要环境因子,绿藻门与TN关系密切,蓝藻门与TP、温度关系密切,硅藻门与pH关系密切。
猜你喜欢
当代水产(2022年6期)2022-06-29
百科探秘·海底世界(2022年6期)2022-06-24
疯狂英语·新读写(2021年8期)2021-11-05
奇闻怪事(2021年2期)2021-04-20
海洋通报(2020年5期)2021-01-14
流行色(2019年10期)2019-12-06
浙江农林大学学报(2019年4期)2019-07-24
农业与技术(2019年11期)2019-07-12
小品文选刊(2016年21期)2016-11-27
小朋友·聪明学堂(2015年12期)2016-01-07
