
金沙江下游干热河谷岩柿群落特征及种子萌发特性研究
2020-04-07 10:26:10杨永琼潘晶余彦林余志祥李贵能龚丽莉徐登海
陕西林业科技 2020年6期
杨永琼,潘晶,余彦林,余志祥*,李贵能,龚丽莉,徐登海
(1.四川攀枝花苏铁国家级自然保护区管理局;2. 攀枝花市林业技术服务中心,四川 攀枝花 617000;3.湖南工学院,湖南 衡阳 421000)
金沙江干热河谷蜿蜒于四川、西藏、云南等省份,全长约有2 300余千米,是一种位于高温、低湿河谷地带的特殊局部生境,区域内生态条件恶劣,气候炎热少雨,植被稀疏,森林覆盖率不足5%,多以稀树灌草丛为主,乔木不仅数量少,种类也较少,植物恢复困难[1-2],水土流失严重,对长江流域中下游特别是三峡大坝安全具有重要影响,也是长江流域开展以植被恢复为中心的生态环境建设的动因。植物群落的外貌、种类组成和结构既是群落生态环境的反映,也是植物生物学特征和生态学特性的表现,通过研究天然植物群落结构特征,可以为人工群落的生境确定、树种选择与搭配等提供依据和参考[3]。
岩柿(Diospyrosdumetorum)是柿科柿属、泛热带分布常绿乔木[4],天然分布于云南、四川及贵州西南部,海拔700~2 700 m的山地灌丛、混交林、山谷、河边或村边田畔及石灰岩山地[5]。木材可作家具,果可提取柿漆,叶可做药材。是金沙江干热河谷为数不多可构成优势树种的乔木,多分布在海拔1 100~2 200 m之间,自然分布相对狭窄,对维护分布区内的群落稳定起到重要作用。由于生存环境易受到采挖、砍伐、火烧、放牧等人为活动干扰,长势衰弱,数量减少,加之近年来城市化进程加快,外来物种的大量侵入等对岩柿生长造成的威胁,加剧了岩柿野生种群数量的减少[6]。当前岩柿研究报道很少,仅见解剖结构研究[7]。为了解岩柿在金沙江干热河谷的种群数量、分布范围、生态适应性,指导该区植被恢复及引种筛选工作,我们进行了岩柿群落特征、岩柿种子形态与萌发特性等调查研究。
1 研究区概况
研究地点四川攀枝花苏铁国家级自然保护区(东经101°32′15″~101°35′46″,北纬26°36′31″~26°38′24″)(以下简称苏保区),位于川、滇两省交界的横断山脉东南前缘向云贵高原过渡地区,地处金沙江下游河谷地带,气候干燥炎热,雨量集中,干湿分明,年均降水量760 mm, 6-10月降水量占全年降水量的89.2%,最干月(1月)降水量仅1.1 mm,年蒸发量为年降水量的3倍以上。保护区内山高坡陡,多岩石裸露,保护区内土壤发育年轻,具明显的垂直带谱特征:海拔1 500 m以下为红色石灰土,1 500 m以上为棕黄色石灰土,土壤肥力中下。岩柿是苏保区内植物群落建群种之一[2,8]。
2 研究方法
2.1 群落调查
群落调查覆盖整个苏保区。按典型抽样法设置样线(样线密度400条·hm-2),设置5条垂直方向和4条水平方向的样线,每间隔400 m设置一个20 m×20 m样地,共设置样地92个。沿海拔梯度的变化调查样地内植物的种类、数量,盖度等,并对树高2.5 m 以上乔木进行每木检尺,记录胸径、树高,统计样地内岩柿成株(2.5 m以上)、幼树(0.8~2.5 m,低于0.8 m的萌生植株)、幼苗(0.8 m以下,高度木质化实生苗)的株数及频度。在调查中发现,受干热河谷特殊气候条件等影响,岩柿在原生境部分成萌生状灌丛,通常为树干遭破坏后根部萌发形成,岩柿种群年龄组成统计中,萌生植株均不在幼苗统计范畴。
2.2 岩柿种子千粒重与形态特征
从阴干保存的种子中随机选取100粒种子,用解剖刀将其纵剖,根据胚乳饱满度判定种子饱满率。胚乳饱满度低于50%、难于发芽的判定为不饱满,胚乳饱满度高于50%、条件适合可能会发芽的判定为饱满。调查饱满率,重复3次,取其平均值。
随机选取30粒种子,用游标卡尺测量种子长与宽,精度为0.01 mm。取其平均值。
随机选取500粒种子,用电子天平(精度0.001 g)称重测定种子千粒重,重复3次, 取其平均值计算种子千粒重。
2.3 种子萌发及育苗实验
2016年2月22日,选用自然阴干的饱满无病虫害的种子,采用200颗种子进行三个重复试验,用多菌灵300倍液浸泡2 min,清水清洗后用温水浸泡催芽。到2016年2月26日,观察到大部份种子种胚外露时进行育苗。在苏保区珍稀植物繁育苗圃(下称苗圃)内采取粘土、轻基质1∶1进行配比,挖沟15 cm深,形成宽150 cm、长200 cm的苗床组。育苗时种子散播于苗床,覆一层薄土,用薄膜覆盖,苗床顶部用50%的遮荫网遮荫,根据天气情况,人工浇水,保持土壤湿润。采用600~800倍液的噁霉灵和800~1 000倍液的甲霜噁霉灵交互防治根腐病。
从开始出苗起,每周统计出苗株数,直到出苗基本结束,统计幼苗出苗率。
2016年4月20日将实验幼苗进行轻基质营养袋装袋培育。装营养袋前随机抽取30株幼苗测量根长、苗高;6月29日再随机抽取30株幼苗进行苗高测量;2017年6月29日出圃前,随机抽取30株幼苗进行苗高测量,并统计幼苗成苗率及保存数量。
3 结果与分析
3.1 岩柿种群特征
根据样地调查,岩柿从海拔1 100 m到2 200 m均有分布。在设置的92个样地中,岩柿在37个样地中出现,为所有乔木中出现频率最高的植物(40%),比频度最低的小檗美登木高出45%,在苏保区内为广布种(图1)。
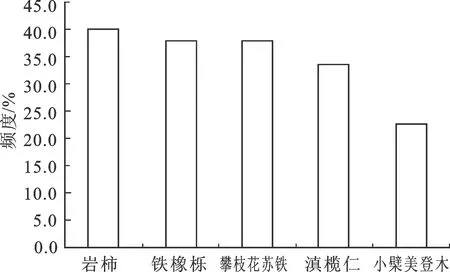
图1 5种乔木植物频度
在海拔1 400 m以下,群落为稀树灌草丛,以草本为主。草本平均高为0.62 m,盖度58%,主要有黄茅(Heteropogoncontortus)、芸香草(Cymbopogondistans);灌木平均高为1.6 m,盖度19%,主要有车桑子(Dodonaeaviscosa)、黄荆(Vitexnegundo);乔木树种少,主要有铁橡栎(Quercuscocciferoides)、岩柿,且多为孤立木,乔木层平均高5.1 m,郁闭度0.15。岩柿平均高5.4 m,处于林冠上层,对光热竞争表现出较强优势与抗逆性。
隧洞所在地貌单元为黄河Ⅳ级基座式阶地,阶地表面黄土覆盖,黄土厚度约7~15 m,以粗粉粒为主,上部风积,下部水成。黄土下伏冲洪积砂卵石层,为黄河古道。形成时代相当于Q3期,砂卵石层与上覆黄土层组成二元结构。下伏二叠系砂岩、页岩、泥岩地层。
在海拔1 400~1 700 m,群落为稀树灌草丛到干性常绿阔叶林过渡状态,乔木树种逐渐成为群落的重要组成部分。草本平均高为0.48 m,盖度32%;灌木平均高为1.8 m,盖度16%;乔木平均高7.4 m,郁闭度0.53;乔木层建群种为铁橡栎、攀枝花苏铁(Cycaspanzhihuaensis)与岩柿。在群落中,岩柿占比乔木树种8.7%,平均高5.8 m,处于林冠中、上层,对光热竞争较有优势,岩柿在此区域在乔木层数量占比虽有所下降,但单位面积数量比稀树灌草丛明显上升。表明这一海拔高度水热条件更适于岩柿分布与生长。
在海拔1 700 m以上,群落为干性常绿阔叶林,群落主要由乔木树种组成。草本平均高为0.42 m,盖度16%;灌木平均高为1.6 m,盖度14%;乔木平均高8.6 m,郁闭度0.76;建群种为铁橡栎、滇榄仁(Terminaliafranchetii)。在群落中,岩柿平均树高6.1 m,低于其他乔木树种,处于林冠中、下层;岩柿在此区域树型更加高大,生长势更好,但占比及数量均有所下降,岩柿在竞争中虽处于劣势,但其抗逆性强(表1)。
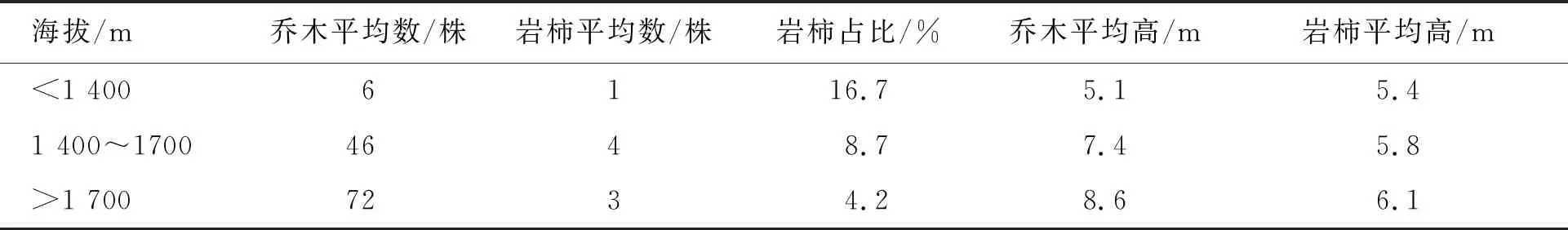
表1 不同海拔样地内岩柿在乔木树种占比及性状
根据样地调查数据统计,在92个样方中共有322株岩柿,其中成年岩柿239株,占比74.2%;幼树69株,占比21.4%;幼苗14株,占比4.3%。受生态学特性、种间竞争及自然和人类活动干扰等影响,调查区域内岩柿自然更新幼苗较少,占统计总株数的4.3%,种群更新能力差。根据调查,岩柿伴生植物种类较为丰富,主要有108种,其中蕨类植物5科5属5种,裸子植物1科1属1种,被子植物40科89属102种。岩柿栖息地分布较多的维管植物有车桑子、铁橡栎、攀枝花苏铁、滇榄仁、黄荆、黄茅、芸香草、水蔗草(Apludamutica)、筒轴茅(Rottboelliaexaltata)、多毛青藤(Illigeracordata)等,群落多样性较高。
3.2 岩柿千粒重与种子形态特征
岩柿种子卵形,黑褐色,长约(9.00±0.08)mm,宽约(5.02±0.12)mm,岩柿千粒重为(107.6±4.9) g,对其种子进行观察测定,平均饱满率(93.3±1.45)%。最高组饱满率可达96%,最低组也达91%,说明岩柿天然种群结实质量较好,种子饱满率高,可为种子繁殖提供较好种源。
3.3 岩柿种子萌发及育苗
岩柿种子经催芽播种5 d后幼苗相继出土。播种后第一周(5 d)平均出苗15株,第二周出苗89株,第三周出苗43株,第四周出苗14株,第五周仅出苗1株(图2)。播种后1~2周是岩柿出苗增长期,第二周时达到高峰期,随后出苗数逐渐减少,约经5周后完成出苗,总计种子出苗率81.15% (表2)。
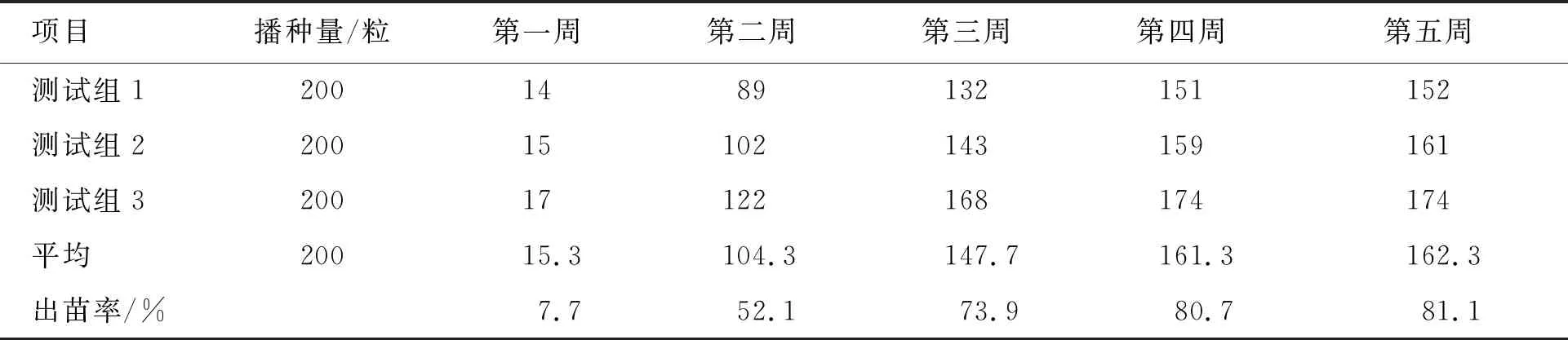
表2 岩柿种子累计出苗情况 单位:株
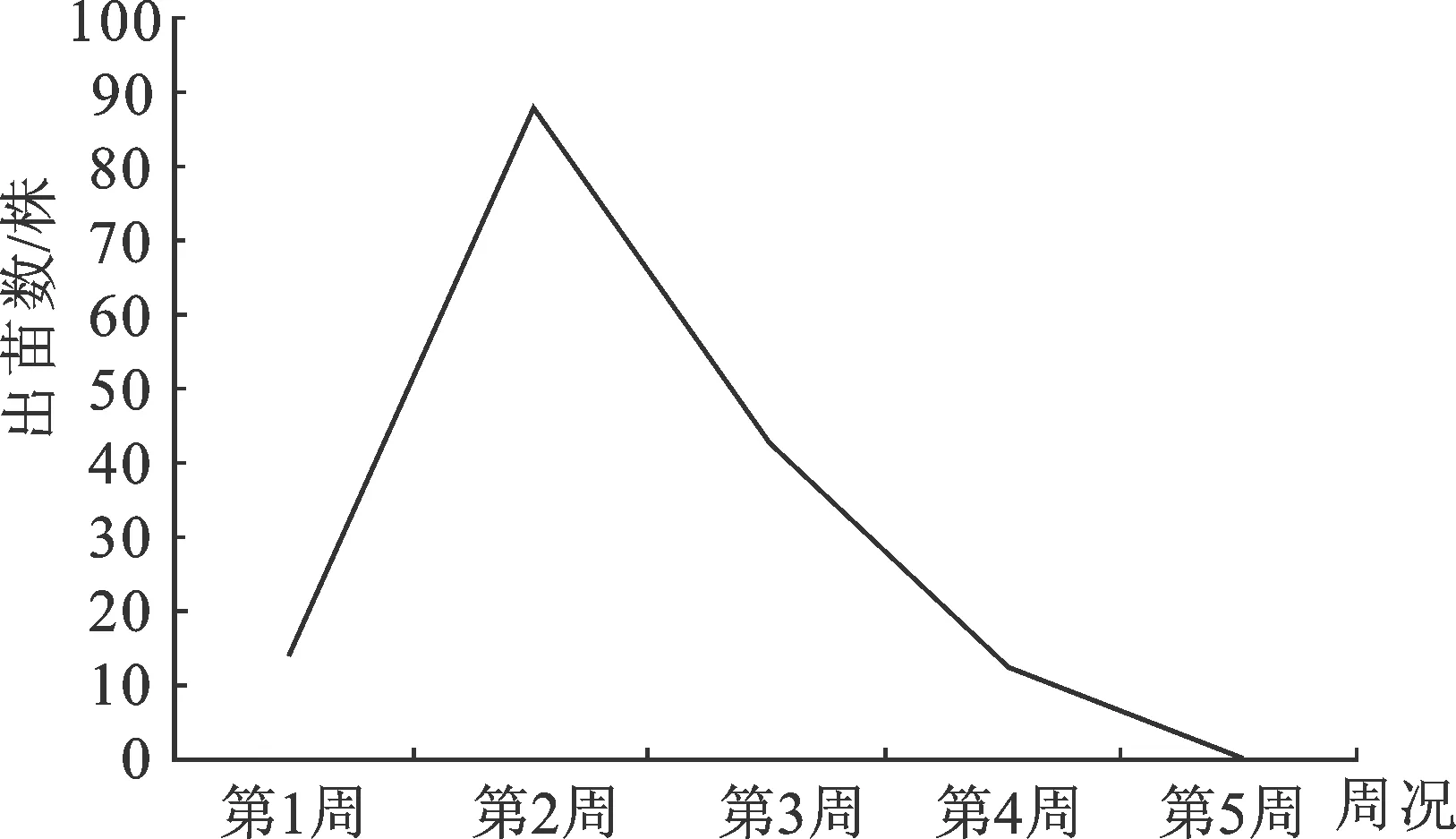
图2 播种育苗各周出苗数统计图
播种5周出苗完成后,在装袋前测量幼苗平均根长为12.0 cm,苗高6.1 cm,根苗比达1.97, 表明岩柿在幼苗生长初期,优先进行根系生长,且根系生长旺盛,而后地上部分生长。较大根苗比为幼苗后期生长提供良好水分与营养吸收保障,有利于岩柿幼苗成活与成长。 播种出苗后第16周(约4个月,当年雨季)幼苗平均高21.6 cm,2年3个月(第二年雨季)幼苗平均高51.3 cm(图3),表明播种后的当年雨季至第二年雨季是岩柿幼苗生长的旺盛时期。第二年雨季后苗高已达出圃要求,至此,600粒种子共培育幼苗353株,育苗成苗率为59%(表3)。
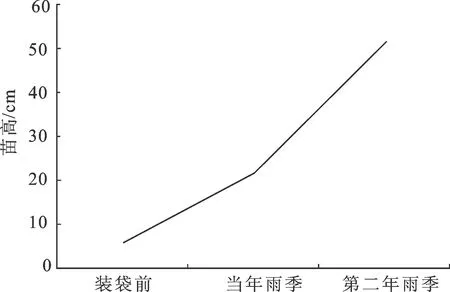
图3 岩柿幼苗各时期苗高

表3 幼苗成苗率统计表
4 结论与讨论
4.1 岩柿抗逆性强、竞争力弱,自然更新力差
岩柿在分布区范围内自然分布相对狭窄,栖息地常受到人类活动的干扰,野生种群数量减少,种群更新受到威胁。在保护区,从河谷1 000 m以上,随着海拔梯度的增加,温度在降低,湿度却在增加。刘洋等(2007)对云南纵向岭谷地区物种多样性的海拔梯度格局分析研究认为,实际蒸散量作为区域水热平衡的表征因子,能较好地解释不同山地植物物种丰富度垂直分布格局及其异同,亚热带山地下部较低的植物物种丰富度受制于高温、少雨的气候特征,随着环境干燥程度的增加植物物种丰富度迅速减小。高海拔地区温度较低,能量构成了植物分布的限制因子,而在低海拔地区温度较高,降雨较少,过多的水分通过蒸发的方式耗散,造成了水热状况的失衡,水分构成了植物分布的限制因子,在温、湿状况理想的海拔范围内容纳了最多的物种数。对低纬热带山地而言,即使在低海拔降雨较少区域,水分仍未构成限制因子,因此植物物种丰富度沿海拔梯度由平缓至递减。而对于中、高纬度亚热带山地而言,低海拔地区水热状况的失衡造成了物种丰富度减小;随着海拔的升高降雨增加、温度降低,越过了水分的界限,能量开始成为限制因子,物种丰富度随着能量的减少而减小,物种丰富度沿海拔梯度呈先增后减的单峰型分布。在金沙江干热河谷区,低海拔地段,焚风效应导致干燥度大,随着海拔升高,干燥度下降,可能是实际蒸散量导致了保护区内随着海拔升高物种丰富度增加的分布格局[5]。岩柿在分布区群落中属乔木层,出现频度最高,但占比及数量均不高,并随着海拔的增高而降低。分析原因应为岩柿抗逆性强,在适生区均有分布,但岩柿为小乔木,竞争力弱,随着海拔升高,湿度增加,其他高大乔木快速生长而使岩柿占比及数量下降。受岩柿伴生维管植物种类丰富影响,岩柿自然更新幼苗极少。从岩柿抗逆性强、出现频度高等特点,岩柿可列为干热河谷造林优选乡土树种。为扩大岩柿种群数量及分布范围,提高岩柿种群质量,需要进行岩柿种群保护和扩繁研究。
4.2 岩柿开花量大、结实能力强,种子饱满率高,能满足人工繁殖需求
岩柿为雌雄同株植物,花期主要集中在4月,盛花期4月中旬,自然条件下花量大,雄花量远大于雌花,能很好满足授粉及繁殖需要。虽然攀枝花岩柿开花时间集中在旱季(4月中旬),但果实生长发育期从4月开始,一直到11月(部份可持续到第二年2月)成熟,持续整个雨季,此期阳光充足、雨水充沛,利于坐果结实。从野外调查结果来看,岩柿的单株结实率非常高,种子千粒重达107.6 g,饱满率及发芽率均较高。
4.3 岩柿种子发芽率高,幼苗易成活,可作为种群繁殖主要方式
年初随采随播的岩柿种子,出芽时间短,仅十余天就可达到出芽高峰,出苗结束时,出苗率可达81.15% ,大量的种子播撒,可保证足量幼苗繁殖。岩柿属于子叶出土型的植物,播种时,覆盖种子的土层不宜过厚,幼苗培育时, 易染根腐病,在育苗过程中,可采用600~800倍液的噁霉灵或800~1 000倍液的甲霜噁霉灵交互防治。
岩柿幼苗出苗初期根苗比高,属于初期先行扎根树种,推测与该树种抗旱性强有关,与岩柿群落调查结果表现较强抗逆性相一致。岩柿幼苗生长缓慢,当年春季随采随播种子育苗到雨季苗高仅21.6 cm,达不到造林需求。为满足干热河谷地区雨季造林需求,岩柿种子育苗应从第一年春季随采随播,培育至第二年雨季。
经过岩柿群落特征及种子繁育研究表明,岩柿种子结实率高,量大;播种育苗出苗率高,易繁殖,能满足人工繁殖需要,可人为扩大岩柿种群范围,提高岩柿种群质量。当前研究仅仅为初步结果,今后应加强岩柿开花结实与雄花、雌花配比以及与气候关系的研究,探索更新机制,为岩柿种群维持提供理论及实践指导。
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
散文诗(2021年22期)2022-01-12 06:14:04
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
河北遥感(2014年4期)2014-07-10 13:54:58
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
