
林龄和竞争对日本落叶松各组分生物量异速关系的影响
2020-03-27 09:49:48陈东升孙晓梅金英博张守攻
生态学报 2020年3期
陈东升,孙晓梅,金英博,张 扬,张守攻,*
1 中国林业科学研究院林业研究所, 北京 100091 2 国家林业局林木培育重点实验室,北京 100091 3 辽宁省清原满族自治县大孤家林场, 清原 113305
生物量在植物器官中的积累和分配规律,是植物生长代谢方式的表征,进而影响植物器官的功能属性,反映植物对环境的适应性[1- 3],也是研究经营措施对植物碳储量和生产力影响的基础[4- 7]。由于生物量在植物器官中分配规律的重要性,许多学者研究了各组分生物量之间的内在关系,认为他们在植物不同器官间的分配方式符合“异速关系理论”[8- 12]。该理论指出生物量分配模式主要受植物本身大小调控,可通过一个幂函数的形式来表征(y=βxα,式中:y和x是因变量和自变量,β是常数,α是异速生长尺度参数),此函数形式被称作异速生长方程。异速生长方程由于其以胸径等易测因子为自变量,且结构简单,已被广泛应用于树木生物量估算中[10- 11, 13-15]。该方程可通过异速生长尺度参数来表征因变量和自变量之间的关系,并决定植物各器官生物量分配随植物大小的变化规律。West等[9]的研究认为地上生物量与胸径是8/3=2.67的比例关系,并与树种、年龄和立地不相关;Zianis和Mencuccini[10]对279个生物量异速方程参数分析发现,地上生物量与胸径的方程尺度参数为2.67时,仅可提供一个粗略的生物量估计;Pilli 等[11]收集了49个不同树种生物量和胸径的数据,得出地上生物量和胸径的异速生物量方程尺度参数约为2.36;也有研究应用异速生长方程理论解释了植物的生物量分配格局,得到林分的叶生物量和树干(或树根)生物量之间存在着4/3的异速关系,而干生物量和根生物量是等比例的[9,16- 17];Robinson[18]通过大量生物量数据获取得到的异速生长模型尺度参数值来预测森林生态系统的根系生物量时,发现实测值和预测值有较大偏差,认为应用固定尺度系数对地下生物量估算时并不普遍适用。所以一些学者认为生物量的积累与分配以及其之间的异速关系并不是严格遵守特定的比例,可能受到树种的耐阴性、林分年龄、竞争、立地条件以及环境因素等影响[11- 12,19-22]。进而开展了许多关于森林生物量分配格局与生物、非生物因子相关关系的研究,认为树种特性、林分特征、气候与土壤因素均会影响生物量分配规律,得到生物量分配规律符合“环境优先理论”的假说[23-29]。但Veronica 等[13]的研究认为生物量分配比例应将“异速生长理论”和“环境优先理论”综合考虑,其既受到植物大小的控制,又受到立地条件、竞争类型和林分年龄等因素的影响,建立不同立地条件、竞争类型和发育阶段的异速生物量方程能显著提高生物量估算的准确性。Satoo和Madgwick[30]以及Helmisaari等[31]也认为各组分生物量分配比例及其异速关系是随着树木生长周期变化的,采用没有考虑林分年龄的特定异速生长方程参数估算各组分生物量时可能出现低估或高估现象。因此,分析林龄、竞争类型等因素对生物量分配规律和其异速性的影响,构建精细的生物量异速方程来准确估算生物量是必要的[26- 27,32]。
本研究以辽宁省清原县大孤家林场采集的日本落叶松(Larixkaempferi)人工林各组分生物量数据为基础,分析年龄和树木竞争类型对各组分生物量分配规律异速性的影响,建立各组分准确的生物量异速生长方程,研究各组分生物量之间的比例关系是否受到年龄和树木竞争类型的影响,从而为准确估算日本落叶松生物量和制定合理的林分经营措施提供理论依据。
1研究区域与研究方法1.1研究区域
研究地在辽宁省抚顺市清原满族自治县大孤家林场(124°47′—1258°12′E,42°22′—44°16′N)。该区地处北方温带气候区,气候类型为典型的大陆性季风气候,春秋季较短,夏季炎热多雨,冬季寒冷漫长,温差较大。年平均气温5.4—7.2℃,年平均降水量400—800mm,无霜期约130d。土壤类型主要为暗棕壤,植被属长白山植物区系。
1.2 研究方法
2011年在7年、17年、30年和40年的日本落叶松林分中分别设置样地9块,总计36块,样地面积0.08 hm2(28.3m×28.3m)。上述林分造林前均采用穴状整地,2年生日本落叶松人工植苗造林,初植密度为3300株/hm2。设置样地后对样地内树木进行每木检尺,样地基本情况见表1。根据样地调查结果,按照等断面积法分别选取优势木、平均木和被压木木各1株伐倒,每个林龄阶段林分伐倒解析木15株,共采伐解析木60株。样木伐倒后使用油锯在树干0、1、1.3、2、3、4m……直到树梢处将树干锯断,称各段鲜重。在各段锯取圆盘,分别测量圆盘干材和树皮鲜重,通过树皮和干材的比值,得出各段干材和树皮的鲜重,并将得到的所有圆盘干材和树皮样品带回实验室烘干处理。将树冠分上、中、下3层,分别称量各层枝叶的总鲜重,从每一层枝条中选取生长良好的3个标准枝,将标准枝摘叶,称其枝重和叶重。根据每层标准枝的枝叶比例和各层枝叶总鲜重,推算枝、叶鲜重。并将每个标准枝的枝和叶鲜样品各取样50g带回实验室。树根采用全根挖掘法,分别测定根桩、大根(≥2cm)、中根(0.5—2cm)和小根(≤0.5cm)的鲜重,然后分别取样200g带回实验室。将树干、树皮、树枝、树叶和根所取样品在85℃恒温下烘干24h直至恒重,根据样品鲜质量与干质量的比例关系分别推算样木各部分干质量。
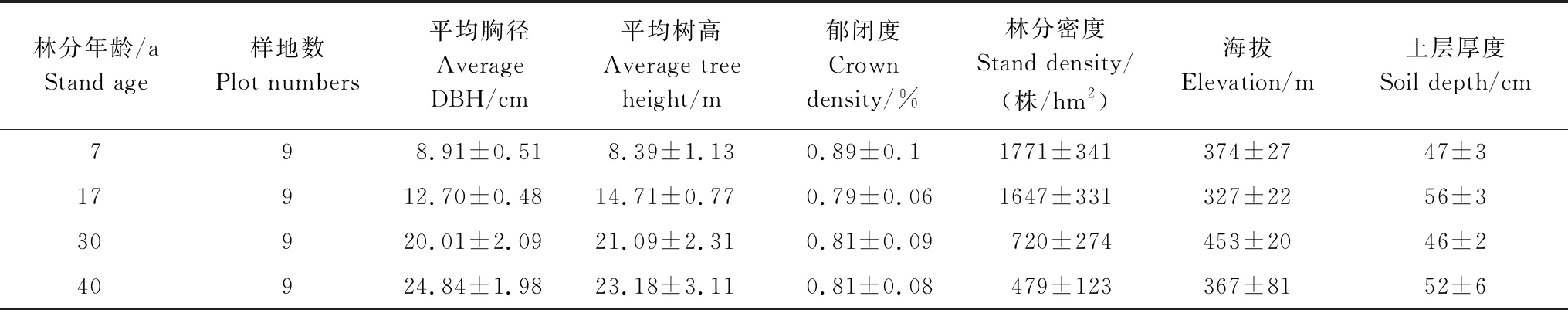
表1 不同年龄阶段日本落叶松样地基本情况
本研究采用Veronica等[13]以胸径作为自变量建立的各组分异速生物量模型(公式1)。
yi=ax1b
(1)
式中,yi是树木各组分(树干、树皮、树枝、树叶、根)的生物量(kg),a、b是方程参数,x1是胸径(cm)。
由于本文旨在分析林龄和树木竞争类型对各组分生物量异速关系的影响,因此,我们在公式(1)的基础上,在方程b参数上引入林龄和树木竞争类型作为哑变量重新构建了模型,进而分析林龄和树木竞争类型对异速生长方差参数b值的影响。哑变量模型是对等级性(定性)数据x,用变量δ(x,i或j)表示成关于定性因子的(0, 1)展开,即关于δ(x,i或j)=(δ(x, 1),δ(x, 2)......,δ(x,m)),其中一个定性变量(m个等级)对应一个向量,一个定性变量就变成可以进行数值运算的数值向量。因此,以林分年龄和树木竞争类型为哑变量,利用定性代码Ki和kj分别表示不同林分年龄和树木等级类型,将定性数据Ki、kj转化为(0, 1):

(2)
式中,i=1,2,3,4(分别表示幼龄林、中龄林、近熟林和成熟林);j=1,2,3(分别表示优势木、平均木和被压木)。
然后根据哑变量模型的计算结果应用SPSS 17.0进行方差分析,分别通过F检验判断林分年龄和树木竞争类型对参数b值的影响。
为了研究不同年龄和树木竞争类型对各组分生物量之间比例关系的影响,进一步应用SPSS 17.0中的方差分析和多重比较的方法来检验不同年龄阶段和树木竞争类型下树冠生物量与树干生物量(Wc/Ws)、树根生物量与树干生物量(Wr/Ws)、树冠与树根生物量(Wc/Wr)比值关系,进而分析其是否受到林龄和树木竞争类型的影响。
2 结果与分析
2.1 日本落叶松各组分生物量异速性
在本研究中,树干、树冠和树根生物量均符合异速生长关系,与树木胸径方程的R2分别为0.89、0.78和0.85,异速生长方程的尺度参数分别为2.53、1.62和2.50(图1)。树冠、树干和树根生物量相互之间也符合异速生长关系。树冠与树干和树根生物量异速生长方程的R2分别为0.70、0.73,异速生长尺度参数分别为0.63,0.55;树根和树干生物量异速生长方程的R2为0.89,异速生长尺度参数为0.95(图2)。研究表明:异速生长方程是适合估算日本落叶松各组分生物量的,当已知胸径或者某一组分的生物量值时,可用其来估计其他组分的生物量。
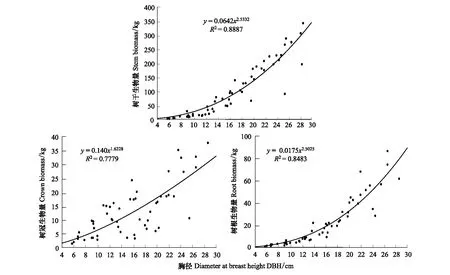
图1 树干、树冠和树根生物量与树木胸径的异速关系Fig.1 The allometric relationship of stem, crown and root biomass with DBH
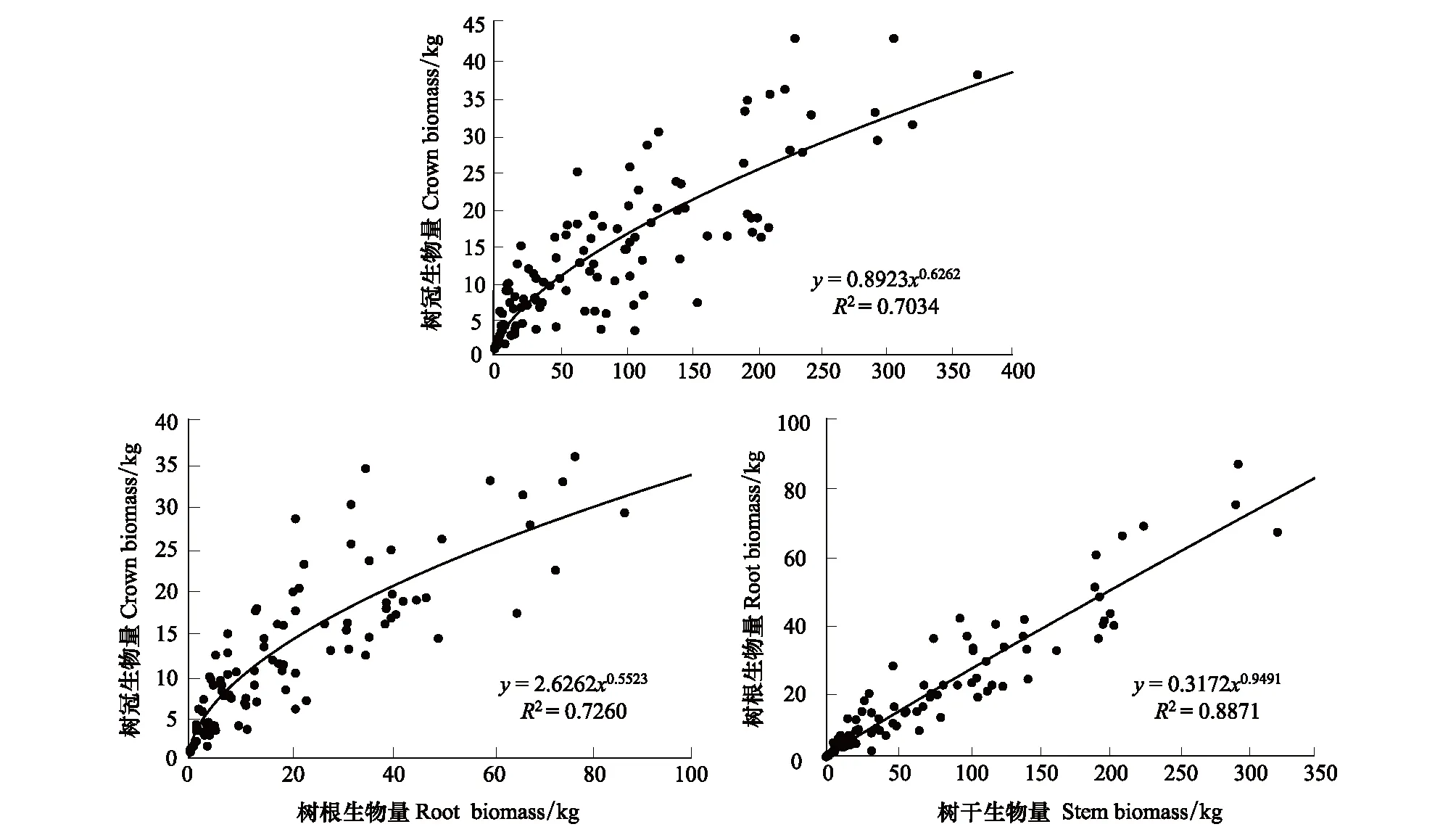
图2 树冠、树干和树根生物量相互之间的异速生长关系Fig.2 Allometric relationships among tree crown, stem and root biomass
2.2 林龄和竞争类型对各组分生物量与树木大小异速关系的影响
通过在公式1参数b中引入林分年龄和树木竞争类型构建的各组分生物量异速生长方程,参数拟合结果和统计量见表2。虽然仅用胸径作为自变量,各组分生物量的异速生长方程参数也有统计学意义(P<0.01),但是枝和叶生物量方程R2较低(0.616和0.533),当模型中增加林分年龄作为哑变量后,树干、树皮、树枝、树叶和根生物量拟合结果均有改善,与公式1相比,F检验显示有显著性差异,其中,尤以枝和叶生物量改善效果最显著,R2分别达到0.757和0.776,提高了22.9%和45.6%。当模型中增加树木竞争类型作为哑变量后,仅树皮、树枝和树叶拟合结果有改善,F检验显示有显著性差异,枝和叶生物量R2分别提高了20.3%和46.5%。这表明在应用异速生长方程预测树冠生物量时,不考虑竞争类型和林分年龄的预测结果是有偏的。
表2 含有林龄和竞争哑变量的日本落叶松林分树木各组分生物量(叶、枝、干、皮、根、地上和总生物量)异速方程
Table 2 Biomass allometric equations for the tree components(needle, branch, stem, bark, root, aboveground and total biomass)with stand age and tree competition as dummy variables
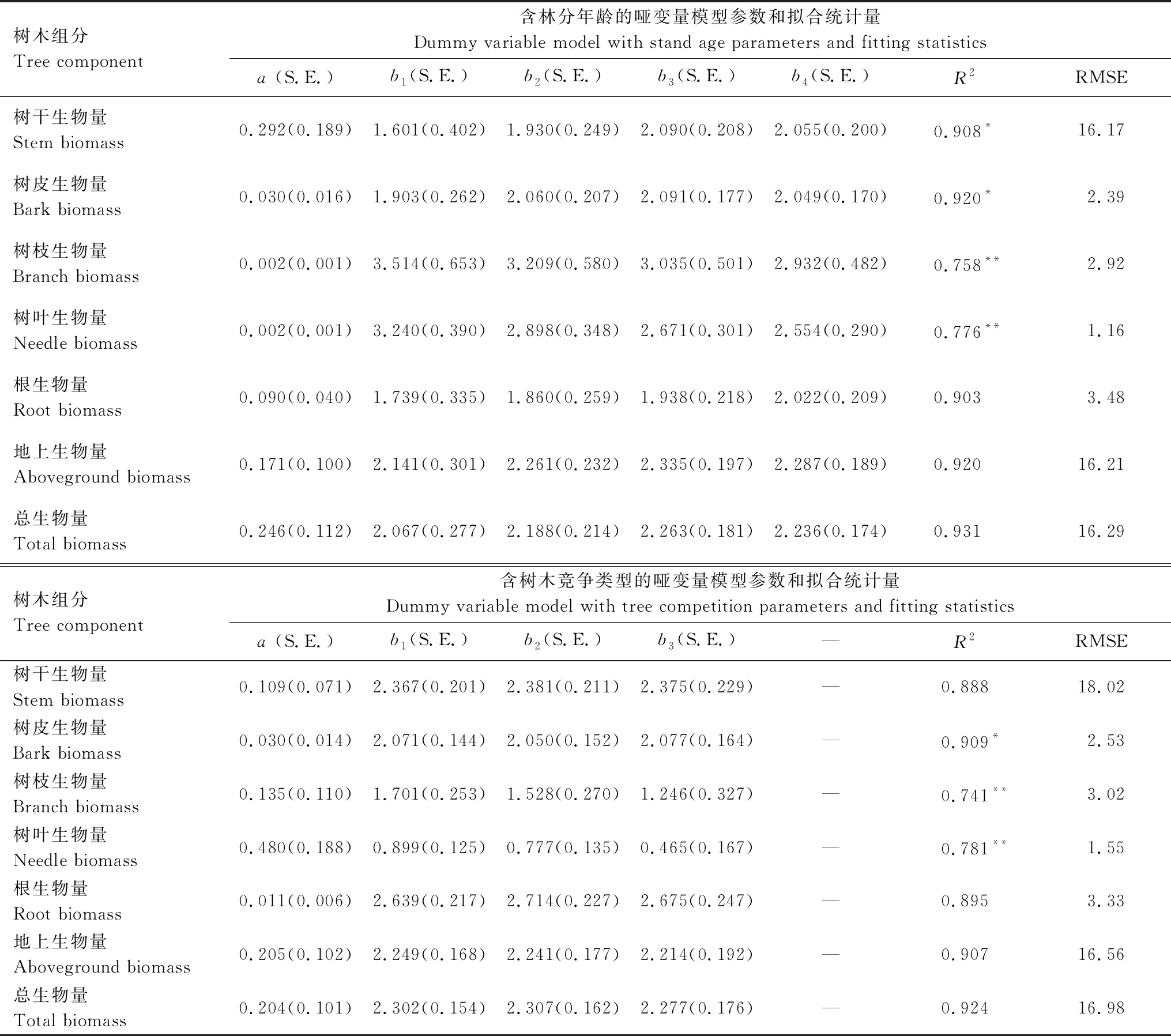
树木组分Tree component含林分年龄的哑变量模型参数和拟合统计量Dummy variable model with stand age parameters and fitting statistics a (S.E.)b1(S.E.)b2(S.E.)b3(S.E.)b4(S.E.)R2RMSE树干生物量Stem biomass0.292(0.189)1.601(0.402)1.930(0.249)2.090(0.208)2.055(0.200)0.908*16.17树皮生物量Bark biomass0.030(0.016)1.903(0.262)2.060(0.207)2.091(0.177)2.049(0.170)0.920*2.39树枝生物量Branch biomass0.002(0.001)3.514(0.653)3.209(0.580)3.035(0.501)2.932(0.482)0.758**2.92树叶生物量Needle biomass0.002(0.001)3.240(0.390)2.898(0.348)2.671(0.301)2.554(0.290)0.776**1.16根生物量Root biomass0.090(0.040)1.739(0.335)1.860(0.259)1.938(0.218)2.022(0.209)0.9033.48地上生物量Aboveground biomass0.171(0.100)2.141(0.301)2.261(0.232)2.335(0.197)2.287(0.189)0.92016.21总生物量Total biomass0.246(0.112)2.067(0.277)2.188(0.214)2.263(0.181)2.236(0.174)0.93116.29树木组分Tree component含树木竞争类型的哑变量模型参数和拟合统计量Dummy variable model with tree competition parameters and fitting statistics a (S.E.)b1(S.E.)b2(S.E.)b3(S.E.)—R2RMSE树干生物量Stem biomass0.109(0.071)2.367(0.201)2.381(0.211)2.375(0.229)—0.88818.02树皮生物量Bark biomass0.030(0.014)2.071(0.144)2.050(0.152)2.077(0.164)—0.909*2.53树枝生物量Branch biomass0.135(0.110)1.701(0.253)1.528(0.270)1.246(0.327)—0.741**3.02树叶生物量Needle biomass0.480(0.188)0.899(0.125)0.777(0.135)0.465(0.167)—0.781**1.55根生物量Root biomass0.011(0.006)2.639(0.217)2.714(0.227)2.675(0.247)—0.8953.33地上生物量Aboveground biomass0.205(0.102)2.249(0.168)2.241(0.177)2.214(0.192)—0.90716.56总生物量Total biomass0.204(0.101)2.302(0.154)2.307(0.162)2.277(0.176)—0.92416.98
a、b1、b2、b3和b4是模型参数,S.E为参数标准误,R2为调整相关系数,RMSE为均方根误差;*表示与公式1比较F检验结果,*P< 0.05; **P< 0.01

图3 7、17、30年和40年生林分树干(带皮)、枝、叶、根、地上部分和总生物量与胸径的的异速生长关系Fig.3 Relationship of stem (with bark), branch, needle, root, aboveground and total biomass with DBH at the 7-, 17-, 30- and 40-year-old stand
年龄对树木各组分生物量异速关系的影响见图3,相同直径的树木,随着林龄的增加,树干生物量增多,而枝和叶生物量减少,但林龄对地上部分和总生物量作用不显著。这也解释了在地上部分和总生物量方程中增加林龄作为哑变量并没有使方程得到明显改善(表2)。图4分析了竞争对各组分生物量异速关系的影响,当树木直径相同时,优势木比劣势木积累更多的叶和枝生物量。但树木其他组分基本不受到树木竞争类型的影响,在表2中的模型拟合结果也同样证实了加入树木竞争类型作为哑变量基本没有改善其他组分生物量异速方程的精度。总的来看,根生物量相对稳定,仅随着胸径而改变,林龄和树木竞争类型影响不大。

图4 不同竞争类型下树干(带皮)、枝、叶、根、地上部分和总生物量与胸径的的异速生长关系Fig.4 Relationship of stem (with bark), branch, needle, root, aboveground and total biomass with DBH at different tree competition status
2.3 林龄和竞争对各组分生物量间比例关系的影响
林龄和树木竞争类型对树冠生物量(Wc)、树干生物量(Ws)和树根生物量(Wr)相互之间比例关系的影响见表3。树冠和树干、树根和树干、树冠和树根生物量比值(Wc/Ws、Wr/Ws和Wc/Wr)在林龄间表现出显著差异。在7年、17年、30年和40年的林分中,树冠和树干生物量比值分别是0.71,0.23,0.09和0.09,随林龄增加先下降然后趋于稳定;树根和树干生物量比值分别为0.35、0.22、0.19和0.23,表现出随林龄增加先减小后增大的趋势;树冠和树根生物量比值分别为0.51、0.99、2.21和2.48,随林龄增大而增大(图5)。但从总体来看,30年生和40年生林分(近成熟阶段)的树冠生物量(Wc)、树干生物量(Ws)和树根生物量(Wr)相互之间比例关系较为稳定,而幼中龄林阶段(7年生和17年生林分)各组分之间的比例关系变化较大,且与近成熟阶段差异较大,因此,当应用固定的异速生长方程参数估算幼中龄阶段的各组分生物量时可能会造成较大的误差。
树木的竞争类型也显著影响着树冠生物量(Wc)、树干生物量(Ws)和树根生物量(Wr)相互之间的比例关系(表3)。优势木的树冠和树干、树冠和树根生物量比值(Wc/Ws和Wc/Wr)与平均木和被压木木差异显著,优势木的树冠和树干生物量(Wc/Ws)比值为0.37,而被压木为0.28;树冠和树根生物量(Wc/Wr)比值为1.09,而被压木为1.69;但树根和树干生物量(Wr/Ws)比值基本稳定,平均为0.25。这表明不同树木间各组分之间的比例差异主要体现在树冠生物量上,而树根生物量相对稳定,因此,当使用固定异速参数估算不同树木间各组分生物量时,会造成树冠生物量估算出现大的误差,而树根生物量估算精度较为稳定。
表3 不同年龄和树木竞争类型对Wc/Ws(树冠/树干)、Wr/Ws(树根/树干)和Wc/Wr(树冠/树根)的影响
Table 3 The difference ofWc/Ws(crown/stem),Wr/Ws(root/stem)andWc/Wr(crown/root)in different tree age classes and competition status
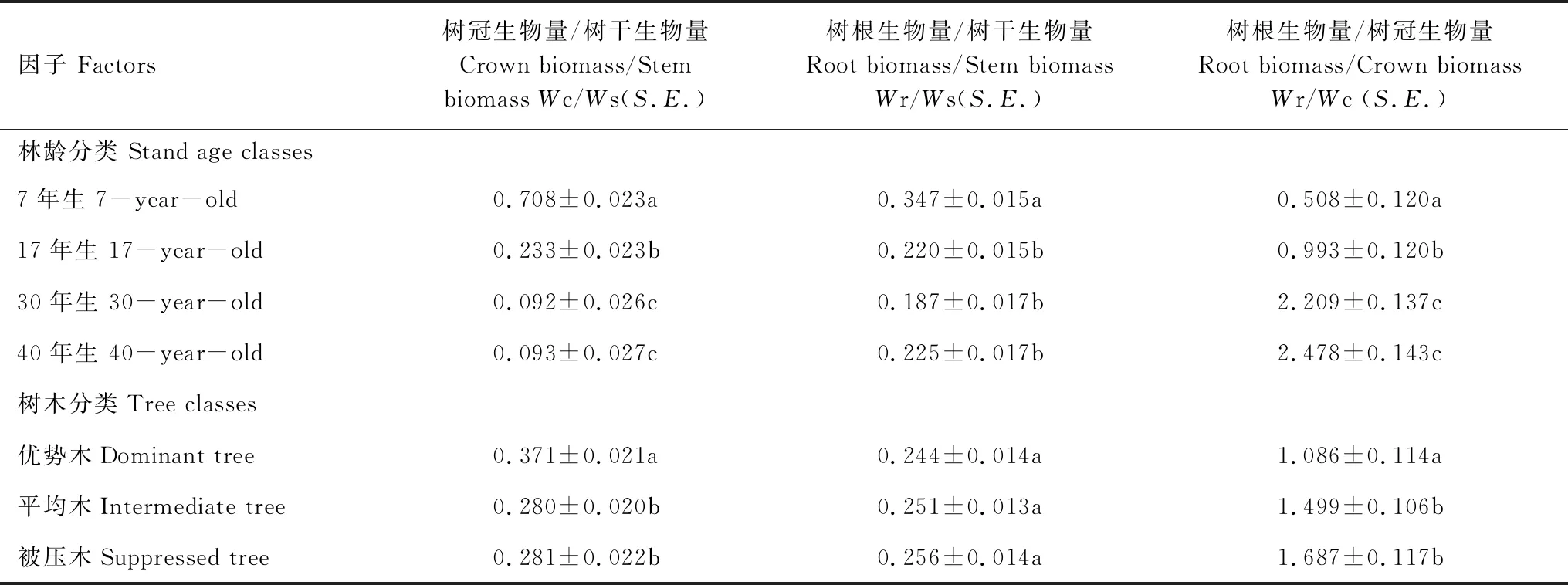
因子 Factors树冠生物量/树干生物量Crown biomass/Stem biomass Wc/Ws(S.E.)树根生物量/树干生物量Root biomass/Stem biomassWr/Ws(S.E.)树根生物量/树冠生物量Root biomass/Crown biomassWr/Wc (S.E.)林龄分类 Stand age classes7年生 7-year-old0.708±0.023a0.347±0.015a0.508±0.120a17年生 17-year-old0.233±0.023b0.220±0.015b0.993±0.120b30年生 30-year-old0.092±0.026c0.187±0.017b2.209±0.137c40年生 40-year-old0.093±0.027c0.225±0.017b2.478±0.143c树木分类Tree classes优势木Dominant tree0.371±0.021a0.244±0.014a1.086±0.114a平均木Intermediate tree0.280±0.020b0.251±0.013a1.499±0.106b被压木Suppressed tree0.281±0.022b0.256±0.014a1.687±0.117b
同列不同字母表示差异显著(P<0.05)
3 讨论
本研究表明日本落叶松各组分生物量分配格局符合“异速关系理论”。这与异速生长方程广泛应用于各树种生物量估算中的结论一致[10- 11,13- 15]。林龄和竞争类型对树木各组分生物量的异速关系影响显著。当林龄作为哑变量加入到各组分生物量异速生长方程时,干、枝和叶生物量异速方程R2均有改善,尤其是枝和叶生物量方程改善最为明显。Peichl和Arain[33]以及King等[34]研究年龄序列下白松(Pinusarmandii)和红松(Pinuskoraiensis)各组分生物量异速生长方程时也发现年龄效应可改善枝和叶生物量估算结果,但对地上、地下和总生物量没有影响,与本研究结果一致。因此建议当预估大区域日本落叶松地上、地下和总生物量时可以仅用胸径作为变量来估算。树木在林分中的优势地位对生物量分配比例也有影响,将树木竞争类型作为哑变量加入到各组分与胸径的生物量方程中对模型有一定改善,但效果不明显。树木竞争类型主要可以通过树高和树冠来反映,许多研究也讨论了树高或者树冠作为第二变量对生物量异速生长方程的影响[35- 37]。Wagner 和Termikaelian[35]将树高作为第二变量加入模型时,仅改善了白松枝和叶生物量预估精度,对根生物量预测没有影响;Cienciala等[37]研究指出树高变量的加入均轻微改善了赤松(Pinussylvestris)各组分的生物量预测效果,我们的研究结果与此一致。Jenkins 等[38]在对美国主要树种异速方程的研究中认为树高并不是一个有效的变量,仅改变小部分组分的预测效果,且树高数据获取困难,会降低预估效率。因此,相对于获取树高因子所消耗的时间和成本来看,考虑树高作为变量加入各组分生物量方程中对模型精度的改善效果并不理想。
林龄对日本落叶松各组分生物量之间的比例关系影响显著,随林龄增加,Wc/Ws先下降然后趋于稳定,Wr/Ws先减小后增大,Wc/Wr持续增大。特别是在幼龄林阶段(7年生林分),它与近成熟阶段林分(30年生和40年生)各组分分配比例差异极显著,表明幼龄林阶段是树木生长的快速时期,各组分生物量的异速关系变化较大,而近成熟阶段生物量分配较为稳定。以往的研究也证实在成熟林中地下生物量与地上生物量成稳定的比例关系[39- 40]。Dickson[41]及Tobin和Nieuwenhuis[42]发现的前10年林分构建完成后,根生物量减少然后稳定在20%左右,本研究中日本落叶松根干生物量比例为0.12—0.41,平均为0.25,幼龄林阶段可达到0.35,到近成熟林也逐渐稳定在20%左右。在林分早期阶段分配较多的资源到根可能是为了帮助植物构建以及强化营养物质的吸收能力[43]。我们的研究也证实了该观点,在幼林中根生物量占据了较大的比例,而成熟林中树干生物量占比较大。在林分生长发育阶段,年龄对生物量分配比例的影响会导致树木和林分水平生理行为的变化。例如,在幼林阶段优势的高生长和树冠扩展对树木的早期竞争及生存是至关重要的,而树干和根生长可能主要集中在后期生长阶段来支撑树木的稳定性。因此在准确估算生物量和碳储量时,年龄对生物量分配比例的影响是需要被关注的,特别是在幼林阶段的研究应该更引起我们的重视。当在中幼林阶段采用根干生物量的标准比值来推算地下生物量是有偏差的,但是在近成熟林分中应用标准比值来估计地下生物量是可行的。
日本落叶松树冠生物量(Wc)、树干生物量(Ws)和树根生物量(Wr)相互之间的比例关系也受到树木竞争类型的影响。优势木的树冠和树干、树冠和树根生物量比值(Wc/Ws和Wc/Wr)与平均木和被压木差异显著。各组分之间的比例差异主要体现在树冠生物量上,而树根生物量相对稳定,当使用固定的比例关系来估算树冠生物量时会产生误差,而树根生物量估算精度较为稳定。Ilomaki等[44]和 Naidu 等[45]在研究白桦(Betulaplatyphylla)和火炬松(Pinustaeda)优势木和被压木对生物量分配比例影响时,被压木分配较多的树干生物量和较少的叶生物量,本研究中日本落叶松被压木生物量分配方式与其一致。从上述研究结果来看并没有因为光是限制被压木生长的主要因子,从而分配较多的叶生物量来获取更多的光照优势,未体现出依据环境变化生物量分配遵守环境优先原则。但Veronica 等[13]分析湿润和干燥土壤条件下不同树木类型山毛榉的生物量分配比例时,发现在干燥的立地条件下树木会分配更多的生物量到根部,同时研究得出强风环境下劣势木也会分配更多的地下生物量来抵抗风。因此,本研究认为当树木在在同一特定环境条件下,林内竞争虽然导致优势木、平均木和被压木在生物量积累上差异较大,但是它们分配生物量到不同器官的方式是相同的[13]。但当环境条件差异较大的情况下,分配生物量到各个器官的比例模式会由于环境的变异出现变化。
4 结论
本研究中证实日本落叶松各器官生物量分配符合异速生长关系,因此,异速生长方程是准确估算生物量分配格局的有效方法。但是,生物量异速方程的尺度函数值受到林分年龄和林木竞争类型的影响。随着林龄增加日本落叶松干生物量比例增大,枝叶生物量比例减小,而根生物量逐渐趋于稳定。将林龄作为哑变量加入到各组分生物量异速方程中,干、枝和叶生物量方程显著改善。在幼龄林阶段(7年生林分)各组分生物量分配比例与近成熟阶段林分(30年生和40年生)差异极显著,幼龄林有最大的根茎比和根冠比。因此在准确估算生物量和碳储量时,年龄对生物量分配比例的影响是需要被关注的,特别是当林分处于幼林阶段时,需单独构建生物量和碳储量预测模型来提高准确性。树木竞争类型对生物量分配的影响小于林龄。当立地条件差异不大的情况下,林内竞争虽然会导致优势木、被压木各器官生物量积累上产生差异,但是它们分配生物量到不同器官的方式是基本相同的。因此在估算各组分生物量时林木等级差异并不会带来太大的偏差,可以不予考虑。
猜你喜欢
小学科学(学生版)(2021年10期)2021-12-28 15:01:27
防护林科技(2020年10期)2020-12-22 11:46:42
少儿画王(7-10)(2020年11期)2020-09-13 02:44:51
东方企业家(2020年5期)2020-05-29 08:13:43
山西文学(2019年8期)2019-11-01 02:14:24
文学港(2019年5期)2019-05-24 14:19:42
小溪流(画刊)(2018年3期)2018-06-29 06:38:56
幼儿画刊(2017年10期)2017-10-18 00:46:02
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
